1. Introduction
Biotechnological processes have been found to be suitable and low-cost options for the removal of organic and inorganic contaminants in wastewaters [
1,
2,
3]. Phenol, phenolic derivatives and their mixtures are among the extremely toxic pollutants arising from industrial effluents. Although sodium salicylate (SA) is used as a drug derivative in medicine and as preservative in foods production, it is recently qualified as a typical contaminant in wastewater due to its high level toxicity (cf., for example, [
4] and the references therein). The phenol/sodium salicylate mixture is found in wastewater from various industries (chemical, pharmaceutical, cosmetic and others). For that reason, modern technologies for removal of toxic compounds from industrial and pharmaceutical wastewater are constantly being developed [
5,
6,
7]. The availability of clean water is important for ensuring human health, societal development and environmental sustainability. Wastewater must be treated before being released into the environment or reused according to international and national regulatory requirements, emphasized in many European documents [
8,
9].
Recently, biodegradation of phenol and its derivatives, as well as of SA is successfully carried out with various specific microorganisms such as
Trichosporoncutaneum,
Arthrobacter,
Pseudomonas putida,
Gliomastix indicus,
Aspergillus awamori,
Trametes hirsute,
Rhodococcus,
Burkholderia,
Candida tropicalis and many others [
10,
11,
12,
13,
14,
15,
16]. The experimental work is performed mainly at laboratory scales using a chemostat as a part of apparatus. It seems that the name “chemostat” appears for the first time in [
17]. The chemostat is also known as “continuous culture” and “continuously stirred tank reactor” (CSTR). Using the chemostat, numerous mathematical models have been developed in different areas of natural sciences and bioengineering [
18,
19,
20].
Usually, the Haldane kinetic model describes the specific cell growth rates on a single substrate in wastewater treatment models. However, the specific cell growth rates in a substrate mixture of two and more pollutant components are expressed by complex nonlinear functions [
12,
21]. These functions are presented as sums or products of modified kinetic models to take into account the mutual influence between the substrates on their biodegradation rate by the so-called interaction coefficients. The latter account for the inhibition of the degradation of one substrate in the presence of the other or of the binary mixture as a whole [
14,
16]. The following models are widely used in the analysis and control of the wastewater treatment processes involving mixtures of pollutant components: sum kinetics with interaction parameters (SKIP), self-inhibition EC-SKIP (SIEC-SKIP), elimination capacity-sum kinetics with interaction parameter (EC-SKIP), etc. [
22,
23,
24,
25]. The SKIP models describe well the biodegradation by different microorganisms of various mixtures of interacting phenolic pollutants in wastewater: phenol and p-cresol or phenol and resorcinol by
Gliomastix indicus MTCC 3869 [
12]; phenol and SA by
Pseudomonas putida [
14,
15,
16]; 4-bromophenol and 4-chlorophenol by
Arthrobacter chlorophenolicus A6 [
26].
Controlling a biotechnological process is a delicate and not easy task. This is due to the complexity of the process, involving a variety of living microorganisms which dynamics is often unstable and not well known. Model-based control is used to predict the behavior of the bioreactor systems and is gaining an increased importance in recent decades. The controller type depends on many factors such as the knowledge of the system, availability and complexity of the considered model, etc. Among the classical controllers are the proportional-integral (PI) controller, the proportional integral-differential (PID) controller, the adaptive PID and the cascade PI controls; all they have been recognized as a good alternative for the regulation of the plants (cf. [
27,
28] and the references therein). Other recently developed approaches for controlling continuous bioreactors are nonlinear adaptive control [
29,
30], feedback control [
31], extremum seeking control [
32,
33,
34]. More detailed information about instrumentation and control of biotechnological processes can be found in the review paper [
35].
A significant characteristic of chemostat cultivation is the dilution rate
D. In practice,
D is defined as the flow of medium per time over the volume of the culture in the reactor and can be directly manipulated by the experimenter. For that reason a large number of studies is devoted to investigating the effect of
D on the long-term behavior of the chemostat dynamics. Among the rich literature we can mention e.g., the papers [
36,
37] and the references therein, as well as the books [
18,
19]. Using
D as a control parameter is considered in [
38,
39] and applied to a CSTR model for simultaneous degradation of phenol and p-cresol in industrial wastewater.
Biodegradation of phenol and SA mixture by the strain
Pseudomonas putida (P. putida) CCRC 14365 is reported in [
14,
15], where series of batch tests are conducted and used to determine the interaction parameters in the kinetic growth models. The obtained results show that the cells preferably degrade phenol than SA.
The high biodegradation rate of phenol and SA by
P. putida 49451 is established in details by Lin and Ho in [
16]. Based on eight batch tests, the kinetic parameters are determined by comparing the model-fitted specific growth rates with that ones of the experimental results. Experimental results show that the addition of SA to phenol does not significantly affect the time required for complete biodegradation of phenol. However, the presence even of a small amount of phenol accelerates the complete biodegradation of SA. Moreover, the authors present in their paper for the first time a continuous-time (chemostat) model for biodegradation of the mixture by
P. putida 49451. They use two chemostats to evaluate the biodegradation of phenol and SA with different initial conditions. It is shown that the experimental results in the chemostat system fit very well with the predicted values of the model for a particular value of the dilution rate
. All results in [
16] are also discussed and compared with other experimental data on phenol and SA by
P. putida given in [
12,
14,
15].
Here we consider the chemostat model for biodegradation of phenol and SA mixture by the strain
P. putida 49451 proposed by Lin and Ho in [
16]. As already mentioned before, only a quantitative verification of the dynamics at a particular value of the dilution rate
has been carried out in the latter paper. Till now this model has not yet been investigated qualitatively. Our paper aims to perform a detailed mathematical analysis of the model solutions.
The mathematical analysis is based on the theory of autonomous dynamical systems, described by nonlinear ordinary differential equations [
18,
40]. The latter offers a rich arsenal of techniques and methods, which are recently widely used in mathematical modelling of real-life processes. Based on this theory, the objectives of our study are to (i) determine bounds (interval) for the dilution rate
D and to establish existence of model equilibrium points within these bounds; (ii) investigate the local asymptotic stability of the equilibria; (iii) establish existence, uniqueness and boundedness of positive model solutions; (iv) prove global stabilizability of the dynamics towards a prescribed equilibrium point by using
D as a control function. The obtained theoretical results provide a good framework for practical applications. They can be used in the design of effective and sustainable management of the biodegradation process of phenol and SA mixture in wastewater.
The paper is structured in the following way.
Section 2 shortly presents the mathematical model for biodegradation of phenol and SA mixture by the
P. putida cells, given in [
16]. The main results are reported in
Section 3 and
Section 4.
Section 3 is devoted to local stability analysis of the model, including computation of the equilibrium points as well as investigation of their local asymptotic stability with respect to the parameter
D.
Section 4 reports on general and important properties of the model solutions and provides results on the global stabilizability of the system.
Section 5 presents numerical examples as illustration of the theoretical studies on the model dynamics. The last
Section 6 discusses the presented theoretical results and points out their importance and practical applicability.
2. The Chemostat Model
The chemostat model for biodegradation of the binary mixture of phenol and sodium salicylate (SA) by the strain
Pseudomonas putida 49451 is described by the following system of nonlinear ordinary differential equations [
16]
where
and
are the specific cell growth rates on phenol and SA respectively, presented by the following analytical expressions [
12,
14,
16]
The meaning of the state variables
,
,
X and of the model parameters is summarized in
Table 1. The numerical values in the last column are taken from [
16], where they are obtained and verified by laboratory experiments.
In our study we assume that the influent concentrations of phenol () and SA () are constant. The dilution rate D is considered as a control function in the model.
The specific growth rates
and
represent the so called SKIP (Sum Kinetics with Interaction Parameters) models of cell growth, which as shown in [
16], give the best fit to the experimental results of phenol and SA biodegradation. Each one of
respectively
contains two interaction parameters,
and
, respectively
and
. The considerably grater numerical value of
compared to
(see last column in
Table 1) indicates that SA shows higher uncompetitive inhibition on phenol biodegradation in comparison to that of phenol on SA biodegradation. The value of
in
is also larger that the value of
in
, which is indicative for the fact that the inhibition of phenol biodegradation by SA is higher than the inhibition of SA biodegradation by phenol. These phenomena have also been experimentally validated, see e.g., [
14,
16] and the references therein. Obviously, if
, respectively
then
, respectively
represent the Haldane growth (Halling type IV) function.
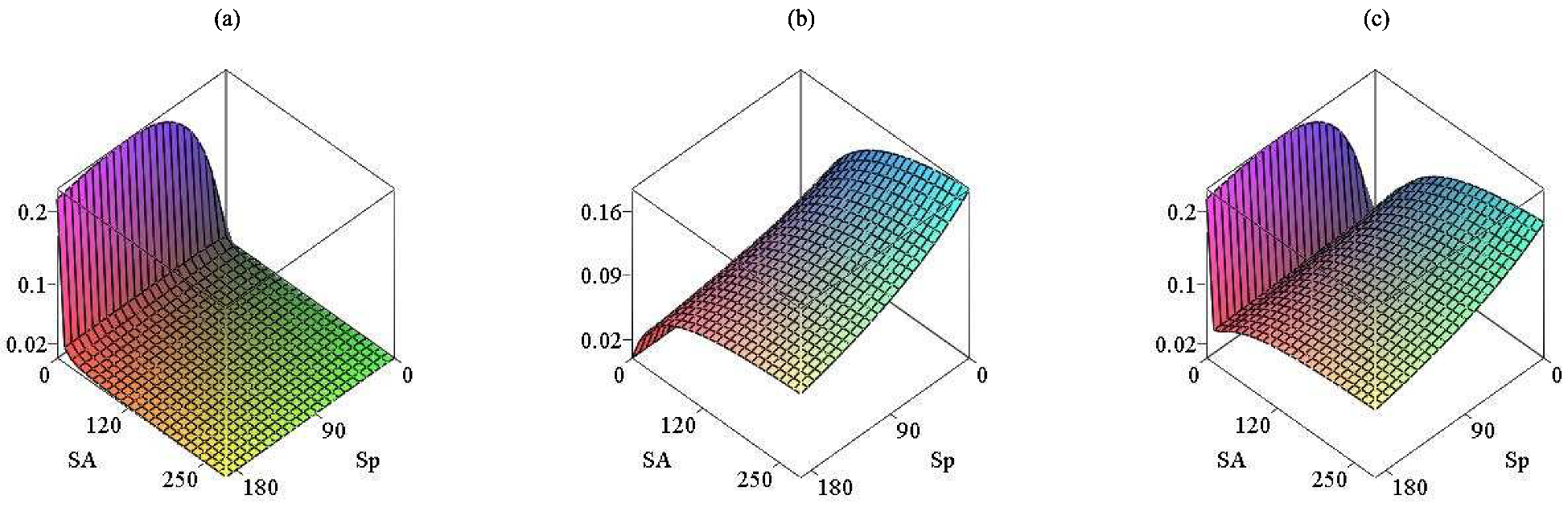
Figure 1 visualizes the functions
,
and
.
The explicit expressions of
and
(see (
4)) suggest the following properties of the latter:
Property 1. For , , with if , ; is continuously differentiable and bounded; Property 2. For , , with if , ; is continuously differentiable and bounded; 4. Global Analysis
In this section we provide the most important properties of the dynamics (
1)–(
3). We establish existence and positivity of the solutions for all time
—properties, that ensure the ability of the mathematical model to describe the bioprocess, regarding its practical applicability. Further, we show the global asymptotic stability of the equilibrium points with respect to the dilution rate
D, which actually means model-based control design of the process. These results provide a good framework for practical applications by indicating to the experimenter how to choose the proper control strategy in order to ensure best process performance and wastewater depollution up to known ecological norms.
Theorem 1. The nonnegative cone and the interior of the nonnegative cone in are positively invariant under the flow (1)–(3). Proof. If
at some time moment
then by Equation (
3) it follows
for all
due to uniqueness of solutions of Cauchy’s problem. Then the model reduces to
which solutions are
Obviously,
and
exponentially as
. So, the face
is invariant under the flow (
1)–(
3).
If
then it follows from Equation (
3)
which means that
for all
.
If
for some
then by Equation (
1),
. If
for some
then Equation (
2) implies
. Therefore the vector field of (
1)–(
3) points inside the positive orthant, i.e., all model solutions are positive. This completes the proof of Theorem 1. □
In what follows we shall consider initial conditions for the dynamics (
1)–(
3) in the set
According to Theorem 1 the set is positively invariant for the model, i.e., starting with initial conditions in the corresponding solutions remain in for all time .
Theorem 2. Let . Then all solutions are uniformly bounded and thus exist for all time .
Proof. After multiplying Equation (
1) by
, Equation (
2) by
and adding the latter to Equation (
3) we obtain
Denoting
, Equation (
12) implies
which yields
. According to Theorem 1 all solutions are positive, and the latter presentation means that all solutions are uniformly bounded and thus exist for all
. The proof of Theorem 2 is completed. □
In the following we shall use the next Lemma.
Barbălat’s Lemma (cf. [
41])
. If is uniformly continuous and there exists then . Theorem 3. Let . The following assertions are valid.
- (i)
For any and for any there exists time such that for all , and hold true.
- (ii)
If , then there exists time such that for all , and are fulfilled.
Proof. Let
be any value of the control function. If
holds for all
then by Equation (
1) we obtain
for all
. If there is a time moment
such that
then
. This means that if there is a time moment
such that
then
for all
is valid. Therefore,
converges to some
as
. If
then
for all
, which means that
as
, a contradiction. Thus, either
for all sufficiently large
or
converges to
as
. Hence, for any
there exists time
such that
for all
holds true.
Similar conclusion can be made for
using the model Equation (
2), i.e., either
for all sufficiently large
or
converges to
as
. Equivalently, for any
there exists time
so that for all
the inequality
holds true. Then choosing
proves point
of the theorem.
Choose and fix some
. The proof of point
implies that
is strictly decreasing with time. Moreover, since the set
is bonded, it follows that there exists
. Similarly,
is strictly decreasing, too, and there exists
. Since
,
and
are bounded differentiable functions for all
it follows that
is uniformly continuous. Applying Barbălat’s Lemma yields
We have by Theorem 1 that
,
,
, and because
, the latter equality (
13) implies
,
as
. In a similar way one obtains that
as
.
Since
, by Equation (
3) we obtain
Further, the relations
,
as
, as well as the properties
,
of
and
imply that there exists a time moment
and a constant
such that
for all
is fulfilled. Then
for all
. The invariance of
with respect to the trajectories of the system implies that
. Then from
for all
it follows that
for each
, a contradiction with
as
. Hence, there exists a sufficiently large time
such that
holds true for all
. If for some time moment
the equality
is fulfilled, then
This shows that
for all sufficiently large
is satisfied.
In a similar way it can be shown that there exists time such that for all holds true. Choosing it follows that and are simultaneously satisfied for all .
The proof of Theorem 3 is completed. □
Below we shall establish the global asymptotic stability of the boundary equilibrium . This property of the washout steady state is also important because it characterizes the inability of the microorganisms to survive in the chemostat system and to degrade the organic chemical compounds.
Theorem 4. For any initial point from Ω
and any the corresponding solution of (1)–(3) converges asymptotically to the boundary equilibrium . Proof. Choose an arbitrary initial point
, and let
be some value of the dilution rate. Suppose that
. By Barbălat’s Lemma we obtain from Equation (
3)
which leads to
Based on Theorem 3
, on the properties
and
of
and
, and since
, the latter relation implies that there exists a time moment
and a constant
such that
for all
. This yields
, or equivalently,
for all
, a contradiction with
. Hence,
holds true. Further, applying the theory of the asymptotically autonomous systems, the model (
1)–(
3) reduces to the limiting system
which means that
,
. This proves the global asymptotic stability of the washout equilibrium
. □
The next considerations concern the global asymptotic stability of the interior equilibrium whenever .
Experimental results in [
16] indicate that SA is degraded more rapidly by
P. putida 49451 cells than phenol. For that reason let us assume that the model dynamics is already stabilized at
for some value
. Denote
Then model (
1)–(
3) can be reduced to the following 2-dimensional system with respect to
and
X:
Further, by (
9) and (
10) we have
, and the above two equations can be rewritten in the form
We shall show that the dynamics (
15) is asymptotically stabilizable towards
.
Theorem 5. For any initial point the corresponding solution of (15) converges asymptotically to . Proof. From Theorem 3
it follows that there is no loss of generality if we restrict our considerations to initial conditions from the set
Define the following Lyapunov function
where
is a positive constant, which will be determined later. Obviously,
V is continuously differentiable in
,
for all
with
, and
at
. It is straightforward to see, that the derivative of
V along the solutions of (
15) is
Since all model solutions are positive and bounded, we can choose the constant
sufficiently large so that
for all
. Obviously,
if and only if
and
are fulfilled. By LaSalle’s invariance principle (cf. [
42]) every solution of (
15) initiating in
approaches the largest invariant set
. Since
is locally asymptotically stable, it follows that
. Therefore,
is globally asymptotically stable for system (
15), and this means that all solutions of (
1)–(
3) converge to
as
. The proof of Theorem 5 is completed. □
Remark 1. Similar conclusions about the global stability of can be made by assuming that the dynamics is first stabilized at for some and then show that the solutions converge asymptotically to as . This will be in agreement with the experimental work in [14] where it is concluded that P. putida CCRC 14365 cells preferably degrade phenol rather than SA. 5. Numerical Simulation of the Model Dynamics
In this section we consider numerical examples demonstrating the dynamic behavior of the model (
1)–(
3) in accordance with the theoretical results.
Example 1. As mentioned before, the model (
1)–(
3) has been tested at this value of
D in [
16]. It is shown there that the solutions fall in finite time into the point
, called a steady state, but it is not. Our computer simulations deliver the following components for the interior equilibrium
, which are quite different from that ones of
F. According to Theorem 5 namely the equilibrium
is globally asymptotically stable and attracts all solutions for any initial point from the set
as time tends to infinity. Practically this means that after finite time the solutions fall into a neighborhood of
, say a ball with center
and radius
, where the value
r (called also tolerance) can be chosen by the user.
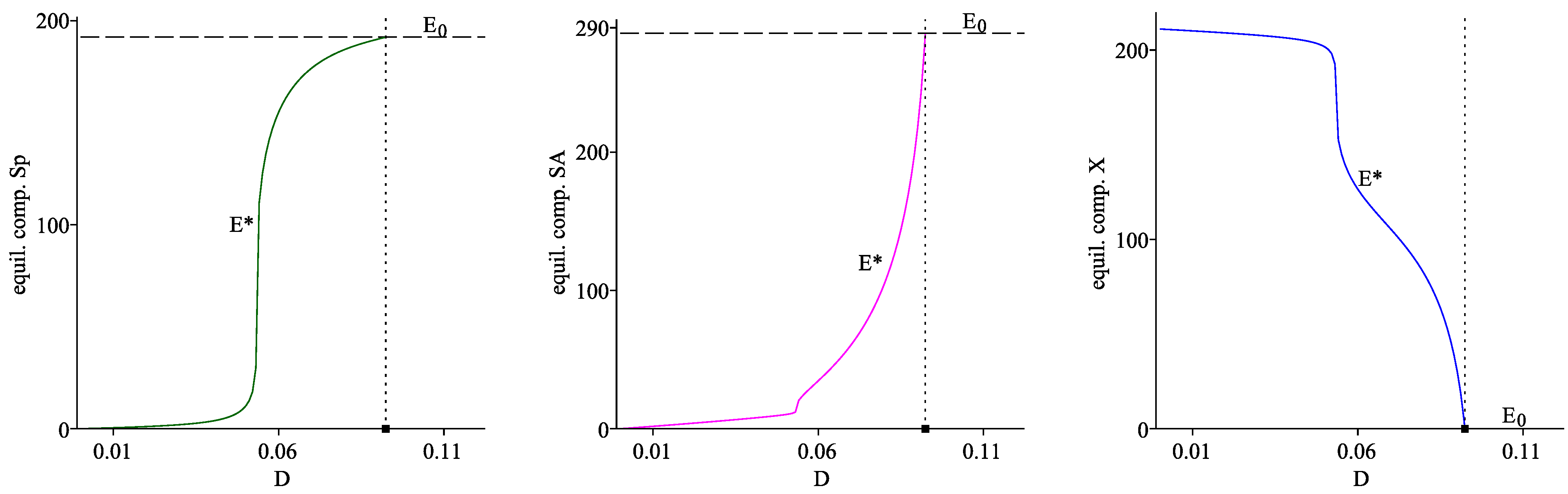
At the equilibrium components of the interior equilibrium are . Obviously, lower values of the dilution rate D lead to lower values of and , but high values of in the global attractor .
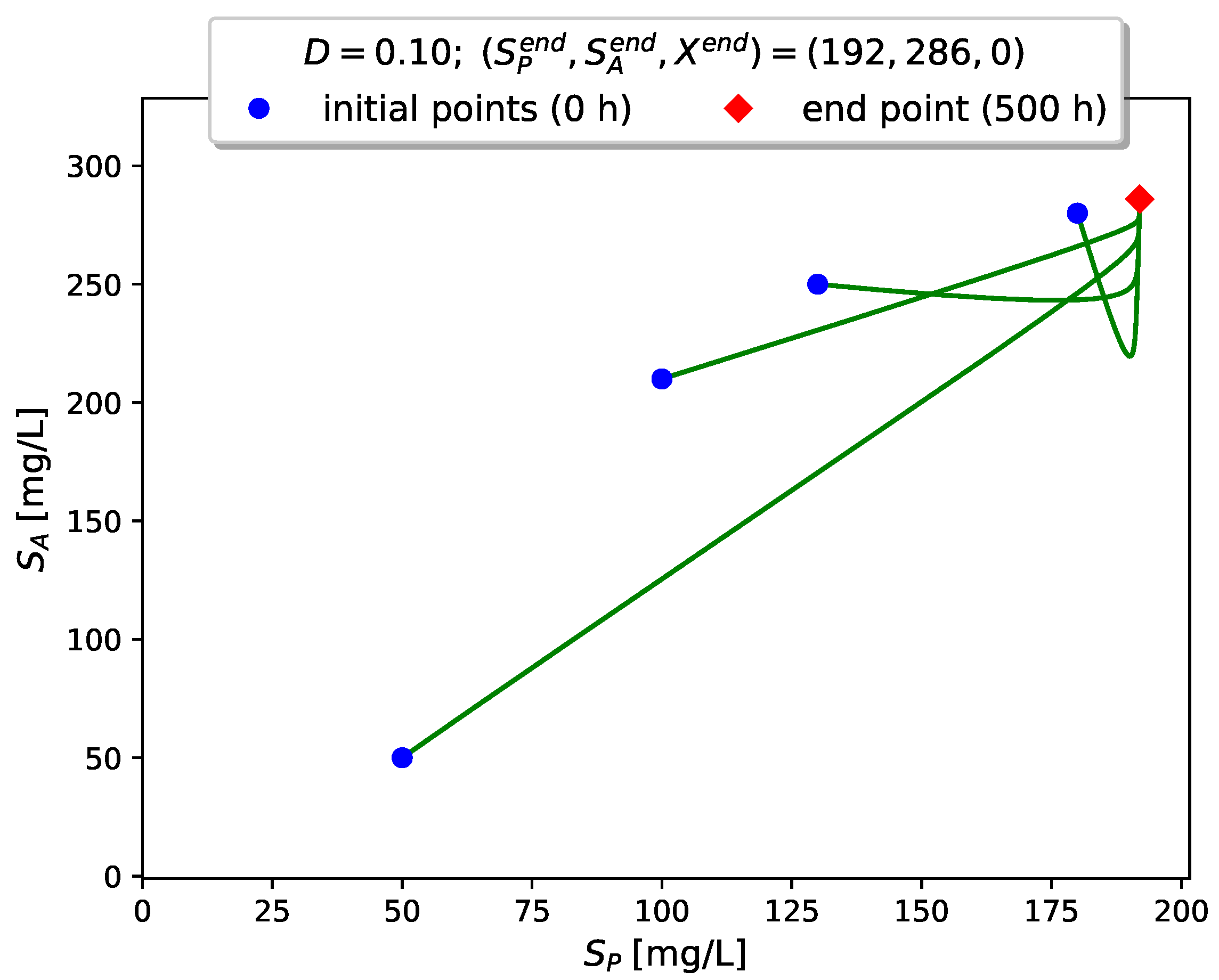
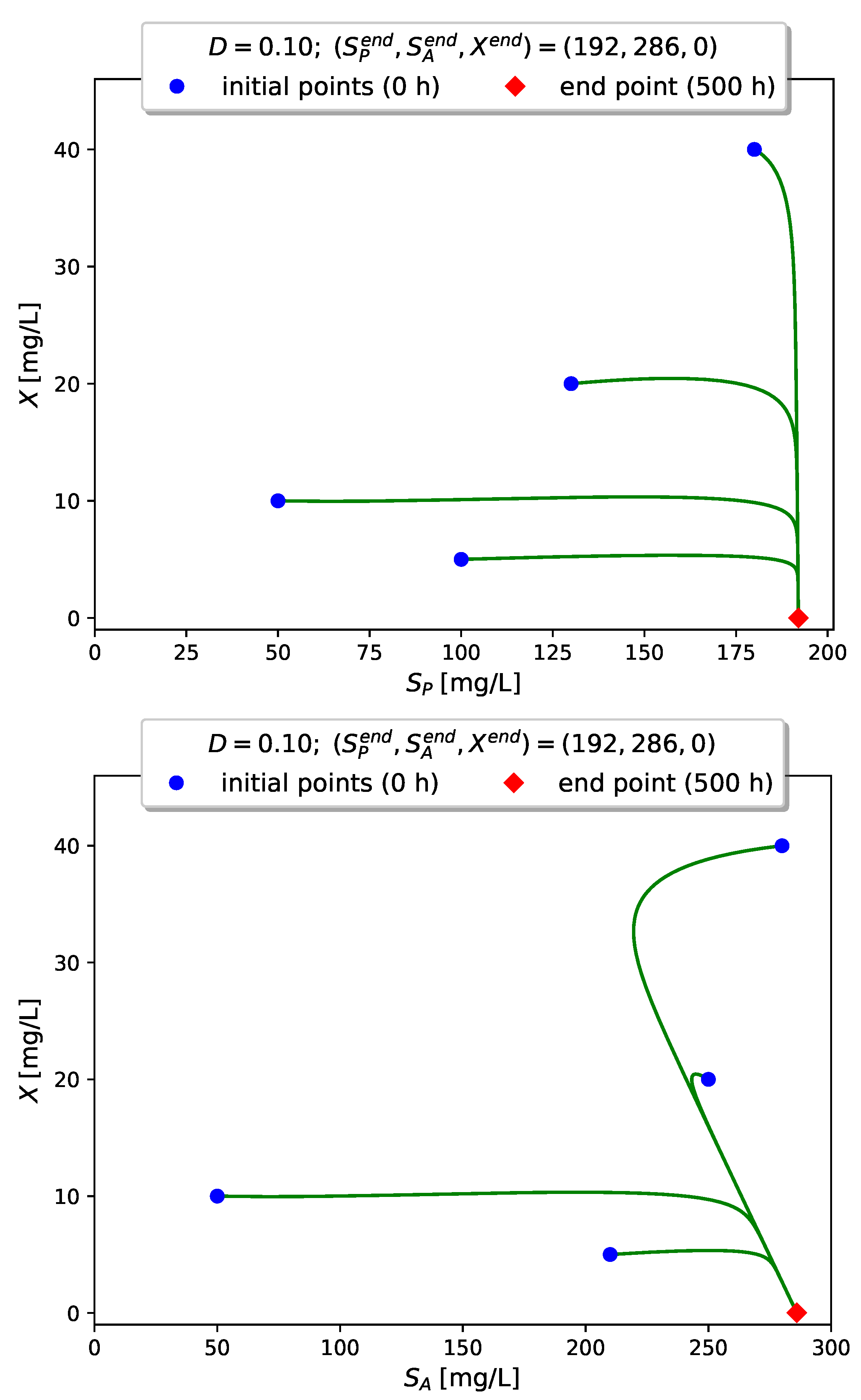
In this case we have , so that the boundary equilibrium is the unique global attractor of the model.
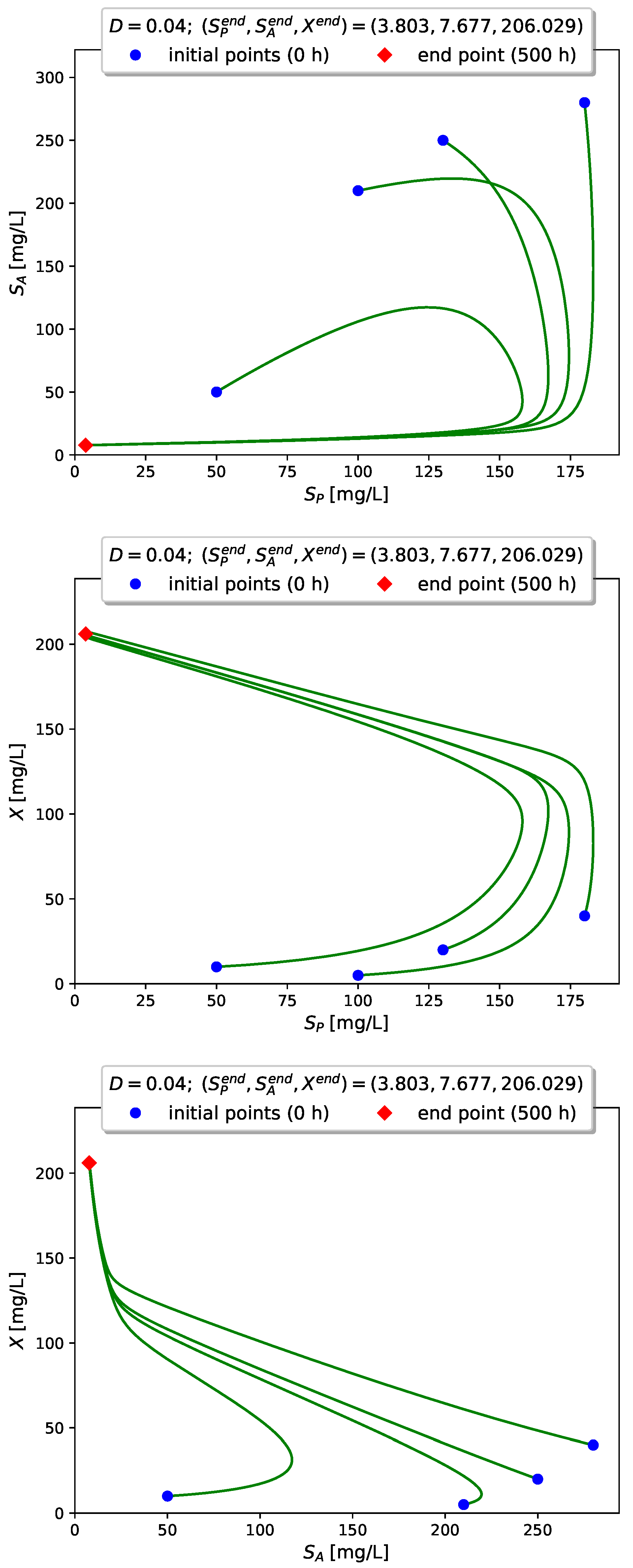
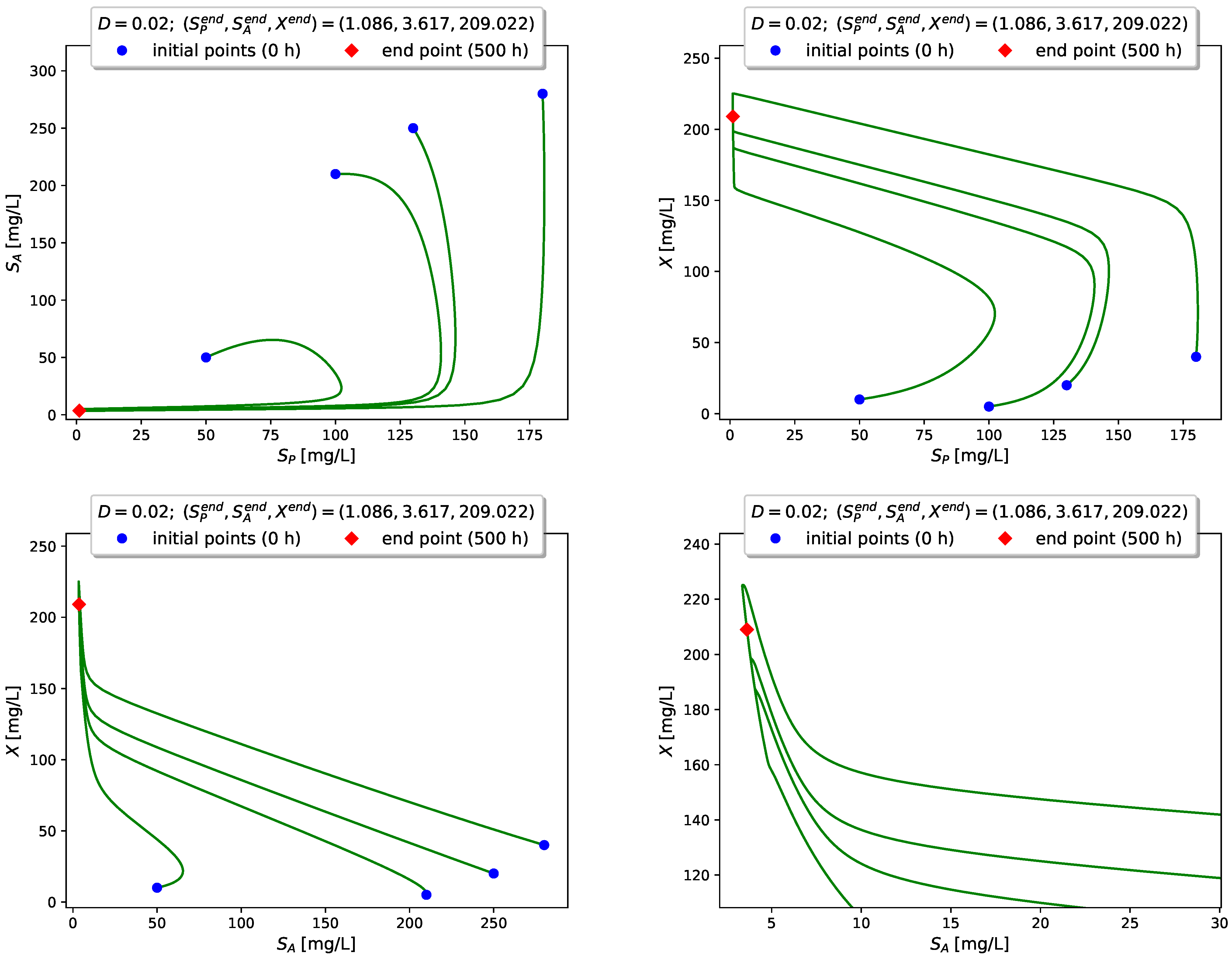
Figure 4 visualizes the time evolution of
,
and
for the 3 different values of
D corresponding to Examples 1–3.
Figure 5,
Figure 6 and
Figure 7 show projections of several trajectories in different phase planes for values of
D according to Examples 1, 2 and 3, respectively.
The computer simulations with model (
1)–(
3) confirm the global stabilizability of the dynamics to either the interior (persistence) equilibrium
if
or to the boundary (washout) equilibrium
when
.
6. Discussion and Conclusions
In this paper we provide a mathematical analysis of the model for biodegradation of phenol and sodium salicylate in a chemostat by
P. putida 49451 cells, proposed for the first time and experimentally validated in [
16]. The model is described by a system of three nonlinear ordinary differential equations involving SKIP kinetics as specific growth rate of the microorganisms. The mathematical investigation of the dynamical system includes local and global analysis of the solutions. Two equilibrium points—one interior (persistence) and one boundary (washout) equilibrium—are computed in dependance of the dilution rate
D as an important model parameter. A critical value
is found, such that the interior equilibrium point
exists if
. The boundary steady state
is available for all values of
. It is shown by numerical computations that
is locally asymptotically stable whenever it exists, and
is locally asymptotically stable for
, and unstable if
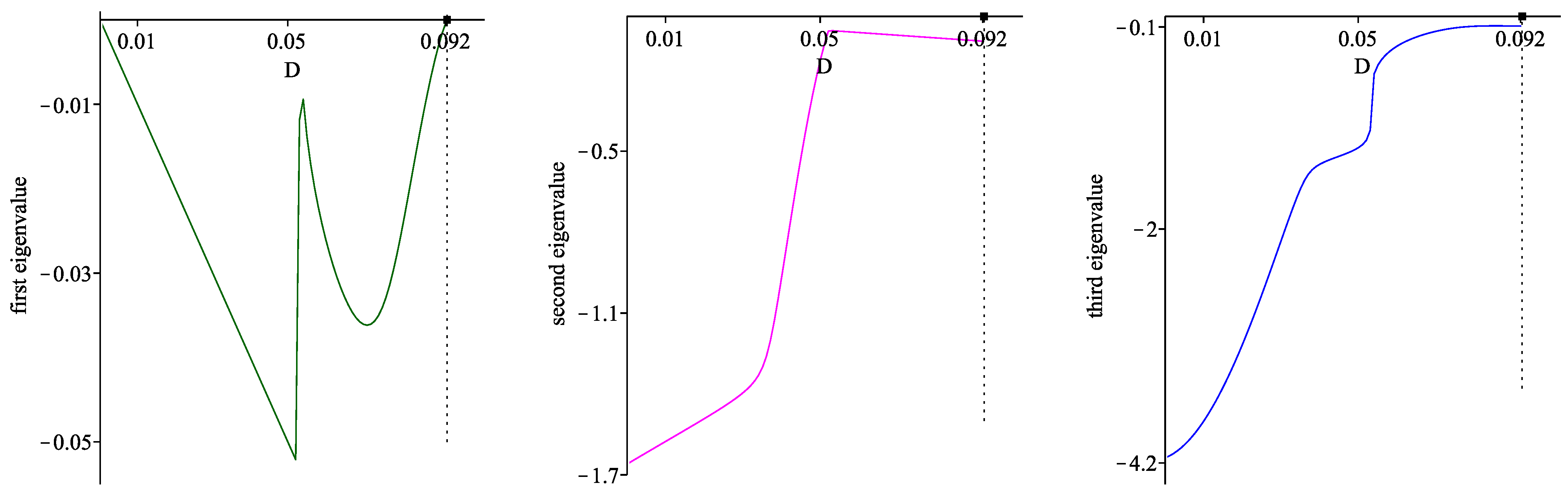
. These conclusions are summarized in Proposition 1.
The most important properties of the model solutions—existence, positivity, uniqueness and uniform boundedness—are established theoretically in
Section 4, by Theorems 1–3. In Theorem 4 we prove the global stability of the boundary equilibrium
(within
) if the values of the dilution rate
D are large, i.e., if
. As usual, the global stability of
is interpreted as total washout of the microorganisms from the chemostat leading to process breakdown. Theorem 5 is devoted to global stability of the interior equilibrium
for any
. The theorem is proved by assuming that the model dynamics is already stabilized to
for some value
, and then it is shown, by providing an explicit Lyapunov function, that the solutions
and
converge asymptotically to
and
respectively as
for any initial point in the set
. A similar result can be obtained supposing that the dynamics is first stabilized to
, and then showing that
and
respectively as
(see Remark 1). The global stability characteristics give useful advises to the experimenter how to tune the dilution rate
D in order to control the biodegradation of the chemical compounds up to prescribed ecological norms.
It remains an open problem to prove the global asymptotic stability of the interior equilibrium
with
for the whole system (
1)–(
3), for example by constructing an appropriate Lyapunov function or using other approaches. This will be a subject of future studies.
Some numerical examples for different values of the dilution rate
D support the theoretical studies and illustrate the dynamic behavior of the solutions. The model predictions are in agreement with the experimental work in [
16] for phenol and SA biodegradation by
P. putida cells.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}