
Curcumin Reduces Colorectal Cancer Cell Proliferation and Migration and Slows In Vivo Growth of Liver Metastases in Rats

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Preparation
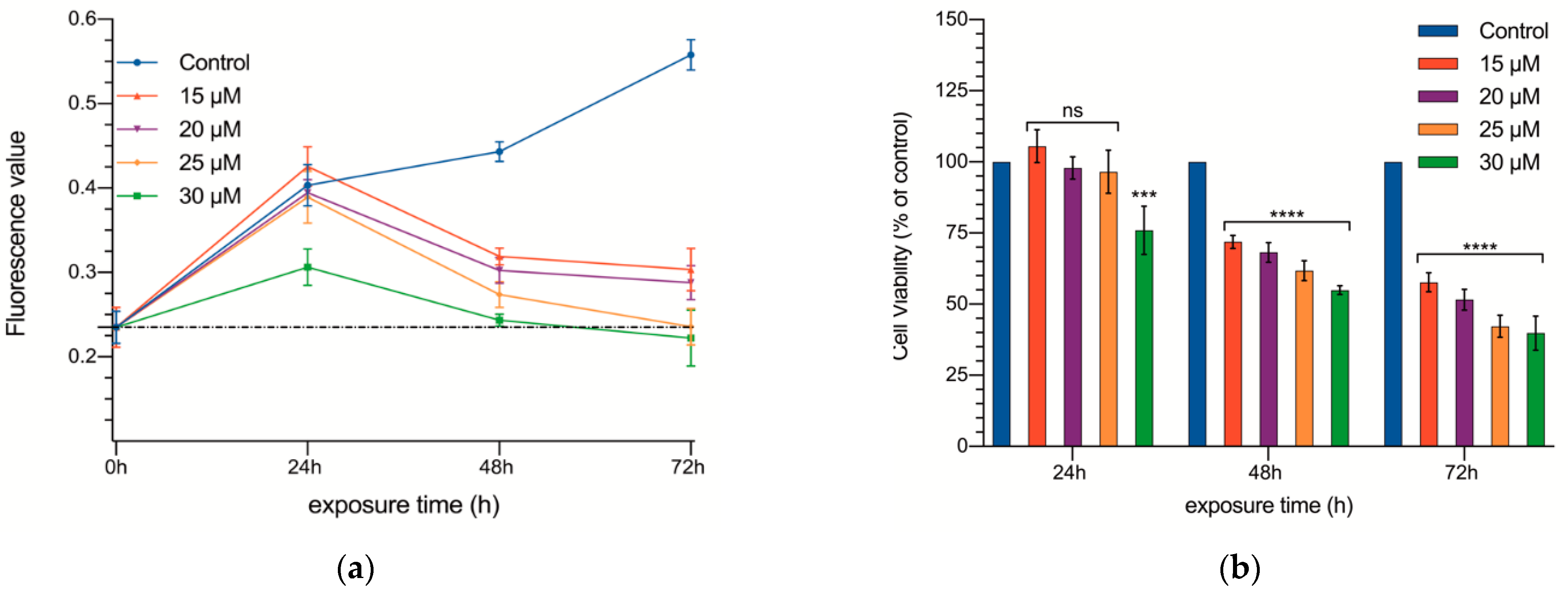
2.2. Proliferation Assays
2.3. Wound Healing Test
2.4. Tumor Induction
2.5. Treatment Administration
2.6. Blood and Tissue Collection and Analysis
2.7. Statistical Analysis
3. Results
3.1. In Vitro Experiments
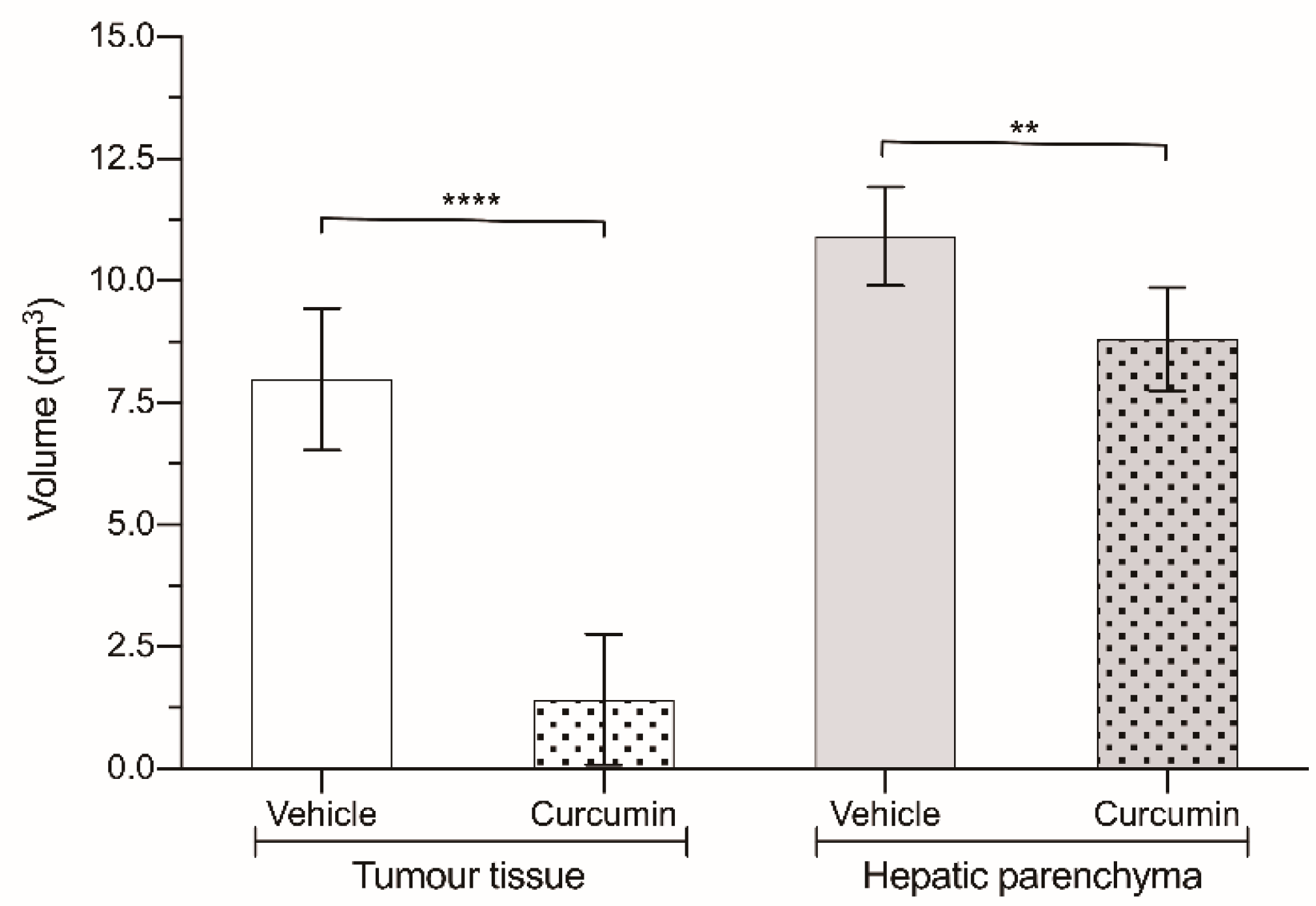
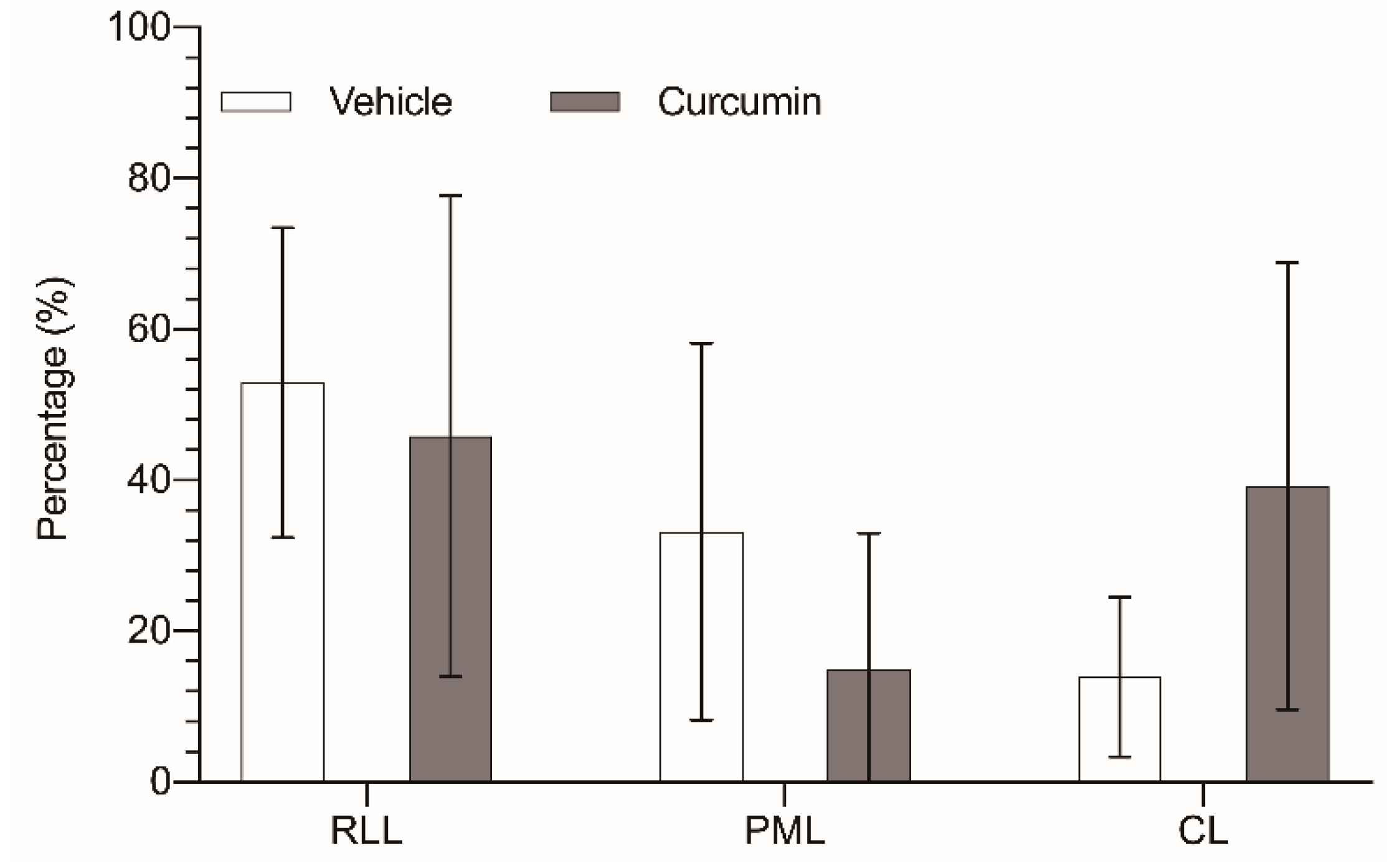
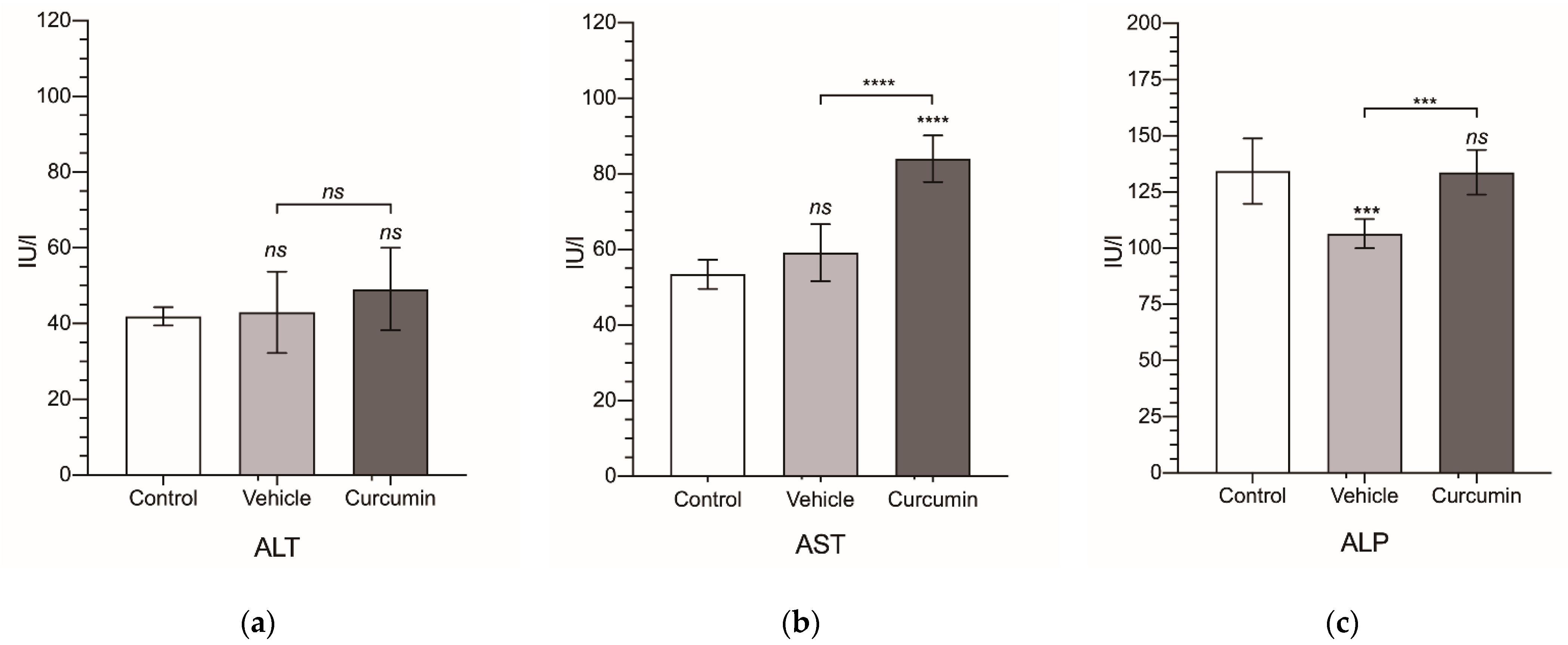
3.2. In Vivo Experiments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bahmani, M.; Shirzad, H.; Shahinfard, N.; Sheivandi, L.; Rafieian-Kopaei, M. Cancer Phytotherapy: Recent Views on the Role of Antioxidant and Angiogenesis Activities. J. Evid. Based Complementary Altern. Med. 2016, 22, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apaya, M.K.; Chang, M.-T.; Shyur, L.-F. Phytomedicine polypharmacology: Cancer therapy through modulating the tumor microenvironment and oxylipin dynamics. Pharmacol. Ther. 2016, 162, 58–68. [Google Scholar] [CrossRef]
- Hu, C.; Li, M.; Guo, T.; Wang, S.; Huang, W.; Yang, K.; Liao, Z.; Wang, J.; Zhang, F.; Wang, H. Anti-metastasis activity of curcumin against breast cancer via the inhibition of stem cell-like properties and EMT. Phytomedicine 2018, 58, 152740. [Google Scholar] [CrossRef]
- Fratantonio, D.; Molonia, M.S.; Bashllari, R.; Muscara’, C.; Ferlazzo, G.; Costa, G.; Saija, A.; Cimino, F.; Speciale, A. Curcumin potentiates the antitumor activity of Paclitaxel in rat glioma C6 cells. Phytomedicine 2018, 55, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, W.; Li, P.; Zheng, Z.; Tu, Y.; Zhang, Y.; You, T. Curcumin Enhances the Effects of 5-Fluorouracil and Oxaliplatin in Inducing Gastric Cancer Cell Apoptosis Both In Vitro and In Vivo. Oncol. Res. 2016, 23, 29–34. [Google Scholar] [CrossRef]
- Shakibaei, M.; Buhrmann, C.; Kraehe, P.; Shayan, P.; Lueders, C.; Goel, A. Curcumin Chemosensitizes 5-Fluorouracil Resistant MMR-Deficient Human Colon Cancer Cells in High Density Cultures. PLoS ONE 2014, 9, e85397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosieniak, G.; Adamowicz, M.; Alster, O.; Jaskowiak, H.; Szczepankiewicz, A.A.; Wilczynski, G.M.; Ciechomska, I.A.; Sikora, E. Curcumin induces permanent growth arrest of human colon cancer cells: Link between senescence and autophagy. Mech. Ageing Dev. 2012, 133, 444–455. [Google Scholar] [CrossRef]
- Chang, B.-D.; Broude, E.V.; Fang, J.; Kalinichenko, T.V.; Abdryashitov, R.; Poole, J.C.; Roninson, I.B. p21Waf1/Cip1/Sdi1-induced growth arrest is associated with depletion of mitosis-control proteins and leads to abnormal mitosis and endoreduplication in recovering cells. Oncogene 2000, 19, 2165–2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, J.L.; Hill, R.; Yaffe, P.B.; Greenshields, A.; Walsh, M.; Lee, P.W.; Giacomantonio, C.A.; Hoskin, D.W. Curcumin causes superoxide anion production and p53-independent apoptosis in human colon cancer cells. Cancer Lett. 2010, 297, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-C.; Lin, J.-G.; Li, T.-M.; Chung, J.-G.; Yang, J.-S.; Ip, S.-W.; Lin, W.-C.; Chen, G.-W. Curcumin-induced apoptosis of human colon cancer colo 205 cells through the production of ROS, Ca2+ and the activation of caspase-3. Anticancer. Res. 2007, 26, 4379–4389. [Google Scholar]
- Killian, P.H.; Kronski, E.; Michalik, K.M.; Barbieri, O.; Astigiano, S.; Sommerhoff, C.P.; Pfeffer, U.; Nerlich, A.G.; Bachmeier, B. Curcumin inhibits prostate cancer metastasis in vivo by targeting the inflammatory cytokines CXCL1 and -2. Carcinogenesis 2012, 33, 2507–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmansi, A.M.; El-Karef, A.A.; El-Shishtawy, M.M.; Eissa, L.A. Hepatoprotective Effect of Curcumin on Hepatocellular Carcinoma Through Autophagic and Apoptic Pathways. Ann. Hepatol. 2017, 16, 607–618. [Google Scholar] [CrossRef]
- Hong, J.H.; Ahn, K.S.; Bae, E.; Jeon, S.S.; Choi, H.Y. The effects of curcumin on the invasiveness of prostate cancer in vitro and in vivo. Prostate Cancer Prostatic Dis. 2006, 9, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Wang, Z.; Zhao, Y.; Wang, D.; Li, Y.; Ma, L.; Li, X.; Li, J.; Xiao, N.; Tian, J.; et al. Effects of curcumin on bladder cancer cells and development of urothelial tumors in a rat bladder carcinogenesis model. Cancer Lett. 2008, 264, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Zhang, J.; Zhang, J.; Miao, Q.; Zhang, J.; Yao, L. Curcumin promotes apoptosis by activating the p53-miR-192-5p/215-XIAP pathway in non-small cell lung cancer. Cancer Lett. 2015, 357, 196–205. [Google Scholar] [CrossRef]
- Li, W.; Jiang, L.; Lu, X.; Liu, X.; Ling, M. Curcumin protects radiation-induced liver damage in rats through the NF-κB signaling pathway. BMC Complement. Med. Ther. 2021, 21, 10. [Google Scholar] [CrossRef]
- Aboelella, N.; Brandle, C.; Kim, T.; Ding, Z.-C.; Zhou, G. Oxidative Stress in the Tumor Microenvironment and Its Relevance to Cancer Immunotherapy. Cancers 2021, 13, 986. [Google Scholar] [CrossRef]
- Wild, C.P.; Weiderpass, E.; Stewart, B.W. (Eds.) World Cancer Report: Cancer Research for Cancer Prevention; International Agency for Research on Cancer: Lyon, France, 2014; ISBN 9789283204299. [Google Scholar]
- Dekker, E.; Tanis, P.J.; A Vleugels, J.L.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- Van Der Geest, L.G.M.; Lam-Boer, J.; Koopman, M.; Verhoef, C.; Elferink, M.A.G.; De Wilt, J.H.W. Nationwide trends in incidence, treatment and survival of colorectal cancer patients with synchronous metastases. Clin. Exp. Metastasis 2015, 32, 457–465. [Google Scholar] [CrossRef]
- Bokemeyer, C.; Bondarenko, I.; Makhson, A.; Hartmann, J.T.; Aparicio, J.; de Braud, F.; Donea, S.; Ludwig, H.; Schuch, G.; Stroh, C.; et al. Fluorouracil, Leucovorin, and Oxaliplatin With and Without Cetuximab in the First-Line Treatment of Metastatic Colorectal Cancer. J. Clin. Oncol. 2009, 27, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Primrose, J.; Falk, S.; Finch-Jones, M.; Valle, J.; O’Reilly, D.; Siriwardena, A.; Hornbuckle, J.; Peterson, M.; Rees, M.; Iveson, T.; et al. Systemic chemotherapy with or without cetuximab in patients with resectable colorectal liver metastasis: The New EPOC randomised controlled trial. Lancet Oncol. 2014, 15, 601–611. [Google Scholar] [CrossRef]
- Carrato, A.; Abad, A.; Massuti, B.; Grávalos, C.; Escudero, P.; Longo-Muñoz, F.; Manzano, J.-L.; Gómez, A.; Safont, M.J.; Gallego, J.; et al. First-line panitumumab plus FOLFOX4 or FOLFIRI in colorectal cancer with multiple or unresectable liver metastases: A randomised, phase II trial (PLANET-TTD). Eur. J. Cancer 2017, 81, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Douillard, J.-Y.; Siena, S.; Cassidy, J.; Tabernero, J.; Burkes, R.; Barugel, M.; Humblet, Y.; Bodoky, G.; Cunningham, D.; Jassem, J.; et al. Randomized, Phase III Trial of Panitumumab With Infusional Fluorouracil, Leucovorin, and Oxaliplatin (FOLFOX4) Versus FOLFOX4 Alone As First-Line Treatment in Patients With Previously Untreated Metastatic Colorectal Cancer: The PRIME Study. J. Clin. Oncol. 2010, 28, 4697–4705. [Google Scholar] [CrossRef] [PubMed]
- Mellor, H.R.; Callaghan, R. Resistance to Chemotherapy in Cancer: A Complex and Integrated Cellular Response. Pharmacology 2008, 81, 275–300. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Xu, X.-Y.; Yunita, F.; Zhou, Q.; Wu, Y.-R.; Hu, Y.-X.; Wang, Z.-Q.; Tian, X.-F. Synergistic anti-liver cancer effects of curcumin and total ginsenosides. World J. Gastrointest. Oncol. 2020, 12, 1091–1103. [Google Scholar] [CrossRef]
- Ramachandran, C.; Fonseca, H.B.; Jhabvala, P.; Escalon, E.A.; Melnick, S.J. Curcumin inhibits telomerase activity through human telomerase reverse transcritpase in MCF-7 breast cancer cell line. Cancer Lett. 2002, 184, 1–6. [Google Scholar] [CrossRef]
- Chen, C.-H.; Chen, R.-J. Prevalence of Telomerase Activity in Human Cancer. J. Formos. Med Assoc. 2011, 110, 275–289. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Loo, W.T.; Sze, S.; Tong, Y. Curcumin inhibits cell proliferation of MDA-MB-231 and BT-483 breast cancer cells mediated by down-regulation of NFκB, cyclinD and MMP-1 transcription. Phytomedicine 2009, 16, 916–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-W.; Lee, J.-Y.; Huang, J.-Y.; Wang, C.-C.; Chen, W.-J.; Su, S.-F.; Huang, C.-W.; Ho, C.-C.; Chen, J.J.; Tsai, M.-F.; et al. Curcumin Inhibits Lung Cancer Cell Invasion and Metastasis through the Tumor Suppressor HLJ1. Cancer Res. 2008, 68, 7428–7438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, N.I.; Othman, I.; Abas, F.; Lajis, N.H.; Naidu, R. The Curcumin Analogue, MS13 (1,5-Bis(4-hydroxy-3-methoxyphenyl)-1,4-pentadiene-3-one), Inhibits Cell Proliferation and Induces Apoptosis in Primary and Metastatic Human Colon Cancer Cells. Molecules 2020, 25, 3798. [Google Scholar] [CrossRef]
- Sanaei, M.; Kavoosi, F. Effect of Curcumin and Trichostatin A on the Expression of DNA Methyltransfrase 1 in Hepatocellular Carcinoma Cell Line Hepa 1-6. Iran. J. Pediatr. Hematol. Oncol. 2018, 8, 193–201. [Google Scholar]
- Lee, S.J.; Krauthauser, C.; Maduskuie, V.; Fawcett, P.T.; Olson, J.M.; Rajasekaran, S.A. Curcumin-induced HDAC inhibition and attenuation of medulloblastoma growth in vitro and in vivo. BMC Cancer 2011, 11, 144. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Zhang, X.; Yu, P.; Chen, X.; Lu, P.; Li, M.; Liu, X.; Li, Z.; Wei, F.; Wang, K.; et al. Cinobufagin Induces Cell Cycle Arrest at the G2/M Phase and Promotes Apoptosis in Malignant Melanoma. Cells. Front. Oncol. 2019, 9, 853. [Google Scholar] [CrossRef] [Green Version]
- Weaver, B.A.; Cleveland, D.W. Decoding the links between mitosis, cancer, and chemotherapy: The mitotic checkpoint, adaptation, and cell death. Cancer Cell 2005, 8, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Liu, Y.-Y.; Ma, Y.-G.; Xue, Y.-X.; Liu, D.-G.; Ren, Y.; Liu, X.-B.; Li, Y.; Li, Z. Curcumin Blocks Small Cell Lung Cancer Cells Migration, Invasion, Angiogenesis, Cell Cycle and Neoplasia through Janus Kinase-STAT3 Signalling Pathway. PLoS ONE 2012, 7, e37960. [Google Scholar] [CrossRef]
- Su, J.; Zhou, X.; Wang, L.; Yin, X.; Wang, Z. Curcumin inhibits cell growth and invasion and induces apoptosis through down-regulation of Skp2 in pancreatic cancer cells. Am. J. Cancer Res. 2016, 6, 1949–1962. [Google Scholar]
- Zhang, Y.; Xue, Y.-B.; Li, H.; Qiu, D.; Wang, Z.-W.; Tan, S.-S. Inhibition of Cell Survival by Curcumin Is Associated with Downregulation of Cell Division Cycle 20 (Cdc20) in Pancreatic Cancer Cells. Nutrients 2017, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- Odot, J.; Albert, P.; Carlier, A.; Tarpin, M.; Devy, J.; Madoulet, C. In vitro and in vivo anti-tumoral effect of curcumin against melanoma cells. Int. J. Cancer 2004, 111, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.-Y.; Kim, D.-B.; Kim, E. Curcumin ameliorates the tumor-enhancing effects of a high-protein diet in an azoxymethane-induced mouse model of colon carcinogenesis. Nutr. Res. 2015, 35, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Diagaradjane, P.; Guha, S.; Deorukhkar, A.; Shentu, S.; Aggarwal, B.B.; Krishnan, S. Curcumin Sensitizes Human Colorectal Cancer Xenografts in Nude Mice to γ-Radiation by Targeting Nuclear Factor-κB–Regulated Gene Products. Clin. Cancer Res. 2008, 14, 2128–2136. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.L.; Norhaizan, M.E. Curcumin Combination Chemotherapy: The Implication and Efficacy in Cancer. Molecules 2019, 24, 2527. [Google Scholar] [CrossRef] [Green Version]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A Review of Curcumin and Its Derivatives as Anticancer Agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.M.; Fredman, E.T.; Coppa, C.; El-Gazzaz, G.; Aucejo, F.N.; Abdel-Wahab, M. Patterns of cancer recurrence in localized resected hepatocellular carcinoma. Hepatobiliary Pancreat. Dis. Int. 2015, 14, 269–274. [Google Scholar] [CrossRef]
- Cha, C.; Fong, Y.; Jarnagin, W.R.; Blumgart, L.H.; DeMatteo, R.P. Predictors and patterns of recurrence after resection of hepatocellular carcinoma. J. Am. Coll. Surg. 2003, 197, 753–758. [Google Scholar] [CrossRef]
- Matsumata, T.; Kanematsu, T.; Takenaka, K.; Yoshida, Y.; Nishizaki, T.; Sugimachi, K. Patterns of intrahepatic recurrence after curative resection of hepatocellular carcinoma. Hepatology 1989, 9, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Matkowskyj, K.A.; Marrero, J.A.; Carroll, R.E.; Danilkovich, A.V.; Green, R.M.; Benya, R.V. Azoxymethane-induced fulminant hepatic failure in C57BL/6J mice: Characterization of a new animal model. Am. J. Physiol. 1999, 277, G455–G462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobaek-Larsen, M.; Fenger, C.; Ritskes-Hoitinga, J. Secondary effects induced by the colon carcinogen azoxymethane in BDIX rats. APMIS 2004, 112, 319–329. [Google Scholar] [CrossRef]
- Mehendale, H.M. Tissue Repair: An Important Determinant of Final Outcome of Toxicant-Induced Injury. Toxicol. Pathol. 2005, 33, 41–51. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Mandal, D.P.; Sen, G.S.; Pal, S.; Banerjee, S.; Lahiry, L.; Finke, J.H.; Tannenbaum, C.S.; Das, T.; Sa, G. Tumor-Induced Oxidative Stress Perturbs Nuclear Factor-κB Activity-Augmenting Tumor Necrosis Factor-α–Mediated T-Cell Death: Protection by Curcumin. Cancer Res. 2007, 67, 362–370. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Yu, L.; Zhao, L.-Z. Curcumin up regulates T helper 1 cells in patients with colon cancer. Am. J. Transl. Res. 2017, 9, 1866–1875. [Google Scholar]
- Zou, J.Y.; Su, C.H.; Luo, H.H.; Lei, Y.Y.; Zeng, B.; Zhu, H.S.; Chen, Z.G. Curcumin converts Foxp3+ regulatory T cells to T helper 1 cells in patients with lung cancer. J. Cell. Biochem. 2017, 119, 1420–1428. [Google Scholar] [CrossRef]
- de la Parte, B.H.; González-Arribas, M.; Diaz-Sanz, I.; Palomares, T.; García-Alonso, I. Partial hepatectomy enhances the growth of CC531 rat colorectal cancer cells both in vitro and in vivo. Sci. Rep. 2021, 11, 5356. [Google Scholar] [CrossRef] [PubMed]
- Margonis, G.A.; Sasaki, K.; Andreatos, N.; Pour, M.Z.; Shao, N.; Ghasebeh, M.A.; Buettner, S.; Antoniou, E.; Wolfgang, C.L.; Weiss, M.; et al. Increased kinetic growth rate during late phase liver regeneration impacts the risk of tumor recurrence after colorectal liver metastases resection. HPB 2017, 19, 808–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manekeller, S.; Seinsche, A.; Sioutis, M.; Hirner, A. Extended liver resection after preoperative chemotherapy: Influence on regeneration and endoplasmic reticulum stress response. Langenbeck Arch. Surg. 2008, 394, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Nordlinger, B.; Sorbye, H.; Debois, M.; Praet, M.; Glimelius, B.; Poston, G.J.; Schlag, P.M.; Walpole, E.; Bechstein, W.; Gruenberger, T. Feasibility and risks of pre-operative chemotherapy (CT) with Folfox 4 and surgery for resectable colorectal cancer liver metastases (LM). Interim results of the EORTC Intergroup randomized phase III study 40983. J. Clin. Oncol. 2005, 23, 3528. [Google Scholar] [CrossRef]
- Vauthey, J.-N.; Pawlik, T.M.; Ribero, D.; Wu, T.-T.; Zorzi, D.; Hoff, P.M.; Xiong, H.Q.; Eng, C.; Lauwers, G.Y.; Mino-Kenudson, M.; et al. Chemotherapy Regimen Predicts Steatohepatitis and an Increase in 90-Day Mortality After Surgery for Hepatic Colorectal Metastases. J. Clin. Oncol. 2006, 24, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Seehofer, D.; Schirmeier, A.; Bengmark, S.; Carter, J.; Koch, M.; Glanemann, M.; Nussler, A.K.; Neuhaus, P.; Menger, M.D. Inhibitory Effect of Curcumin on Early Liver Regeneration Following Partial Hepatectomy in Rats. J. Surg. Res. 2009, 155, 195–200. [Google Scholar] [CrossRef]
- York, M. Clinical Pathology. In A Comprehensive Guide to Toxicology in Nonclinical Drug Development; Faqi, A.S., Ed.; Academic Press: London, UK, 2017; pp. 325–376. ISBN 978-0-12-803620-4. [Google Scholar]
- Vuppalanchi, R.; Chalasani, N. Laboratory Tests in Liver Disease. In Practical Hepatic Pathology: A Diagnostic Approach; Saxena, R., Ed.; Elsevier: Philadelphia, PA, USA, 2018; pp. 43–53. ISBN 978-0-323-42873-6. [Google Scholar]
- Van Putte, B.P.; Hendriks, J.M.; Romijn, S.; Pauwels, B.; Vermorken, J.B.; Van Schil, P.E. Combination chemotherapy with gemcitabine with isolated lung perfusion for the treatment of pulmonary metastases. J. Thorac. Cardiovasc. Surg. 2005, 130, 125–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herold, C.; Ocker, M.; Ganslmayer, M.; Gerauer, H.; Hahn, E.G.; Schuppan, D. Ciprofloxacin induces apoptosis and inhibits proliferation of human colorectal carcinoma cells. Br. J. Cancer 2002, 86, 443–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocker, M.; Herold, C.; Ganslmayer, M.; Hahn, E.G.; Schuppan, D. The synthetic retinoid adapalene inhibits proliferation and induces apoptosis in colorectal cancer cells in vitro. Int. J. Cancer 2003, 107, 453–459. [Google Scholar] [CrossRef]
- Maneikyte, J.; Bausys, A.; Leber, B.; Horvath, A.; Feldbacher, N.; Hoefler, G.; Strupas, K.; Stiegler, P.; Schemmer, P. Dietary glycine decreases both tumor volume and vascularization in a combined colorectal liver metastasis and chemotherapy model. Int. J. Biol. Sci. 2019, 15, 1582–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, S.B.; Procissi, D.; Chen, J.; Gogineni, V.R.; Tyler, P.; Yang, Y.; Omary, R.A.; Larson, A.C. Characterization of CC-531 as a Rat Model of Colorectal Liver Metastases. PLoS ONE 2016, 11, e0155334. [Google Scholar] [CrossRef] [PubMed]
- Heskamp, S.; Heijmen, L.; Gerrits, D.; Molkenboer-Kuenen, J.D.M.; ter Voert, E.; Heinzmann, K.; Honess, D.J.; Smith, D.-M.; Griffiths, J.R.; Doblas, S.; et al. Response Monitoring with [18F]FLT PET and Diffusion-Weighted MRI After Cytotoxic 5-FU Treatment in an Experimental Rat Model for Colorectal Liver Metastases. Mol. Imaging Biol. 2016, 19, 540–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.; Xu, W.; Kuang, M.; Gong, Y.; Cao, C.; Chen, J. Survival benefit and safety of the combinations of FOLFOXIRI ± bevacizumab versus the combinations of FOLFIRI ± bevacizumab as first-line treatment for unresectable metastatic colorectal cancer: A meta-analysis. OncoTargets Ther. 2016, 9, 4833–4842. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Fu, R. miR-206 regulates 5-FU resistance by targeting Bcl-2 in colon cancer cells. OncoTargets Ther. 2018, 11, 1757–1765. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Huang, S.; Wei, Y.; Cao, S.; Pi, C.; Feng, T.; Liang, J.; Zhao, L.; Ren, G. Curcumin Enhances the Anticancer Effect Of 5-fluorouracil against Gastric Cancer through Down-Regulation of COX-2 and NF-κB Signaling Pathways. J. Cancer 2017, 8, 3697–3706. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Shi, H.; Chen, C.; Ren, K.; Xu, Y.; Liu, X.; He, L. Curcumin enhances cisplatin sensitivity of human NSCLC cell lines through influencing Cu-Sp1-CTR1 regulatory loop. Phytomedicine 2018, 48, 51–61. [Google Scholar] [CrossRef]
- Falah, R.R.; Talib, W.H.; Shbailat, S.J. Combination of metformin and curcumin targets breast cancer in mice by angiogenesis inhibition, immune system modulation and induction of p53 independent apoptosis. Ther. Adv. Med Oncol. 2017, 9, 235–252. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Sesame Oil-Treated Animals | Curcumin-Treated Animals | ||||||
|---|---|---|---|---|---|---|---|---|
| Total | RLL | PML | CL | Total | RLL | PML | CL | |
| Liver parenchyma | 10.9 ± 1.01 | 3.76 ± 1.5 (34.5%) | 5.18 ± 1.4 (47.5%) | 1.97 ± 0.7 (18%) | 8.80 ± 1.06 | 3.69 ± 0.53 (42%) | 3.7 ± 0.89 (42%) | 1.41 ± 0.356 (16%) |
| Tumor tissue | 7.99 ± 1.45 | 4.30 ± 2.27 (53.8%) | 2.59 ± 2.06 (32.4%) | 1.1 ± 0.86 (13.8%) | 1.42 ± 1.34 | 0.67 ± 0.62 (47.2%) | 0.11 ± 0.14 (7.7%) | 0.64 ± 1.09 (45.1%) |
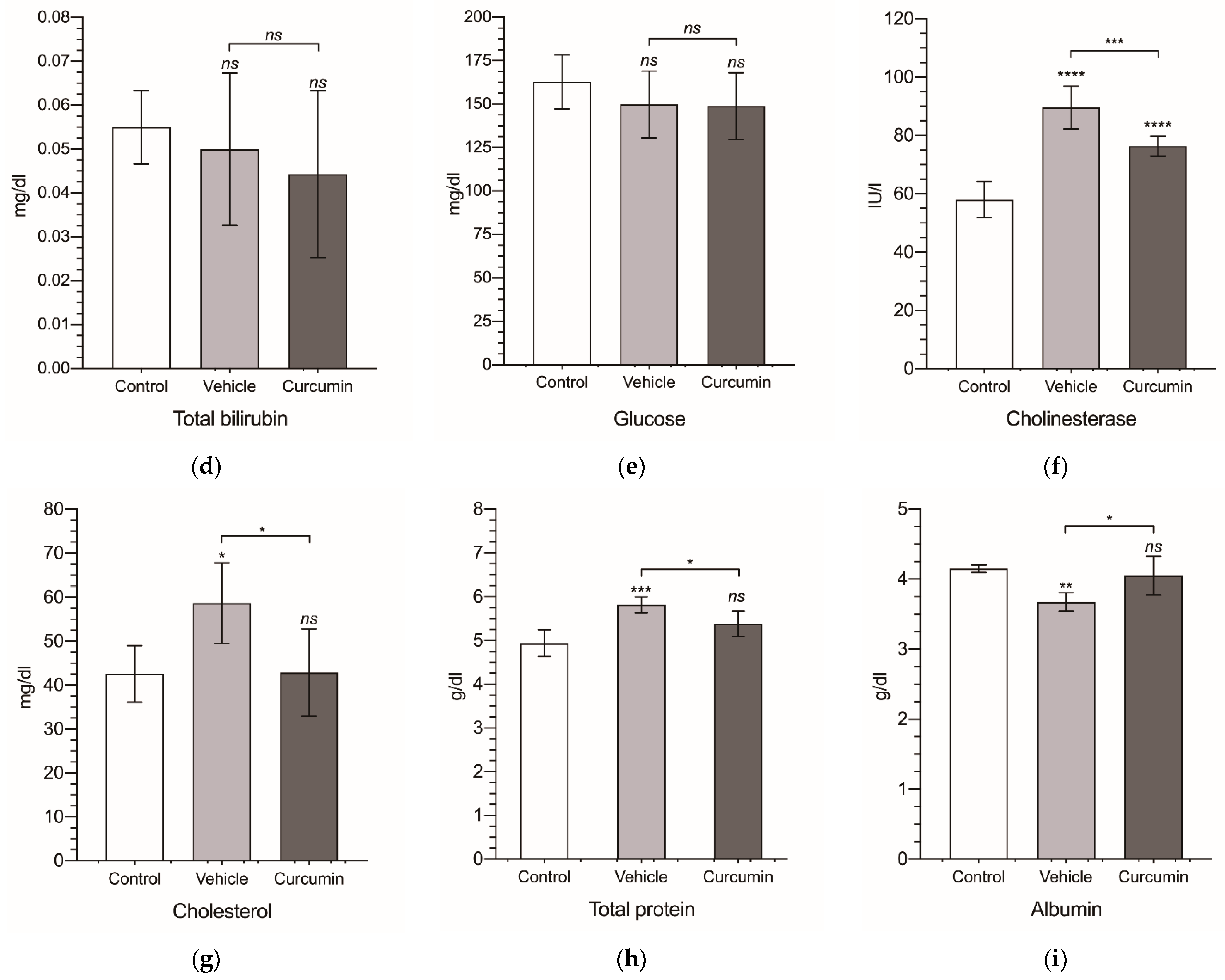
| Group | ALT | AST | ALP | TBil | Glucose | Cholinesterase | Cholesterol | TP | Albumin |
|---|---|---|---|---|---|---|---|---|---|
| Control | 41.9 ± 2.43 | 53.5 ± 3.83 | 134 ± 14.5 | 0.055 ± 0.008 | 163 ± 15.5 | 58.0 ± 6.24 | 42.6 ± 6.43 | 4.94 ± 0.305 | 4.15 ± 0.06 |
| Vehicle | 43.0 ± 10.7 | 59.2 ± 7.52 | 107 ± 6.47 | 0.05 ± 0.017 | 150 ± 19.2 | 89.6 ± 7.35 | 58.7 ± 9.14 | 5.82 ± 0.183 | 3.68 ± 0.13 |
| Curcumin | 49.1 ± 10.9 | 84.0 ± 6.13 | 134 ± 9.95 | 0.044 ± 0.019 | 149 ± 19.1 | 76.4 ± 3.38 | 42.9 ± 9.94 | 5.39 ± 0.291 | 4.05 ± 0.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrero de la Parte, B.; Rodeño-Casado, M.; Iturrizaga Correcher, S.; Mar Medina, C.; García-Alonso, I. Curcumin Reduces Colorectal Cancer Cell Proliferation and Migration and Slows In Vivo Growth of Liver Metastases in Rats. Biomedicines 2021, 9, 1183. https://doi.org/10.3390/biomedicines9091183
Herrero de la Parte B, Rodeño-Casado M, Iturrizaga Correcher S, Mar Medina C, García-Alonso I. Curcumin Reduces Colorectal Cancer Cell Proliferation and Migration and Slows In Vivo Growth of Liver Metastases in Rats. Biomedicines. 2021; 9(9):1183. https://doi.org/10.3390/biomedicines9091183
Chicago/Turabian StyleHerrero de la Parte, Borja, Mikel Rodeño-Casado, Sira Iturrizaga Correcher, Carmen Mar Medina, and Ignacio García-Alonso. 2021. "Curcumin Reduces Colorectal Cancer Cell Proliferation and Migration and Slows In Vivo Growth of Liver Metastases in Rats" Biomedicines 9, no. 9: 1183. https://doi.org/10.3390/biomedicines9091183
APA StyleHerrero de la Parte, B., Rodeño-Casado, M., Iturrizaga Correcher, S., Mar Medina, C., & García-Alonso, I. (2021). Curcumin Reduces Colorectal Cancer Cell Proliferation and Migration and Slows In Vivo Growth of Liver Metastases in Rats. Biomedicines, 9(9), 1183. https://doi.org/10.3390/biomedicines9091183