Abstract
The cell wall, serving as the exoskeleton of plants, is naturally a barrier to resist external stresses. Protoplasts can be obtained by dissolving the cell walls of plant cells without damaging the cell membrane, and are widely used in the rapid propagation, transgenic breeding, and somatic hybridization of plants. However, to regenerate the cell wall is a precondition for cell division. Therefore, to study the culture condition and influencing factors during the cell wall regeneration of protoplasts is vital. Traditionally, culture medium is used to cultivate protoplasts, but it has some disadvantages. Herein, a microfluidic system with crossed channels was constructed to isolate and cultivate the protoplasts of tobacco. Then, the cell wall regeneration of the tobacco protoplasts was also studied based on this microfluidic system. It was found that, compared with the control, benzo-(1, 2, 3)-thiadiazole-7-carbothioic acid S-methyl ester (BTH) could accelerate the regeneration of the cell wall, while Pseudomonas syringae pv. tomato DC3000 (Pst DC3000) could inhibit the regeneration of the cell wall within 24 h. To conclude, this study demonstrated that a crossed microfluidic chip could be an effective tool to study cell wall regeneration or other behavior of plant cells in situ with high resolution. In addition, this study revealed the rate of cell wall regeneration under BTH and Pst DC3000 treatment.
1. Introduction
The plant cell wall is mainly composed of cellulose, hemicellulose, pectin, and lignin, and represents one of the most intricate network structures in nature [1]. It plays vital roles in determining plants’ various shapes and mechanical strengths, and acts as a first tier within the defense system in response to external damage [1,2,3,4,5]. The protoplast is derived from the plant cell whose cell wall is dissolved by enzymes without damaging the cell membrane, and it has been widely used in the rapid propagation, transgenic breeding, and somatic hybridization of plants [6,7]. However, it is worth noting that the cell wall regeneration of a protoplast is a precondition for cell division. Therefore, to understand the culture environment and influencing factors of the cell wall regeneration for more types of plants is extremely important. As a matter of fact, many studies have proposed that there are some major factors that influence the regeneration of cell walls in various plants [8,9]. By consulting these examples from the literature, we found that the use of culture mediums is the most common method to culture the protoplasts, but this method still has some disadvantages; for instance, cells are prone to adhesion, and it is hard to observe the cell growth and development in situ and in real-time [6]. Therefore, to propose a new platform for observing the process of the cell wall regeneration of protoplasts is essential.
Microfluidic technology is a micro-electromechanical technology that can accurately control and manipulate microfluids [10,11]. It is characterized of micro-crossed channels, high integration, high throughput, and easy observation, significantly improving the temporal and spatial resolution. Therefore, the microfluidic devices have been used to study plant cells and organs [12,13,14,15,16,17]. For instance, Grossmann et al. described a microfluidic chip platform to monitor the time-resolved growth and cytosolic sugar levels of Arabidopsis roots at cellular and subcellular resolution and under controlled environmental conditions [12]. Based on microfluidic and microelectromechanical systems technology, Agudelo et al. developed the TipChip to study pollen tube growth under precisely controlled and reproducible conditions, instead using of other time-consuming approaches [13]. Wu et al. used the microfluidic device to present the culture of tobacco protoplasts and their fusion [16]. Furthermore, the microfluidic platform was also widely used in single cell RNA sequencing to study cell-specific gene expression in animals and plants [18,19,20]. In all, the microfluidic technology has been widely used for various types of bioassays and imaging in both plant and animal cells, but it has not been used to dynamically monitor the regeneration of cell wall in situ.
In this study, a crossed microfluidic channel was constructed to isolate and cultivate tobacco protoplasts. Then, the tobacco protoplasts were treated with benzo-(1, 2, 3)-thiadiazole-7-carbothioic acid S-methyl ester (BTH) and Pseudomonas syringae pv. tomato DC3000 (Pst DC3000) to study the regeneration of cell wall based on this microfluidic channel. The results showed that this microfluidic device provides a powerful tool to monitor the plant cell behaviour in situ with high resolution, which will help us to study the culture environment and influencing factors of the cell wall regeneration and, furthermore, to promote the studies of recombination for nuclear and cytoplasmic genes and somatic embryogenesis [6].
2. Materials and Methods
2.1. Reagents and Materials
Polydimethylsiloxane (PDMS) Sylgard 184 was purchased from Dow Corning (Midland, MI, USA). Additionally, SU-8 GM1075 photoresist was purchased from the Gersteltec (Vaud, Switzerland). Silicon chips with a thickness of 360 μm were bought from Suzhou Double Metal Materials, Co., Ltd., Suzhou, China, while 1 mL syringes were bought from Nantong’s Biological Experiment Equipment Co., Ltd., Nantong, China. Cellulase, pectase, and calcium fluoride white were purchased from the Yuanye Biological Co., Ltd., Shanghai, China. The BTH was purchased from Sigma-Aldrich, Saint Louis, MO, USA. Other reagents were of analytical grade.
2.2. Microchip Fabrication
A crossed microfluidic channel was constructed according to our previous report [21]. Briefly, the crossed microfluidic channel with a width of 250 µm, depth of 60 µm, and length of 30 mm (5–6 mm crossed zone) was designed by drawing software (CAD) and printed on transparent film (Scheme 1A). The transparent film was used as a photomask for coating the silicon wafer covered with SU-8 photoresist, which was exposed to EXFO OmniCurc® S1000 ultraviolet point light to form the crossed microchannels. Then, PDMS Sylgard 184 (mixing in the ratio of 10:1) was poured onto the mold (the silicon wafer surface with the consolidate channels) and heated (Scheme 1B). After baking, three holes were punched on the back of the glue using corresponding punchers with diameters of 1, 1.5, and 2.5 mm, which produced the ports for injection of the protoplasts that were suspended in solution (1), injection of the treatment solution (2), and outlet for the solution (3), respectively (Scheme 1C). The structured PDMS component with the crossed microfluidic channel and a glass slide were firmly pasted together. Scheme 1D showed the images which were taken using a Nikon Ti-DH fluorescent inverted microscope (Tokyo, Japan). Teflon tubing was connected to the inlets of the channels for transporting the solution by a Langer LSP02-1B double-channel microsyringe pump (Nanjing, China).
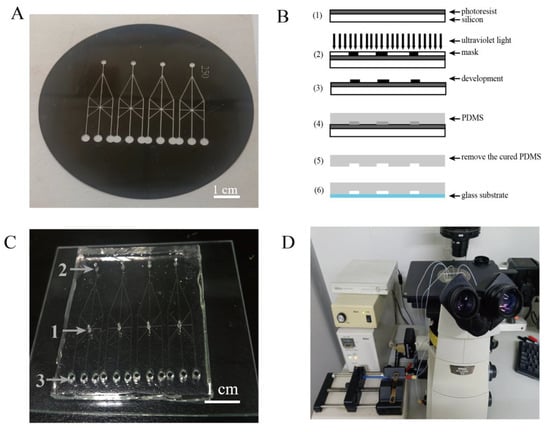
Scheme 1.
The fabrication process of the crossed microfluidic chip and imaging system. (A) The transparent film for the crossed microchannels. (B) The fabrication process of the crossed microfluidic chip. (C) The crossed microfluidic chip as follows: (1) the site for injection of protoplasts, (2) the site for injection of treatment solution, (3) outlet for solution. (D) Observation through the fluorescence microscope system.
2.3. Extraction Process of Tobacco Protoplasts
To obtain the protoplasts, Sun’s method was employed [22]. Briefly, the seeds of tobacco were planted in 1/2 MS culture medium in a climate-controlled incubator with a 14 h light/10 h dark photoperiod at 26 °C/24 °C for 6 weeks. The sterile leaves of tobacco were selected and cut into samples with a width of 1 mm. The samples of leaves were put into the sterilized NP solution (15 mg/mL cellulose, 4 mg/mL pectase, 4 mM morpholineethanesulfonic acid, 0.8 M CaCl2·2H2O, 2 M KCl, and 0.5 M mannitol, pH 5.8) for enzymolysis at 25 °C about 5 h. After that, the solution containing protoplasts was centrifuged for 6 min at 150 rcf. Then, the precipitated protoplasts were resuspended by using the modified W5 solution (0.16 M NaCl, 0.09 M CaCl2·2H2O, 0.5 mM KCl, and 4 mM morpholineethanesulfonic acid, pH 5.8), and washed three times by centrifugation to discard the enzyme. Finally, the protoplasts were resuspended using NP that contained 0.25 mg/L kinetin and 2 mg/L 2, 4-dichlorophenoxyacetic acid (2, 4-D).
2.4. The Separation and Cultivation of the Protoplasts in the Crossed Microfluidic Channels
To realize the separation and cultivation of the protoplasts, the 20 μL protoplast solution was injected into the crossed microchip channels from inlet 1 with a microsyringe pump at a low flow rate of 0.5 µL/min for 40 min. Then, the crossed microchip channel with the protoplasts was kept in a sterile petri dish (diameter 13 cm) and cultured in a climate-controlled incubator with a 0 h light/24 h dark photoperiod at 26 °C. The humidity in the sterile petri dish was maintained through the addition of sterile water. Then, the crossed microchip channel with the protoplasts was observed and imaged at different time points in situ.
2.5. The Observation of the Cell Wall Regeneration
In order to study the cell wall regeneration of the protoplasts, 20 μL of protoplast solution, containing the 0.1% calcium fluoride white and the solution of the treatments, was injected simultaneously from inlet 1 and inlet 2, respectively. Specifically, for the BTH treatment, the 20 μL 1.36% BTH solution was injected from inlet 2. For the Pst DC3000 treatment, the 20 μL suspension of the Pst DC3000 with the OD 600 value of 0.2 was injected from inlet 2. For the control, a similar volume of NP solution was injected from inlet 2. Then, the cell wall regeneration of protoplasts was observed and imaged under the fluorescence inverted microscope at different time points in situ.
2.6. Analysis of the Regeneration Ratio of the Plant Cell Wall
The protoplasts display as red under ultraviolet light because the chloroplasts can produce the red fluorescence. Calcium fluoride white (CFW) can combine with the cellulose and chitosan of the plant cell wall to produce bluish violet (excitation and emission wavelengths at 345 and 430 nm, respectively). Therefore, the red protoplasts represented no cell wall and the bluish violet protoplasts represented regenerated walls under the fluorescence microscope, respectively. Five random microscopic fields of the protoplasts in the crossed microfluidic chip were selected to count the number of red and bluish violet protoplasts. Then, the proportion, i.e., the number of bluish violet protoplasts/(the number of red and bluish violet protoplasts), was calculated and, thus, the regeneration ratio of the plant cell wall was obtained.
2.7. Statistical Analysis
All the experimental data collected were statistically analyzed by one-way analysis of variance followed by a least significant difference (LSD) test at p < 0.05 or p < 0.01 using the IBM SPSS software version 19.0 (New York, NY, USA). Standard errors of the means from three independent biological replicates were calculated.
3. Results and Discussion
3.1. Separation and Cultivation of Protoplasts in Crossed Microfluidic Channels
To realize the separation, cultivation, and continuous monitoring of protoplasts in situ, a crossed microfluidic device was constructed. The protoplasts were injected into the crossed microchip channels with a microsyringe pump at a low flow rate of 0.5 µL/min for 40 min to disperse the protoplasts in the microchip channels. As shown in Figure 1, the protoplasts with a diameter of 20–30 µm were dispersed into crossed channels. The integrity and activity of protoplasts within the 48 h continuous culture suggested that the crossed microchip channels are suited to the separation, cultivation, and continuous observation of protoplasts in situ. It should be emphasized that, compared with the single channel, the crossed microchannel of our study can supply more opportunities for protoplasts to choose, which will help to separate and obtain a single protoplast.

Figure 1.
The separation and cultivation of protoplasts in crossed microfluidic channels at 0 h (A), 24 h (B), and 48 h (C).
3.2. Regeneration of Cell Wall under BTH Treatment
Here, BTH, as a salicylic acid analogue, can widely induce systemic acquired resistance (SAR), which provides long-lasting and broad-spectrum protection at the whole plant level [23,24,25,26]. As shown in Figure 2A,B, the protoplasts fluoresce red due to the irradiation of the chloroplasts with ultraviolet light. The regeneration of the cell wall led to the appearance of a bluish white stain produced by the CFW stain of the cell wall (Figure 2d,f). Compared with the control (Figure 2c), the bluish violet appearance of the protoplasts resulted from the regeneration of the cell wall using the BTH treatment at 12 h (Figure 2d). The cell wall of the protoplasts regenerated more rapidly when treated with BTH compared to the control, which suggested that the BTH can facilitate the regeneration of the cell wall (Video S1). The regeneration rates of the cell walls treated with BTH were higher than the control (Figure 3A). In addition, almost all of the protoplasts had regenerated the cell wall using the BTH treatment at 24 h. Our results also agreed well with previous reports on the effects of BTH, namely that it can induce cell wall strengthening and the deposition of lignin in calluses [27,28,29,30]. The possible reason was that the BTH affected the composition of carbohydrates and down-regulated the abundance of cell structure-related proteins to strengthen the cell wall, which can accelerate the regeneration of the cell wall [29,30]. Furthermore, the dynamic processes of protoplast cell wall regeneration within 7 h after treatment with BTH were also continuously monitored (Video S1). These results demonstrated that the crossed microfluidic channels could be used to observe the regeneration of the cell wall in situ.
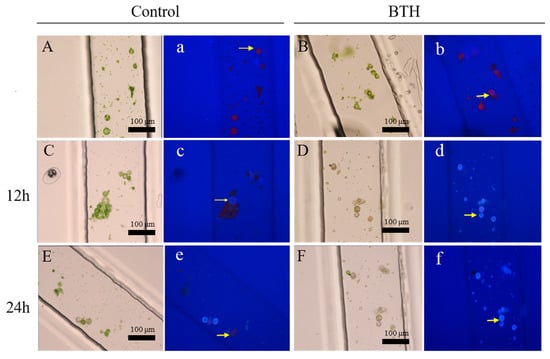
Figure 2.
The cell wall regeneration of protoplasts in microchannels treated with benzo-(1, 2, 3)-thiadiazole-7-carbothioic acid S-methyl ester (BTH). (A,C,E) show the cell wall regeneration of protoplasts treated by the control under the white light; a, c, and e show the cell wall regeneration of protoplasts treated by the control under the fluorescent light. (B,D,F) show the cell wall regeneration of protoplasts treated with the BTH under the white light; b, d, and f show the cell wall regeneration of protoplasts treated with the BTH under the fluorescent light. Protoplasts of (A) (a) and (B) (b) at 0 h, (C) (c) and (D) (d) at 12 h, and (E) (e) and (F) (f) at 24 h.
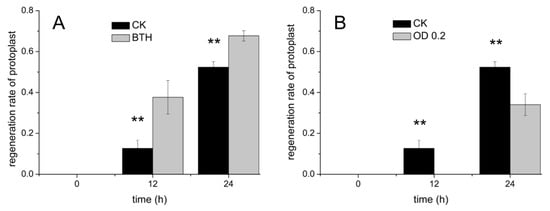
Figure 3.
(A) The cell wall regeneration rates of protoplasts treated with BTH. (B) The cell wall regeneration rates of protoplasts treated with Pseudomonas syringae pv. tomato DC3000 (Pst DC3000). An asterisk (*) represents a significant difference (* p < 0.05, ** p < 0.01).
3.3. Regeneration of Cell Wall under the PstDC3000 Treatment
The Pst DC3000 treatment, as a pattern pathogenesis, has been used to understand the interaction mechanism between pathogenesis and plants [31,32]. Here, Pst DC3000 with an OD value of 0.2 was injected and cultured in the microfluidic chip. As shown in Figure 4, the rod-shaped Pst DC3000, with the length of 5–8 µm, were dispersed into cross channels. There was no significant variation in the amount and activation of the Pst DC3000 during the 24 h culture. The interaction between the Pst DC3000 and protoplasts was also observed in situ. As shown in Figure 5, the protoplasts were surrounded by Pst DC3000, which suggested that both of them could be co-cultured in the microfluidic chip. In addition, constant moving, colliding, and infecting of protoplasts by Pst DC3000 was observed in the microfluidic chip (Video S2). The regeneration of the cell wall for the protoplasts treated by Pst DC3000 was significantly lower than the control within 24 h and especially at 12 h, there was almost no regeneration of the cell wall (Figure 3B and Figure 6). Such results suggested that the Pst DC3000 could inhibit the regeneration of cell wall at the cell level. The previous studies suggested that Pst DC3000 might enhance its infection by manipulating the IDL6-HAE/HSL2-ADPG2 signaling pathway, which could promote pectin degradation in Arabidopsis leaves [33]. Compared to the prior study [33], our results provided direct evidence for the Pst DC3000 infection of the plants as evidenced by the inhibition of cell wall regeneration, which is a novel discovery for the Pst DC3000 infection events at the cellular level.
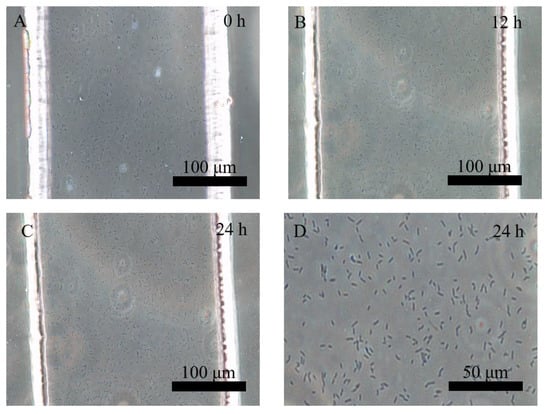
Figure 4.
The separation and cultivation of Pst DC3000 in crossed microfluidic channels at 0 h (A), 12 h (B), and 24 h (C), and the magnified image of Pst DC3000 at 24 h (D).
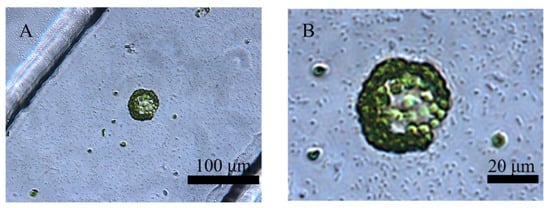
Figure 5.
The co-cultivation of a protoplast and Pst DC3000 in the crossed microfluidic channels (A), and the magnified image (B).
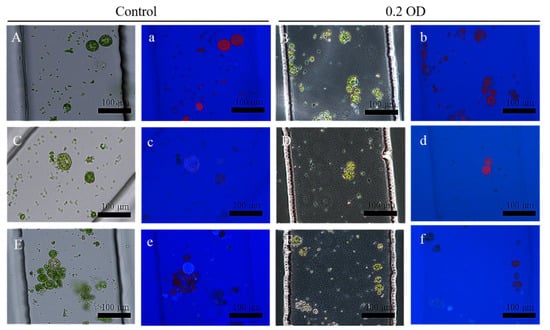
Figure 6.
The cell wall regeneration of protoplasts in microchannels treated with Pst DC3000. (A,C,E) show the cell wall regeneration of protoplasts treated with the control under the white light; (a,c,e) show the cell wall regeneration of protoplasts treated with the control under the fluorescent light. (B,D,F) show the cell wall regeneration of protoplasts treated with Pst DC3000 under white light. (b,d,f) show the cell wall regeneration of protoplasts treated with Pst DC3000 under the fluorescent light. Protoplasts of (A) (a) and (B) (b) at 0 h, (C) (c) and (D) (d) at 12 h, and (E) (e) and (F) (f) at 24 h.
4. Conclusions
The cell wall regeneration of protoplasts is a precondition for cell division. Therefore, the observed dynamics of the protoplast cell wall regeneration is critically important for studying the culture environment and influencing factors during the processes of cell wall regeneration. In this study, a microfluidic system with crossed channels was constructed to isolate and cultivate tobacco protoplasts. More importantly, it was firstly used to study the cell wall regeneration of protoplasts in situ with high resolution. The results also showed that BTH could accelerate the regeneration of the cell wall, while Pst DC3000 inhibited the regeneration of the cell wall within 24 h. Overall, our results demonstrated that the crossed microfluidic device provides a powerful tool to monitor the plant cell behavior and the interaction between plant cells and plant pathogens, which will help us to explore the interaction mechanisms of plant and pathogens at the level of a single cell.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/pr10122507/s1: Video S1, Dynamic processes of the cell wall regeneration of protoplasts within 7 h after treatment with BTH; Video S2, Dynamic processes of the cell wall regeneration of protoplasts within two minutes after treatment with Pst DC3000.
Author Contributions
Conceptualization by L.S., S.X. and Z.S.; methodology by S.X. and Z.S.; validation by L.L.; formal analysis by Y.Y.; investigation by S.Z.; resources by Y.L. and N.B.; data curation by S.X. and Z.S.; writing—original draft preparation by S.X. and Y.Z.; writing—review and editing by Y.Z. and L.S.; visualization by S.X. and Z.S.; supervision by L.S.; project administration by Y.Z.; funding acquisition by L.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China (grant numbers: 31770399 and 21375066), and the project of the Nantong Natural Science Foundation (grant numbers: MS22021038 and MS12021012).
Data Availability Statement
Not applicable.
Conflicts of Interest
There are no conflicts to declare.
References
- Zhang, B.; Gao, Y.; Zhang, L.; Zhou, Y. The plant cell wall: Biosynthesis, construction, and functions. J. Integr. Plant Biol. 2021, 63, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Vaahtera, L.; Schulz, J.; Hamann, T. Cell wall integrity maintenance during plant development and interaction with the environment. Nat. Plants 2019, 5, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Lin, W.; Zhou, X.; Guo, J.; Dang, X.; Li, B.; Lin, D.; Yang, Z. Mechano-transduction via the pectin-FERONIA complex activates ROP6 GTPase signaling in Arabidopsis pavement cell morphogenesis. Curr. Biol. 2022, 32, 508–517. [Google Scholar] [CrossRef]
- Bacete, L.; Mélida, H.; Miedes, E.; Molina, A. Plant cell wall-mediated immunity: Cell wall changes trigger disease resistance responses. Plant J. 2018, 93, 614–636. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, Z.; Wang, Y.; Wang, J.; Xiao, M.; Liu, H.; Quan, R.; Zhang, H.; Huang, R.; Zhu, L.; et al. Cellulose synthase-like protein OsCSLD4 plays an important role in the response of rice to salt stress by mediating abscisic acid biosynthesis to regulate osmotic stress tolerance. Plant Biotechnol. J. 2022, 20, 468–484. [Google Scholar] [CrossRef]
- Zhang, Q.; Wei, W.; Zhang, H.; Gu, L. Research progress on regeneration of cell wall from forest tree protoplasts. Mol. Plant Breed. 2021. Available online: http://kns.cnki.net/kcms/detail/46.1068.S.20210430.1320.006.html (accessed on 1 November 2022).
- Sahab, S.; Hayden, M.J.; Mason, J.; Spangenberg, G. Mesophyll Protoplasts and PEG-Mediated Transfections: Transient Assays and Generation of Stable Transgenic Canola Plants. In Transgenic Plants; Kumar, S., Barone, P., Smith, M., Eds.; Humana Press: New York, NY, USA, 2019; Volume 1864, pp. 131–132. [Google Scholar]
- Borgato, L.; Pisani, F.; Furini, A. Plant regeneration from leaf protoplasts of Solanum virginianum L. (Solanaceae). Plant Cell Tissue Organ Cult. 2007, 88, 247–252. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Piwowarczyk, B. Studies on cell wall regeneration in protoplast culture of legumes–the effect of organic medium additives on cell wall components. Czech. J. Genet. Plant Breed. 2014, 50, 84–91. [Google Scholar] [CrossRef]
- Lucchetta, E.M.; Lee, J.H.; Fu, L.A.; Patel, N.H.; Ismagilov, R.F. Dynamics of Drosophila embryonic patterning network perturbed in space and time using microfluidics. Nature 2005, 434, 1134–1138. [Google Scholar] [CrossRef]
- Badri, A.M.; Coenen, K.; Vaillancourt, L.P.; Goulet, C.; Michaud, D. On-chip detection of low-molecular-weight recombinant proteins in plant crude extracts by SELDI-TOF MS. Methods Mol. Biol. 2009, 483, 313–324. [Google Scholar]
- Grossmann, G.; Guo, W.J.; Ehrhardt, D.W.; Frommer, W.B.; Sit, R.V.; Quake, S.R.; Meier, M. The RootChip: An integrated microfluidic chip for plant science. Plant Cell 2011, 23, 4234–4240. [Google Scholar] [CrossRef]
- Agudelo, C.G.; Sanati, N.A.; Ghanbari, M.; Naghavi, M.; Packirisamy, M.; Geitmann, A. TipChip: A modular, MEMS-based platform for experimentation and phenotyping of tip-growing cells. Plant J. 2013, 73, 1057–1068. [Google Scholar] [CrossRef]
- Sanati, N.A. Microfluidic platforms for plant cells studies. Lab Chip 2014, 14, 3262–3274. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, N.; Kozgunova, E.; Grossmann, G.; Geitmann, A.; Higashiyama, T. Microfluidics-Based Bioassays and Imaging of Plant Cells. Plant Cell Physiol. 2021, 62, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Liu, W.; Tu, Q.; Song, N.; Li, L.; Wang, J.; Wang, J. Culture and chemical-induced fusion of tabacco mesophy11 protoplasts in a microfluidic device. Microfluid. Nanofluid. 2011, 10, 867–876. [Google Scholar] [CrossRef]
- Yu, Z.; Boehm, C.R.; Hibberd, J.M.; Abell, C.; Haseloff, J.; Burgess, S.J.; Reyna-Llorens, I. Droplet-based microfluidic analysis and screening of single plant cells. PLoS ONE 2018, 13, e0196810. [Google Scholar] [CrossRef] [PubMed]
- Ryu, K.H.; Huang, L.; Kang, H.M.; Schiefelbein, J. Single-Cell RNA Sequencing Resolves Molecular Relationships Among Individual Plant Cells. Plant Physiol. 2019, 179, 1444–1456. [Google Scholar] [CrossRef]
- Li, X.; Lee, A.P. Chapter 2-high-throughput microfluidic single-cell trapping arrays for biomolecular and imaging analysis. Method Cell Biol. 2018, 148, 35–50. [Google Scholar]
- Xu, X.; Wang, J.X.; Wu, L.L.; Guo, J.J.; Song, Y.L.; Tian, T.; Wang, W.; Zhu, Z.; Yang, C.Y. Microfluidic Single-Cell Omics Analysis. Small 2020, 16, 1903905. [Google Scholar] [CrossRef]
- Sun, L.; Liu, L.; Lin, X.; Xia, Z.; Cao, J.; Xu, S.; Gu, H.; Yang, H.; Bao, N. Microfluidic Devices for Monitoring the Root Morphology of Arabidopsis Thaliana in situ. Anal. Sci. 2021, 37, 605–611. [Google Scholar] [CrossRef]
- Sun, Y.; Xue, Q.; Ding, C.; Zhang, X.; Zhang, L.; Wang, W.; Ali, S. Somatic cybridization between Nicotiana tabacum and N. repanda based on a single inactivation procedure of nuclear donor parental protoplasts. Plant Sci. 2005, 168, 303–308. [Google Scholar] [CrossRef]
- Gao, H.; Guo, M.; Song, J.; Ma, Y.; Xu, Z. Signals in systemic acquired resistance of plants against microbial pathogens. Mol. Biol. Rep. 2021, 48, 3747–3759. [Google Scholar] [CrossRef] [PubMed]
- Lawton, K.A.; Friedrich, L.; Hunt, M.; Weymann, K.; Delaney, T.; Kessmann, H.; Staub, T.; Ryals, J. Benzothiadiazole induces disease resistance in Arabidopsis by activation of the systemic acquired resistance signal transduction pathway. Plant J. 1996, 10, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Shine, M.B.; Xiao, X.; Kachroo, P.; Kachroo, A. Signaling mechanisms underlying systemic acquired resistance to microbial pathogens. Plant Sci. 2019, 279, 81–86. [Google Scholar] [CrossRef]
- Cooper, B.; Beard, H.S.; Garrett, W.M.; Campbell, K.B. Benzothiadiazole Conditions the Bean Proteome for Immunity to Bean Rust. Mol. Plant-Microbe Interact. 2020, 33, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Burlini, N.; Iriti, M.; Daghetti, A.; Faoro, F.; Ruggiero, A.; Bernasconi, S. Benzothiadiazole (BTH) activates sterol pathway and affects vitamin D3 metabolism in Solanum malacoxylon cell cultures. Plant Cell Rep. 2011, 30, 2131–2141. [Google Scholar] [CrossRef]
- Barilli, E.; Rubiales, D.; Amalfitano, C.; Evidente, A.; Prats, E. BTH and BABA induce resistance in pea against rust (Uromyces pisi) involving differential phytoalexin accumulation. Planta 2015, 242, 1095–1106. [Google Scholar] [CrossRef]
- Apolinar-Valiente, R.; Ruiz-García, Y.; Williams, P.; Gil-Muñoz, R.; Gómez-Plaza, E.; Doco, T. Preharvest Application of Elicitors to Monastrell Grapes: Impact on Wine Polysaccharide and Oligosaccharide Composition. J. Agric. Food Chem. 2018, 66, 11151–11157. [Google Scholar] [CrossRef]
- Li, X.; Bi, Y.; Wang, J.; Dong, B.; Li, H.; Gong, D.; Zhao, Y.; Tang, Y.; Yu, X.; Shang, Q. BTH treatment caused physiological, biochemical and proteomic changes of muskmelon (Cucumis melo L.) fruit during ripening. J. Proteom. 2015, 120, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.F.; He, S.Y. Pseudomonas syringae pv. tomato DC3000: A model pathogen for probing disease susceptibility and hormone signaling in plants. Annu. Rev. Phytopathol. 2013, 51, 473–498. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Cai, G.; Jiang, S.; Sun, L.; Li, D. Response of tobacco to the Pseudomonas syringae pv. tomato DC3000 is mainly dependent on salicylic acid signaling pathway. FEMS Microbiol. Lett. 2013, 344, 77–85. [Google Scholar] [CrossRef]
- Wang, X.; Hou, S.; Wu, Q.; Lin, M.; Acharya, B.R.; Wu, D.; Zhang, W. IDL6-HAE/HSL2 impacts pectin degradation and resistance to Pseudomonas syringae pv. tomato DC3000 in Arabidopsis leaves. Plant J. 2017, 89, 250–263. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).