
An Optimized Purification Design for Extracting Active ADAMTS13 from Conditioned Media

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line Generation and Growth
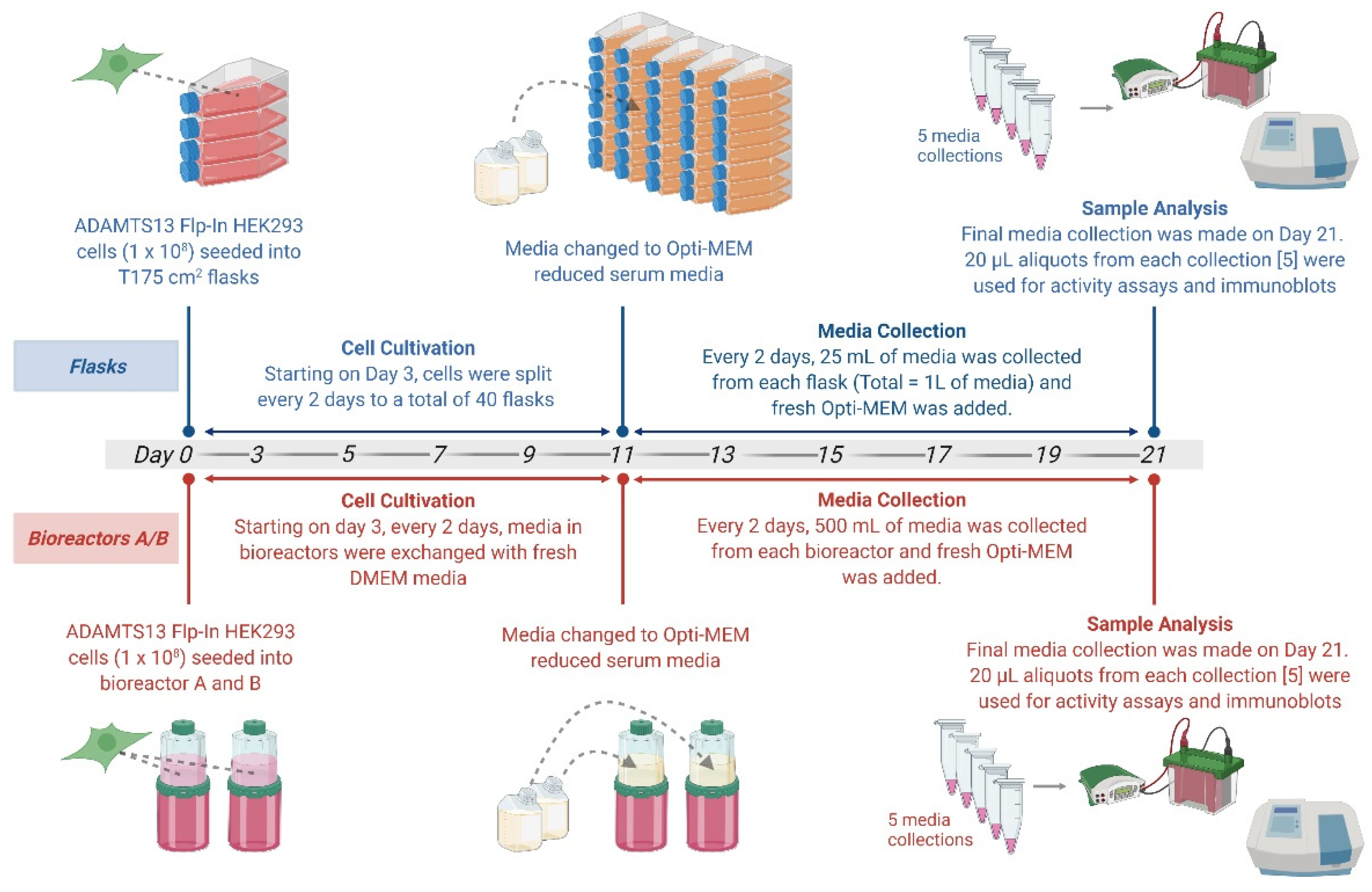
2.2. Cell Growth in BelloCell Culture Bottle System
2.3. Cell Harvest and Media Collection
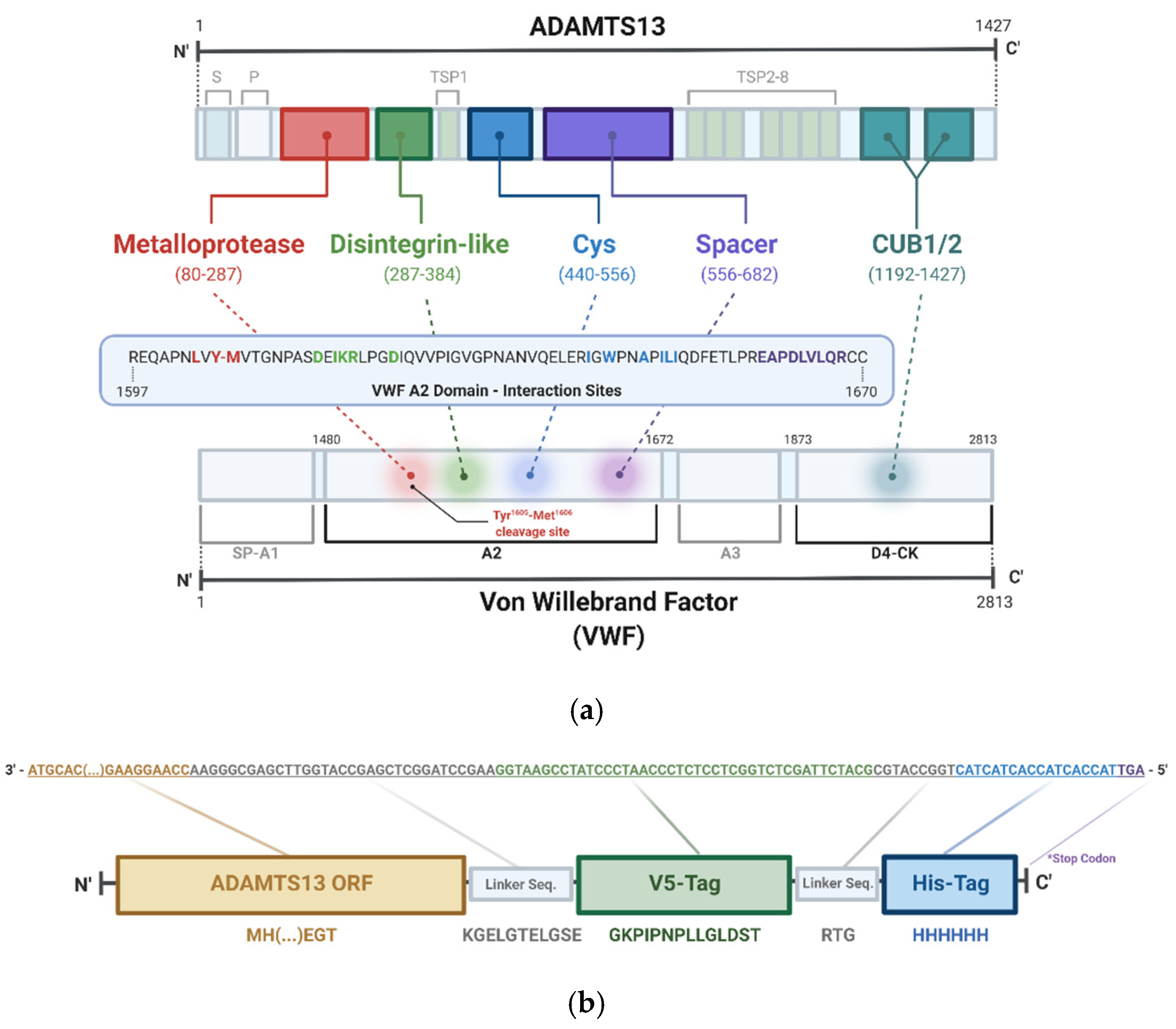
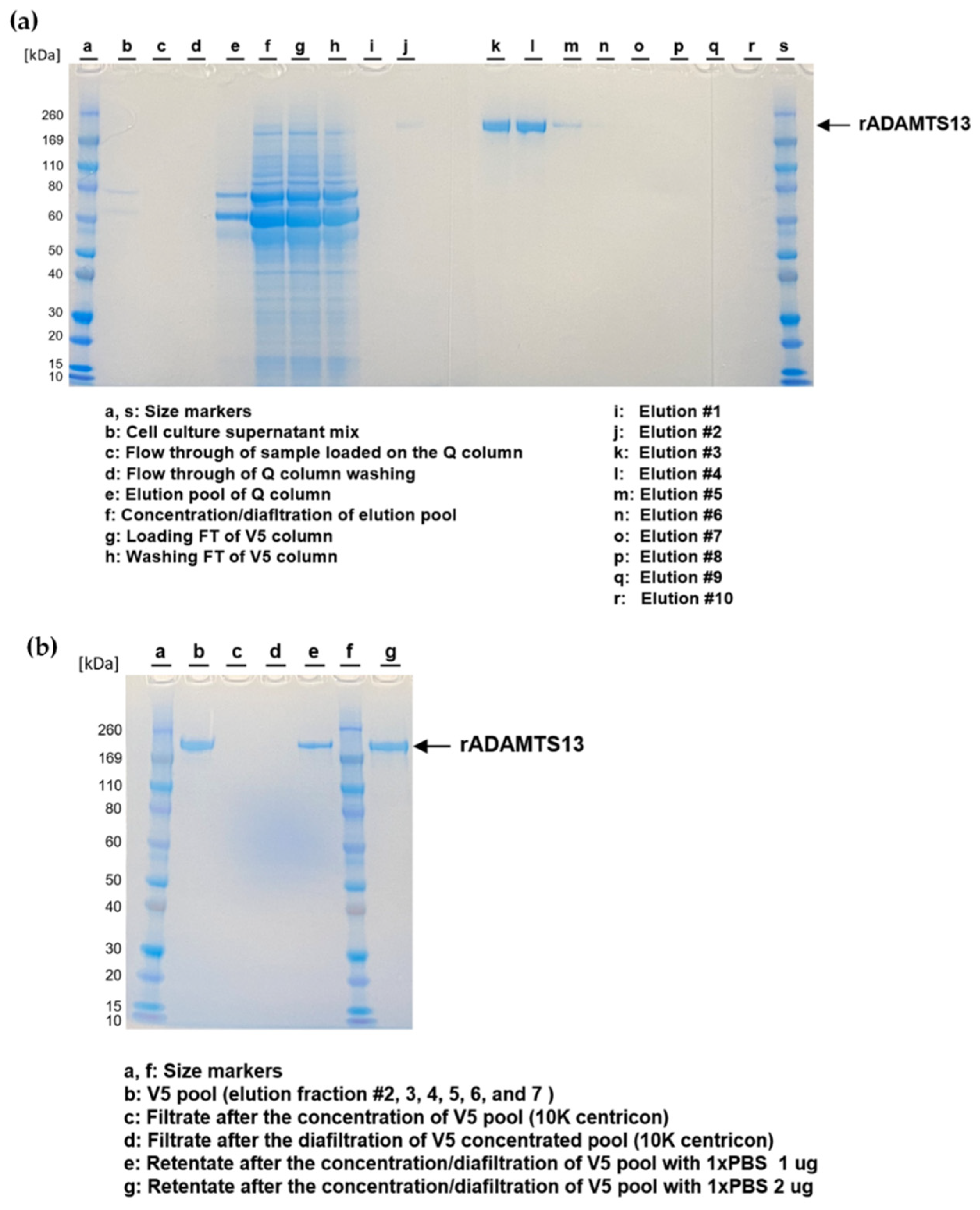
2.4. rADAMTS13 Purification
2.5. rADAMTS13 Assessment
2.5.1. Analysis of rADAMTS13 Using SDS-PAGE and Western Blotting
2.5.2. rADAMTS13 Antigen Measurement
2.5.3. rADAMTS13 Activity Measurement
2.5.4. Glycosylation Analysis
3. Results
3.1. Purification of rADAMTS13 from Conditioned Media of Flp-In HEK293 Cells
3.1.1. Expression of rADAMTS13 in Flp-In HEK293 Cells
3.1.2. Comparison of Flasks and Bioreactors as Optimal Cultivation Strategies
3.2. Purification and Quality Assessment of rADAMTS13 from Conditioned Media
3.3. Post-Translational Modification Analysis of Purified rADAMTS13
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Shang, D.; Zheng, X.W.; Niiya, M.; Zheng, X.L. Apical sorting of ADAMTS13 in vascular endothelial cells and Madin-Darby canine kidney cells depends on the CUB domains and their association with lipid rafts. Blood 2006, 108, 2207–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickens, B.; Mao, Y.; Li, D.; Siegel, D.L.; Poncz, M.; Cines, D.B.; Zheng, X.L. Platelet-delivered ADAMTS13 inhibits arterial thrombosis and prevents thrombotic thrombocytopenic purpura in murine models. Blood J. Am. Soc. Hematol. 2015, 125, 3326–3334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Inada, M.; Lee, T.-P.; Benten, D.; Lyubsky, S.; Bouhassira, E.E.; Gupta, S.; Tsai, H.-M. ADAMTS13 is expressed in hepatic stellate cells. Lab. Investig. 2005, 85, 780–788. [Google Scholar] [CrossRef]
- Zheng, X.L. Structure-function and regulation of ADAMTS-13 protease. J. Thromb. Haemost. 2013, 11 (Suppl. 1), 11–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggeri, Z.M. Von Willebrand factor, platelets and endothelial cell interactions. J. Thromb. Haemost. 2003, 1, 1335–1342. [Google Scholar] [CrossRef]
- Barg, A.; Ossig, R.; Goerge, T.; Schneider, M.F.; Schillers, H.; Oberleithner, H.; Schneider, S.W. Soluble plasma-derived von Willebrand factor assembles to a haemostatically active filamentous network. Thromb. Haemost. 2007, 97, 514–526. [Google Scholar] [CrossRef]
- Furlan, M. Von Willebrand factor: Molecular size and functional activity. Ann. Hematol. 1996, 72, 341–348. [Google Scholar] [CrossRef]
- Mannucci, P.M.; Canciani, M.T.; Forza, I.; Lussana, F.; Lattuada, A.; Rossi, E. Changes in health and disease of the metalloprotease that cleaves von Willebrand factor. Blood 2001, 98, 2730–2735. [Google Scholar] [CrossRef]
- Ma, Q.; Jacobi, P.M.; Emmer, B.T.; Kretz, C.A.; Ozel, A.B.; McGee, B.; Kimchi-Sarfaty, C.; Ginsburg, D.; Li, J.Z.; Desch, K.C. Genetic variants in ADAMTS13 as well as smoking are major determinants of plasma ADAMTS13 levels. Blood Adv. 2017, 1, 1037–1046. [Google Scholar] [CrossRef] [Green Version]
- Mannucci, P.M.; Capoferri, C.; Canciani, M.T. Plasma levels of von Willebrand factor regulate ADAMTS-13, its major cleaving protease. Br. J. Haematol. 2004, 126, 213–218. [Google Scholar] [CrossRef]
- Page, E.E.; Kremer Hovinga, J.A.; Terrell, D.R.; Vesely, S.K.; George, J.N. Thrombotic thrombocytopenic purpura: Diagnostic criteria, clinical features, and long-term outcomes from 1995 through 2015. Blood Adv. 2017, 1, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.L. ADAMTS13 and von Willebrand factor in thrombotic thrombocytopenic purpura. Annu. Rev. Med. 2015, 66, 211–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, M.; McDaniel, J.K.; Zheng, X.L. Thrombotic thrombocytopenic purpura: Pathogenesis, diagnosis and potential novel therapeutics. J. Thromb. Haemost. 2017, 15, 1889–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambers, M.; Goldenberg, N.A.; Kenet, G.; Kirkham, F.J.; Manner, D.; Bernard, T.; Mesters, R.M.; Junker, R.; Stoll, M.; Nowak-Göttl, U. Role of reduced ADAMTS13 in arterial ischemic stroke: A Pediatric Cohort Study. Ann. Neurol. 2013, 73, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Cao, Y.; Yang, X.; Cai, P.; Kang, L.; Zhu, X.; Luo, H.; Lu, L.; Wei, L.; Bai, X.; et al. ADAMTS13 controls vascular remodeling by modifying VWF reactivity during stroke recovery. Blood 2017, 130, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Eerenberg, E.S.; Teunissen, P.F.; van den Born, B.J.; Meijers, J.C.; Hollander, M.R.; Jansen, M.; Tijssen, R.; Beliën, J.A.; van de Ven, P.M.; Aly, M.F.; et al. The role of ADAMTS13 in acute myocardial infarction: Cause or consequence? Cardiovasc. Res. 2016, 111, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Chion, C.K.N.K.; Doggen, C.J.M.; Crawley, J.T.B.; Lane, D.A.; Rosendaal, F.R. ADAMTS13 and von Willebrand factor and the risk of myocardial infarction in men. Blood 2006, 109, 1998–2000. [Google Scholar] [CrossRef] [Green Version]
- Katneni, U.K.; Ibla, J.C.; Hunt, R.; Schiller, T.; Kimchi-Sarfaty, C. von Willebrand factor/ADAMTS-13 interactions at birth: Implications for thrombosis in the neonatal period. J. Thromb. Haemost. 2019, 17, 429–440. [Google Scholar] [CrossRef]
- Mancini, I.; Baronciani, L.; Artoni, A.; Colpani, P.; Biganzoli, M.; Cozzi, G.; Novembrino, C.; Boscolo Anzoletti, M.; De Zan, V.; Pagliari, M.T.; et al. The ADAMTS13-von Willebrand factor axis in COVID-19 patients. J. Thromb. Haemost. 2021, 19, 513–521. [Google Scholar] [CrossRef]
- Tang, N.; Li, D.; Wang, X.; Sun, Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J. Thromb. Haemost. 2020, 18, 844–847. [Google Scholar] [CrossRef] [Green Version]
- Hindilerden, F.; Yonal-Hindilerden, I.; Akar, E.; Kart-Yasar, K. COVID-19 associated autoimmune thrombotic thrombocytopenic purpura: Report of a case. Thromb. Res. 2020, 195, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Scully, M.; Knöbl, P.; Kentouche, K.; Rice, L.; Windyga, J.; Schneppenheim, R.; Hovinga, J.A.K.; Kajiwara, M.; Fujimura, Y.; Maggiore, C.; et al. Recombinant ADAMTS-13: First-in-human pharmacokinetics and safety in congenital thrombotic thrombocytopenic purpura. Blood 2017, 130, 2055–2063. [Google Scholar] [CrossRef] [PubMed]
- Turecek, P.L.; Peck, R.C.; Rangarajan, S.; Reilly-Stitt, C.; Laffan, M.A.; Kazmi, R.; James, I.; Dushianthan, A.; Schrenk, G.; Gritsch, H.; et al. Recombinant ADAMTS13 reduces abnormally up-regulated von Willebrand factor in plasma from patients with severe COVID-19. Thromb. Res. 2021, 201, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Franchini, M. Plasma-derived versus recombinant Factor VIII concentrates for the treatment of haemophilia A: Recombinant is better. Blood Transfus. 2010, 8, 292–296. [Google Scholar] [CrossRef]
- Petri, A.; Kim, H.J.; Xu, Y.; de Groot, R.; Li, C.; Vandenbulcke, A.; Vanhoorelbeke, K.; Emsley, J.; Crawley, J.T.B. Crystal structure and substrate-induced activation of ADAMTS13. Nat. Commun. 2019, 10, 3781. [Google Scholar] [CrossRef] [Green Version]
- Ercig, B.; Wichapong, K.; Reutelingsperger, C.P.M.; Vanhoorelbeke, K.; Voorberg, J.; Nicolaes, G.A.F. Insights into 3D Structure of ADAMTS13: A Stepping Stone towards Novel Therapeutic Treatment of Thrombotic Thrombocytopenic Purpura. Thromb. Haemost. 2018, 118, 28–41. [Google Scholar] [CrossRef]
- Kim, H.J.; Xu, Y.; Petri, A.; Vanhoorelbeke, K.; Crawley, J.T.B.; Emsley, J. Crystal structure of ADAMTS13 CUB domains reveals their role in global latency. Sci. Adv. 2021, 7, eabg4403. [Google Scholar] [CrossRef]
- South, K.; Luken, B.M.; Crawley, J.T.; Phillips, R.; Thomas, M.; Collins, R.F.; Deforche, L.; Vanhoorelbeke, K.; Lane, D.A. Conformational activation of ADAMTS13. Proc. Natl. Acad. Sci. USA 2014, 111, 18578–18583. [Google Scholar] [CrossRef] [Green Version]
- Zanardelli, S.; Crawley, J.T.; Chion, C.K.; Lam, J.K.; Preston, R.J.; Lane, D.A. ADAMTS13 substrate recognition of von Willebrand factor A2 domain. J. Biol. Chem. 2006, 281, 1555–1563. [Google Scholar] [CrossRef] [Green Version]
- Feys, H.B.; Pareyn, I.; Vancraenenbroeck, R.; De Maeyer, M.; Deckmyn, H.; Van Geet, C.; Vanhoorelbeke, K. Mutation of the H-bond acceptor S119 in the ADAMTS13 metalloprotease domain reduces secretion and substrate turnover in a patient with congenital thrombotic thrombocytopenic purpura. Blood 2009, 114, 4749–4752. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Pan, W.; Rux, A.H.; Sachais, B.S.; Zheng, X.L. The cooperative activity between the carboxyl-terminal TSP1 repeats and the CUB domains of ADAMTS13 is crucial for recognition of von Willebrand factor under flow. Blood 2007, 110, 1887–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, J.; Smith, P.; Wang, S.; Zhang, P.; Zheng, X.L. The proximal carboxyl-terminal domains of ADAMTS13 determine substrate specificity and are all required for cleavage of von Willebrand factor. J. Biol. Chem. 2005, 280, 29428–29434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soejima, K.; Matsumoto, M.; Kokame, K.; Yagi, H.; Ishizashi, H.; Maeda, H.; Nozaki, C.; Miyata, T.; Fujimura, Y.; Nakagaki, T. ADAMTS-13 cysteine-rich/spacer domains are functionally essential for von Willebrand factor cleavage. Blood 2003, 102, 3232–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, S.; Clark, I.M.; Kevorkian, L.; Edwards, D.R. The ADAMTS metalloproteinases. Biochem. J. 2005, 386, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Cerdà-Costa, N.; Xavier Gomis-Rüth, F. Architecture and function of metallopeptidase catalytic domains. Protein Sci. 2014, 23, 123–144. [Google Scholar] [CrossRef] [Green Version]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Hershko, K.; Simhadri, V.L.; Blaisdell, A.; Hunt, R.C.; Newell, J.; Tseng, S.C.; Hershko, A.Y.; Choi, J.W.; Sauna, Z.E.; Wu, A.; et al. Cyclosporin A impairs the secretion and activity of ADAMTS13 (a disintegrin and metalloprotease with thrombospondin type 1 repeat). J. Biol. Chem. 2012, 287, 44361–44371. [Google Scholar] [CrossRef] [Green Version]
- Ricketts, L.M.; Dlugosz, M.; Luther, K.B.; Haltiwanger, R.S.; Majerus, E.M. O-fucosylation is required for ADAMTS13 secretion. J. Biol. Chem. 2007, 282, 17014–17023. [Google Scholar] [CrossRef] [Green Version]
- Sorvillo, N.; Kaijen, P.H.; Matsumoto, M.; Fujimura, Y.; van der Zwaan, C.; Verbij, F.C.; Pos, W.; Fijnheer, R.; Voorberg, J.; Meijer, A.B. Identification of N-linked glycosylation and putative O-fucosylation, C-mannosylation sites in plasma derived ADAMTS13. J. Thromb. Haemost. 2014, 12, 670–679. [Google Scholar] [CrossRef] [Green Version]
- Verbij, F.C.; Stokhuijzen, E.; Kaijen, P.H.P.; van Alphen, F.; Meijer, A.B.; Voorberg, J. Identification of glycans on plasma-derived ADAMTS13. Blood 2016, 128, e51–e58. [Google Scholar] [CrossRef]
- Colige, A.C. Challenges and Solutions for Purification of ADAMTS Proteases: An Overview. Methods Mol. Biol. 2020, 2043, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Nishio, K.; Majerus, E.M.; Sadler, J.E. Cleavage of von Willebrand Factor Requires the Spacer Domain of the Metalloprotease ADAMTS13 *. J. Biol. Chem. 2003, 278, 30136–30141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawley, J.T.B.; Lam, J.K.; Rance, J.B.; Mollica, L.R.; O’Donnell, J.S.; Lane, D.A. Proteolytic inactivation of ADAMTS13 by thrombin and plasmin. Blood 2005, 105, 1085–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokame, K.; Nobe, Y.; Kokubo, Y.; Okayama, A.; Miyata, T. FRETS-VWF73, a first fluorogenic substrate for ADAMTS13 assay. Br. J. Haematol. 2005, 129, 93–100. [Google Scholar] [CrossRef]
- Mackie, I.; Mancini, I.; Muia, J.; Kremer Hovinga, J.; Nair, S.; Machin, S.; Baker, R. International Council for Standardization in Haematology (ICSH) recommendations for laboratory measurement of ADAMTS13. Int. J. Lab. Hematol. 2020, 42, 685–696. [Google Scholar] [CrossRef]
- George, J.N. Measuring ADAMTS13 activity in patients with suspected thrombotic thrombocytopenic purpura: When, how, and why? Transfusion 2015, 55, 11–13. [Google Scholar] [CrossRef] [Green Version]
- Di Stasio, E.; Lancellotti, S.; Peyvandi, F.; Palla, R.; Mannucci, P.M.; De Cristofaro, R. Mechanistic Studies on ADAMTS13 Catalysis. Biophys. J. 2008, 95, 2450–2461. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Liu, W.; Fang, J.; Shi, X.; Wu, J.; Fang, Y.; Lin, J. AFM Imaging Reveals Multiple Conformational States of ADAMTS13. J. Biol. Eng. 2019, 13, 9. [Google Scholar] [CrossRef] [Green Version]
- Hunt, R.; Hettiarachchi, G.; Katneni, U.; Hernandez, N.; Holcomb, D.; Kames, J.; Alnifaidy, R.; Lin, B.; Hamasaki-Katagiri, N.; Wesley, A.; et al. A Single Synonymous Variant (c.354G>A [p.P118P]) in ADAMTS13 Confers Enhanced Specific Activity. Int. J. Mol. Sci. 2019, 20, 5734. [Google Scholar] [CrossRef] [Green Version]
- Cesco BelloCell. BelloCell-500 Technical Report VIII. Available online: https://chemglass.com/download/getmanual/1389 (accessed on 24 December 2021).
- Soejima, K.; Nakamura, H.; Hirashima, M.; Morikawa, W.; Nozaki, C.; Nakagaki, T. Analysis on the Molecular Species and Concentration of Circulating ADAMTS13 in Blood. J. Biochem. 2006, 139, 147–154. [Google Scholar] [CrossRef]
- Zou, G.; Kosikova, M.; Kim, S.R.; Kotian, S.; Wu, W.W.; Shen, R.; Powers, D.N.; Agarabi, C.; Xie, H.; Ju, T. Comprehensive analysis of N-glycans in IgG purified from ferrets with or without influenza A virus infection. J. Biol. Chem. 2018, 293, 19277–19289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Wu, W.W.; Shen, R.; Sjogren, J.; Parsons, L.; Cipollo, J.F. Optimization of O-GIG for O-Glycopeptide Characterization with Sialic Acid Linkage Determination. Anal. Chem. 2020, 92, 10946–10951. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, R.J. Post-translational modifications required for coagulation factor secretion and function. Thromb. Haemost. 1998, 79, 1068–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlaftis, V.; Perera, S.; Monagle, P.; Ignjatovic, V. Importance of post-translational modifications on the function of key haemostatic proteins. Blood Coagul Fibrinolysis 2016, 27, 1–4. [Google Scholar] [CrossRef]
- Wang, L.W.; Leonhard-Melief, C.; Haltiwanger, R.S.; Apte, S.S. Post-translational Modification of Thrombospondin Type-1 Repeats in ADAMTS-like 1/Punctin-1 by C-Mannosylation of Tryptophan*. J. Biol. Chem. 2009, 284, 30004–30015. [Google Scholar] [CrossRef] [Green Version]
- Böhm, E.; Seyfried, B.K.; Dockal, M.; Graninger, M.; Hasslacher, M.; Neurath, M.; Konetschny, C.; Matthiessen, P.; Mitterer, A.; Scheiflinger, F. Differences in N-glycosylation of recombinant human coagulation factor VII derived from BHK, CHO, and HEK293 cells. BMC Biotechnol. 2015, 15, 87. [Google Scholar] [CrossRef] [Green Version]
- Lind, T.; Birch, M.A.; McKie, N. Purification of an insect derived recombinant human ADAMTS-1 reveals novel gelatin (type I collagen) degrading activities. Mol. Cell Biochem. 2006, 281, 95–102. [Google Scholar] [CrossRef]
- Balakirev, M.Y.; Mullally, J.E.; Favier, A.; Assard, N.; Sulpice, E.; Lindsey, D.F.; Rulina, A.V.; Gidrol, X.; Wilkinson, K.D. Wss1 metalloprotease partners with Cdc48/Doa1 in processing genotoxic SUMO conjugates. eLife 2015, 4, e06763. [Google Scholar] [CrossRef]
- Imai, K.; Okada, Y. Purification of matrix metalloproteinases by column chromatography. Nat. Protoc. 2008, 3, 1111–1124. [Google Scholar] [CrossRef]
- Ho, L.; Greene, C.L.; Schmidt, A.W.; Huang, L.H. Cultivation of HEK 293 cell line and production of a member of the superfamily of G-protein coupled receptors for drug discovery applications using a highly efficient novel bioreactor. Cytotechnology 2004, 45, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Waugh, D.S. An overview of enzymatic reagents for the removal of affinity tags. Protein Expr. Purif. 2011, 80, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Alexaki, A.; Hettiarachchi, G.K.; Athey, J.C.; Katneni, U.K.; Simhadri, V.; Hamasaki-Katagiri, N.; Nanavaty, P.; Lin, B.; Takeda, K.; Freedberg, D. Effects of codon optimization on coagulation factor IX translation and structure: Implications for protein and gene therapies. Sci. Rep. 2019, 9, 15449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauro, V.P.; Chappell, S.A. A critical analysis of codon optimization in human therapeutics. Trends Mol. Med. 2014, 20, 604–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayapal, K.P.; Wlaschin, K.F.; Hu, W.; Yap, M.G. Recombinant protein therapeutics from CHO cells-20 years and counting. Chem. Eng. Prog. 2007, 103, 40. [Google Scholar]
- Kim, J.Y.; Kim, Y.-G.; Lee, G.M. CHO cells in biotechnology for production of recombinant proteins: Current state and further potential. Appl. Microbiol. Biotechnol. 2012, 93, 917–930. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Method | Flasks (mg/L) | Culture Bottles (mg/L) |
|---|---|---|
| His-tag | 0.1 1 | - |
| V5 | 0.097 | 0.113 |
| QFF + V5 | 0.678 | 0.145 |
| Purification Method | Protein Yield (mg/L of Media) | Activity (IU/mL) | Specific Activity (IU/ng) |
|---|---|---|---|
| V5 (small-scale) | 0.097 | 1.366 | 0.0593 |
| QFF + V5 (large-scale) | 0.678 | 1.244 | 0.0633 |
| Position | Most Abundant Forms | Glycopeptide Assignment of Tryptic Peptides * | Identified Charge States |
|---|---|---|---|
| N552 | HexNAc(4)Hex(5)Fuc(1) HexNAc(5)Hex(4)Fuc(1) | CQVCGGDNSTCSPR | +3 |
| N579 | HexNAc(5)Hex(3)Fuc(1) HexNAc(5)Hex(4)Fuc(1) | EYVTFLTVTPNLTSVYIANHRPLFTHLAVR | +4 to +6 |
| N614 | HexNAc(2)Hex(6) | MSISPNTTYPSLLEDGR | +2 and +3 |
| N667 | HexNAc(5)Hex(5)Fuc(1) HexNAc(5)Hex(4)Fuc(1) | YGEEYGNLTRPDITFTYFQPKPR | +4 |
| N707 | HexNAc(5)Hex(4)Fuc(1)NeuAc(1) | WVNYSCLDQAR | +3 |
| N1235 | HexNAc(6)Hex(3)Fuc(1)NeuAc(1) | VLESSLNCSAGDMLLLWGR | +3 |
| N1354 | HexNAc(6)Hex(3)Fuc(1)NeuAc(1) | IAIHALATNMGAGTEGANASYILIR | +4 |
| Position | Modification Types | Glycopeptide Assignment of Tryptic Peptides * | Identified Mass † |
|---|---|---|---|
| S399 | Fuc(1)Hex(1) | SCGGGVVTR | 600.7743 (+2) |
| S698 | Fuc(1)Hex(1) | GPCSVSCGAGLR | 764.834 (+2) |
| S757 | Fuc(1)Hex(1) | ELVETVQCQGSQQPPAWPEACVLEPCPPYWAVGDFGPCSASCGGGLR | 1378.8608 (+4) |
| S907 | Fuc(1)Hex(1) | TGAQAAHVWTPAAGSCSVSCGR | 847.0445 (+3) |
| S965 | Fuc(1)Hex(1) | LAACSVSCGR | 694.8052 (+2) |
| S1027 | Fuc(1)Hex(1) | VMSLGPCSASCGLGTAR | 1016.4552 (+2) |
| S1087 | Fuc(1)Hex(1) | WHVGTWMECSVSCGDGIQR | 858.3629 (+3) |
| W387 | Hex | WSSWGPR | 519.2380 (+2) |
| S1170 | HexNAc(1)Hex(1)NeuAc(2) HexNAc(2)Hex(2)NeuAc(2) | GLLFSPAPQPR | 1065.4966 (+2)832.3776 (+3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankowska, K.I.; Katneni, U.; Lin, B.C.; Amarasinghe, R.; Phue, J.-N.; Wu, W.W.; Hamasaki-Katagiri, N.; Jankowski, W.; Shen, R.-F.; Kimchi-Sarfaty, C. An Optimized Purification Design for Extracting Active ADAMTS13 from Conditioned Media. Processes 2022, 10, 322. https://doi.org/10.3390/pr10020322
Jankowska KI, Katneni U, Lin BC, Amarasinghe R, Phue J-N, Wu WW, Hamasaki-Katagiri N, Jankowski W, Shen R-F, Kimchi-Sarfaty C. An Optimized Purification Design for Extracting Active ADAMTS13 from Conditioned Media. Processes. 2022; 10(2):322. https://doi.org/10.3390/pr10020322
Chicago/Turabian StyleJankowska, Katarzyna I., Upendra Katneni, Brian C. Lin, Randilu Amarasinghe, Je-Nie Phue, Wells W. Wu, Nobuko Hamasaki-Katagiri, Wojciech Jankowski, Rong-Fong Shen, and Chava Kimchi-Sarfaty. 2022. "An Optimized Purification Design for Extracting Active ADAMTS13 from Conditioned Media" Processes 10, no. 2: 322. https://doi.org/10.3390/pr10020322