
Time-Series Monitoring of Transgenic Maize Seedlings Phenotyping Exhibiting Glyphosate Tolerance


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Setup
2.2. Acquisition of Hyperspectral Imaging In Vivo
2.3. Measurement of ChlF In Vivo
2.4. A Brief Information of OJIP Fluorescence Transient and JIP-Test
2.5. Measurement of Leaf Chlorophyll Content
2.6. Data Analysis and Model Development
3. Results
3.1. Establishment of LCC Prediction Model Based on Spectral Reflectance
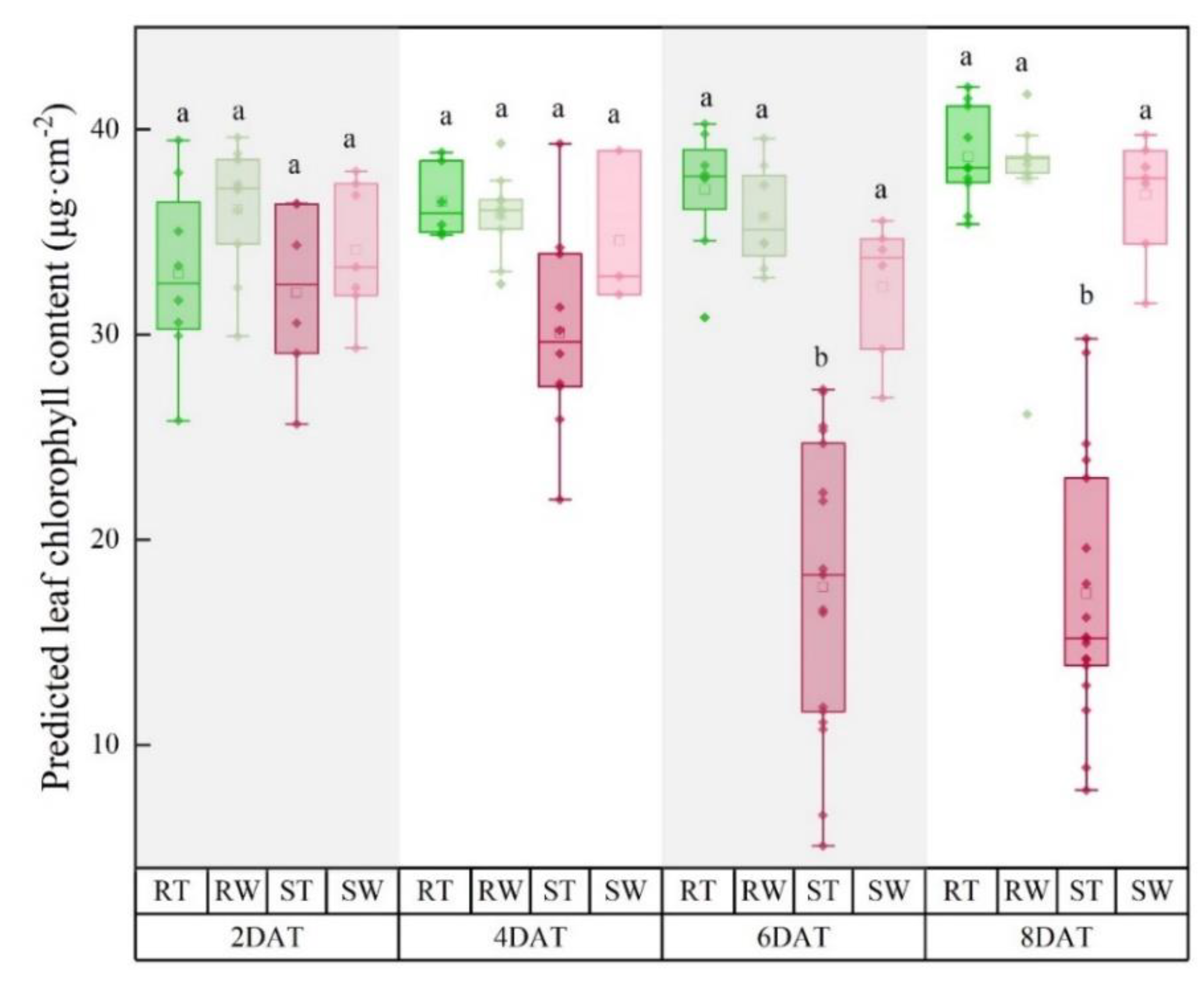
3.2. Effect of Glyphosate Stress on Pigment Content within Leaves of Maize Seedlings
3.3. OJIP Transient Captures the Difference Responses to Glyphosate Stress
3.4. Effect of Glyphosate Stress on PSII and PSI by JIP-Test
3.5. JIP-Test Parameters Selection for Responses of Sensitive Genotype to Glyphosate
4. Discussion
4.1. Robustness of LCC Prediction Model
4.2. Glyphosate Treatment Effects on Photosynthetic Physiology
4.3. Potential Applications and Future Prospect
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef] [PubMed]
- De Freitas-Silva, L.; de Araújo, T.O.; Nunes-Nesi, A.; Ribeiro, C.; Costa, A.C.; da Silva, L.C. Evaluation of morphological and metabolic responses to glyphosate exposure in two neotropical plant species. Ecol. Indic. 2020, 113, 106246. [Google Scholar] [CrossRef]
- Beckie, H.J. Herbicide-resistant weed management: Focus on glyphosate. Pest Manag. Sci. 2011, 67, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Dou, T.; Krimmer, M.; Singh, S.; Humpal, D.; Payne, W.Z.; Sanchez, L.; Voronine, D.V.; Prosvirin, A.; Scully, M.; et al. Raman Spectroscopy Can Distinguish Glyphosate-Susceptible and -Resistant Palmer Amaranth (Amaranthus palmeri). Front. Plant Sci. 2021, 12, 657963. [Google Scholar] [CrossRef]
- Bloem, E.; Gerighausen, H.; Chen, X.; Schnug, E. The potential of spectral measurements for identifying glyphosate application to agricultural fields. Agronomy 2020, 10, 1409. [Google Scholar] [CrossRef]
- Vahtmäe, E.; Kotta, J.; Orav-Kotta, H.; Kotta, I.; Pärnoja, M.; Kutser, T. Predicting macroalgal pigments (Chlorophyll a, chlorophyll b, chlorophyll a + b, carotenoids) in various environmental conditions using highresolution hyperspectral spectroradiometers. Int. J. Remote Sens. 2018, 39, 5716–5738. [Google Scholar] [CrossRef]
- Kuckenberg, J.; Tartachnyk, I.; Noga, G. Temporal and spatial changes of chlorophyll fluorescence as a basis for early and precise detection of leaf rust and powdery mildew infections in wheat leaves. Precis. Agric. 2009, 10, 34–44. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Rastogi, A.; Živčák, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypiński, P.; Samborska, I.A.; et al. Prompt chlorophyll fluorescence as a tool for crop phenotyping: An example of barley landraces exposed to various abiotic stress factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Yang, J.; Zhang, M.; Strasser, R.J.; Qiang, S. Classification and characteristics of heat tolerance in Ageratina adenophora populations using fast chlorophyll a fluorescence rise O-J-I-P. Environ. Exp. Bot. 2016, 122, 126–140. [Google Scholar] [CrossRef]
- Stirbet, A.; Lazár, D.; Kromdijk, J. Govindjee Chlorophyll a fluorescence induction: Can just a one-second measurement be used to quantify abiotic stress responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, Y.; Cong, Y.; Zhu, P.; Xing, J.; Cui, J.; Xu, W.; Shi, Q.; Diao, M.; Liu, H.Y. Ascorbic Acid-Induced Photosynthetic Adaptability of Processing Tomatoes to Salt Stress Probed by Fast OJIP Fluorescence Rise. Front. Plant Sci. 2021, 12, 594400. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Račková, L.; Paganová, V.; Swoczyna, T.; Rusinowski, S.; Sitko, K. Can chlorophyll-a fluorescence parameters be used as bio-indicators to distinguish between drought and salinity stress in Tilia cordata Mill? Environ. Exp. Bot. 2018, 152, 149–157. [Google Scholar] [CrossRef]
- Zhang, T.; Huang, Y.; Reddy, K.N.; Yang, P.; Zhao, X.; Zhang, J. Using machine learning and hyperspectral images to assess damages to corn plant caused by glyphosate and to evaluate recoverability. Agronomy 2021, 11, 583. [Google Scholar] [CrossRef]
- Feng, X.; Zhao, Y.; Zhang, C.; Cheng, P.; He, Y. Discrimination of transgenic maize kernel using NIR hyperspectral imaging and multivariate data analysis. Sensors 2017, 17, 1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaji, H.M.; Oukarroum, A.; Alexandrov, V.; Kouzmanova, M.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Allakhverdiev, S.I.; Goltsev, V. Identification of nutrient deficiency in maize and tomato plants by invivo chlorophyll a fluorescence measurements. Plant Physiol. Biochem. 2014, 81, 16–25. [Google Scholar] [CrossRef]
- Feng, X.; Yu, C.; Chen, Y.; Peng, J.; Ye, L.; Shen, T.; Wen, H.; He, Y. Non-destructive determination of shikimic acid concentration in transgenic maize exhibiting glyphosate tolerance using chlorophyll fluorescence and hyperspectral imaging. Front. Plant Sci. 2018, 9, 468. [Google Scholar] [CrossRef]
- Neubauer, C.; Schreiber, U. The polyphasic rise of chlorophyll fluorescence upon onset of strong continuous illumination: I. saturation characteristics and partial control by the photosystem II acceptor side. Z. Für Nat. C 1987, 42, 1246–1254. [Google Scholar] [CrossRef] [Green Version]
- Tsimilli-Michael, M. Revisiting JIP-test: An educative review on concepts, assumptions, approximations, definitions and terminology. Photosynthetica 2020, 58, 275–292. [Google Scholar] [CrossRef] [Green Version]
- Spyroglou, I.; Rybka, K.; Rodriguez, R.M.; Stefański, P.; Valasevich, N.M. Quantitative estimation of water status in field-grown wheat using beta mixed regression modelling based on fast chlorophyll fluorescence transients: A method for drought tolerance estimation. J. Agron. Crop Sci. 2021, 207, 589–605. [Google Scholar] [CrossRef]
- Zhang, J.; Wan, L.; Igathinathane, C.; Zhang, Z.; Guo, Y.; Sun, D.; Cen, H. Spatiotemporal Heterogeneity of Chlorophyll Content and Fluorescence Response Within Rice (Oryza sativa L.) Canopies Under Different Nitrogen Treatments. Front. Plant Sci. 2021, 12, 499. [Google Scholar] [CrossRef]
- Wan, L.; Cen, H.; Zhu, J.; Zhang, J.; Zhu, Y.; Sun, D.; Du, X.; Zhai, L.; Weng, H.; Li, Y.; et al. Grain yield prediction of rice using multi-temporal UAV-based RGB and multispectral images and model transfer—A case study of small farmlands in the South of China. Agric. For. Meteorol. 2020, 291, 108096. [Google Scholar] [CrossRef]
- Feudale, R.N.; Woody, N.A.; Tan, H.; Myles, A.J.; Brown, S.D.; Ferré, J. Transfer of multivariate calibration models: A review. Chemom. Intell. Lab. Syst. 2002, 64, 181–192. [Google Scholar] [CrossRef]
- Weng, H.; Lv, J.; Cen, H.; He, M.; Zeng, Y.; Hua, S.; Li, H.; Meng, Y.; Fang, H.; He, Y. Hyperspectral reflectance imaging combined with carbohydrate metabolism analysis for diagnosis of citrus Huanglongbing in different seasons and cultivars. Sens. Actuators B Chem. 2018, 275, 50–60. [Google Scholar] [CrossRef]
- Fan, L.; Feng, Y.; Weaver, D.B.; Delaney, D.P.; Wehtje, G.R.; Wang, G. Glyphosate effects on symbiotic nitrogen fixation in glyphosate-resistant soybean. Appl. Soil Ecol. 2017, 121, 11–19. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Zhu, Y.; Xu, H.; He, Y.; Cen, H. Time-series chlorophyll fluorescence imaging reveals dynamic photosynthetic fingerprints of sos mutants to drought stress. Sensors 2019, 19, 2649. [Google Scholar] [CrossRef] [Green Version]
- Magyar, M.; Sipka, G.; Kovács, L.; Ughy, B.; Zhu, Q.; Han, G.; Špunda, V.; Lambrev, P.H.; Shen, J.R.; Garab, G. Rate-limiting steps in the dark-tolight transition of photosystem ii-revealed by chlorophyll-a fluorescence induction. Sci. Rep. 2018, 8, 2755. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, M.A.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli-Michael, M.; Govindjee; Sarin, N.B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1428–1438. [Google Scholar] [CrossRef] [Green Version]
- Oukarroum, A.; Schansker, G.; Strasser, R.J. Drought stress effects on photosystem i content and photosystem II thermotolerance analyzed using Chl a fluorescence kinetics in barley varieties differing in their drought tolerance. Physiol. Plant. 2009, 137, 188–199. [Google Scholar] [CrossRef]
- Jiang, H.X.; Chen, L.S.; Zheng, J.G.; Han, S.; Tang, N.; Smith, B.R. Aluminum-induced effects on Photosystem II photochemistry in Citrus leaves assessed by the chlorophyll a fluorescence transient. Tree Physiol. 2008, 28, 1863–1871. [Google Scholar] [CrossRef]
- Mlinarić, S.; Dunić, J.A.; Babojelić, M.S.; Cesar, V.; Lepeduš, H. Differential accumulation of photosynthetic proteins regulates diurnal photochemical adjustments of PSII in common fig (Ficus carica L.) leaves. J. Plant Physiol. 2017, 209, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.J. Donor side capacity of Photosystem II probed by chlorophyll a fluorescence transients. Photosynth. Res. 1997, 52, 147–155. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Bąba, W.; Gediga, K.; Goltsev, V.; Samborska, I.A.; Cetner, M.D.; Dimitrova, S.; Piszcz, U.; Bielecki, K.; Karmowska, K.; et al. Chlorophyll fluorescence as a tool for nutrient status identification in rapeseed plants. Photosynth. Res. 2018, 136, 329–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambreen, S.; Athar, H.-R.; Khan, A.; Zafar, Z.U.; Ayyaz, A.; Kalaji, H.M. Seed priming with proline improved photosystem II efficiency and growth of wheat (Triticum aestivum L.). BMC Plant Biol. 2021, 21, 502. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Ghosh, R.; Strasser, R.J.; Ponpandian, L.N.; Bae, H. Chlorophyll-a fluorescence evaluation of PEG-induced osmotic stress on PSII activity in Arabidopsis plants expressing SIP1. Plant Biosyst. 2018, 152, 945–952. [Google Scholar] [CrossRef]
- Derks, A.; Schaven, K.; Bruce, D. Diverse mechanisms for photoprotection in photosynthesis. Dynamic regulation of photosystem II excitation in response to rapid environmental change. Biochim. Biophys. Acta Bioenerg. 2015, 1847, 468–485. [Google Scholar] [CrossRef] [Green Version]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of photosynthesis during abiotic stress-induced photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [Green Version]
- Zeng, F.; Wang, G.; Liang, Y.; Guo, N.; Zhu, L.; Wang, Q.; Chen, H.; Ma, D.; Wang, J. Disentangling the photosynthesis performance in japonica rice during natural leaf senescence using OJIP fluorescence transient analysis. Funct. Plant Biol. 2021, 48, 206–217. [Google Scholar] [CrossRef]
- Bano, H.; Athar, H.-u.-R.; Zafar, Z.U.; Kalaji, H.M.; Ashraf, M. Linking changes in chlorophyll a fluorescence with drought stress susceptibility in mung bean [Vigna radiata (L.) Wilczek]. Physiol. Plant. 2021, 172, 1244–1254. [Google Scholar] [CrossRef]
- Zobiole, L.H.S.; Kremer, R.J.; Oliveira, R.S.; Constantin, J. Glyphosate affects chlorophyll, nodulation and nutrient accumulation of “second generation” glyphosate-resistant soybean (Glycine max L.). Pestic. Biochem. Physiol. 2011, 99, 53–60. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formula and Terms | Explanation |
|---|---|
| Parameters extracted from the recorded fluorescence transient OJIP | |
| F0 ≈ F20μs | Fluorescence when all PSII RCs are open |
| FM (=FP) | Maximal fluorescence when all PSII RCs are closed |
| FL ≈ F100μs | Fluorescence at 100 μs |
| FK ≈ F300μs | Fluorescence at 300 μs |
| FJ = F2ms | Fluorescence at the J-step (2 ms) of OJIP |
| FI = F30ms | Fluorescence at the I-step (30 ms) of OJIP |
| Time (in ms) to reach the maximal fluorescence FM | |
| Parameters derived from extracted fluorescence data | |
| Area | Total complementary area between fluorescence induction curve and F = FM |
| Sm | Area/tFM |
| Fv = FM − F0 | Maximal variable fluorescence |
| Fv/F0 | Efficiency of the water-splitting complex on the donor side of PSII |
| Vt = (Ft − F0)/(FM − F0) | Relative variable fluorescence at time t (normalization on FM − F0) |
| W(YZ), t = (Ft − FY)/(FZ − FY) | Different types of relative variable fluorescence at time t, with (FZ − FY) standing for (F300µs − F0) or (FJ − F0) or (FI − FJ) or (FI − F0) or (FP − FI) |
| M0 = [(△F/△t)0]/(FM − F0) = 4 × (F300µs − F20µs)/(FM − F0) = 4 × (V300µs − V20µs) | Approximated initial slope (in ms−1) of the fluorescence transient normalized on the maximal variable fluorescence FM − F0; equivalently, initial slope (20 to 300 µs; in ms−1) of the Vt = f(t) kinetics |
| Specific energy fluxes per QA− reducing PSII reaction center (RC) and per cross section (CS) at t = 0 | |
| ABS/RC = M0·(1/VJ)/(1 − F0/Fm) | Absorption flux per reaction center (RC) by exciting PSII antenna Chlorophyll molecules |
| TR0/RC = M0·(1/VJ) | Trapping energy flux for QA reduction at PSII reaction center (RC) |
| ET0/RC = M0·(1/VJ)·(1 − VJ) | Electron transport flux per active reaction center (RC) |
| RE0/RC = M0·(1/VJ)·(1 − VI) | Electron flux reducing end electron acceptors at the photosystem I (PSI) acceptor side per reaction center (RC) |
| ABS/CS0 ≈ F0 | Absorption flux per cross section (CS) by exciting PSII antenna Chlorophyll molecules |
| TR0/CS0 = φP0·F0 | Trapping energy flux for QA reduction per cross section (CS) |
| ET0/CS0 = φE0·F0 | Electron transport flux per cross section (CS) |
| RE0/CS0 = φE0·φR0·F0 | Electron flux reducing end electron acceptors at the photosystem I (PSI) acceptor side per cross section (CS) |
| Quantum yields and probabilities/efficiencies | |
| φP0 = TR0/ABS = 1 − F0/Fm | Maximum quantum yield of Photosystem II primary photochemistry in the dark-adapted state |
| ψE0 = ET0/TR0 = 1 − VJ | Probability that a trapped exciton moves an electron into the electron chain beyond QA− |
| φE0 = ET0/ABS = (1 − F0/Fm)·(1 − VJ) | Quantum yield for electron transport |
| δR0 = RE0/ET0 = (1 − VI)/(1 − VJ) | Efficiency/probability of electron transfer from the PQ pool, beyond QA− and reduction of end electron acceptors in PSI |
| φR0 = RE0/ABS = (1 − F0/Fm)·(1 − VI) | Quantum yield for the reduction of the end acceptors of PSI per photon absorbed |
| Other biophysical parameters | |
| γRC = RC/(ABS + RC) | Probability that a PSII Chlorophyll a molecule functions as RC |
| RC/CS0 = φP0 − (VJ/M0)·(ABS/CS) | Density of RCs (QA− reducing PSII reactive centers) |
| Performance indices | |
| PIABS = (RC/ABS)·(φP0/(1 − φP0))·(ψE0/(1 − ψE0)) | Performance index for energy conservation from photos absorbed by PSII until the reduction of intersystem electron acceptors |
| Sampling Point | Selected JIP–Test Parameters |
|---|---|
| 2 DAT | VJ, ψE0, M0, ET0/CS0, RC/CS0, φE0 |
| 4 DAT | M0, φE0, PIABS, RE0/CS0, VJ, ψE0 |
| 6 DAT | φE0, M0, VJ, ψE0, ET0/CS0, RE0/CS0 |
| 8 DAT | M0, φE0, γRC, VJ, ψE0, ABS/RC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, M.; Bai, X.; Zhang, J.; Wei, Y.; He, Y. Time-Series Monitoring of Transgenic Maize Seedlings Phenotyping Exhibiting Glyphosate Tolerance. Processes 2022, 10, 2206. https://doi.org/10.3390/pr10112206
Tao M, Bai X, Zhang J, Wei Y, He Y. Time-Series Monitoring of Transgenic Maize Seedlings Phenotyping Exhibiting Glyphosate Tolerance. Processes. 2022; 10(11):2206. https://doi.org/10.3390/pr10112206
Chicago/Turabian StyleTao, Mingzhu, Xiulin Bai, Jinnuo Zhang, Yuzhen Wei, and Yong He. 2022. "Time-Series Monitoring of Transgenic Maize Seedlings Phenotyping Exhibiting Glyphosate Tolerance" Processes 10, no. 11: 2206. https://doi.org/10.3390/pr10112206