
Systemic Efficacy of Sirolimus via the ERBB Signaling Pathway in Breast Cancer

College of Engineering, Sangmyung University, Cheonan 31066, Korea
Processes 2022, 10(3), 552; https://doi.org/10.3390/pr10030552
Submission received: 7 February 2022
/
Revised: 23 February 2022
/
Accepted: 7 March 2022
/
Published: 11 March 2022
{kind=link}
{kind=link}
Abstract
:Rapamycin, also known as sirolimus, inhibits the mTOR pathway in complex diseases such as cancer, and its downstream targets are ribosomal S6 kinases (RPS6K). Sirolimus is involved in regulating cell growth and cell survival through roles such as the mediation of epidermal growth factor signaling. However, the systemic efficacy of sirolimus in pathway regulation is unclear. The purpose of this study is to determine systemic drug efficacy using computational methods and drug-induced datasets. We suggest a computational method using gene expression datasets induced by sirolimus and an inverse algorithm that simultaneously identifies parameters referring to gene–gene interactions. We downloaded two sirolimus-induced microarray gene expression datasets and used a computational method to obtain the most enriched pathway, then adopted an inverse algorithm to discover the gene–gene interactions of that pathway. In the results, RPS6KB1 was a target gene of sirolimus and was associated with genes in the pathway. The common gene interactions from two datasets were a hub gene, RPS6KB1, and 10 related genes (AKT3, CBLC, MAP2K7, NRG1/2, PAK3, PIK3CD/G, PRKCG, and SHC3) in the epidermal growth factor (ERBB) signaling pathway.
Keywords:
ERBB signaling pathway; sirolimus (rapamycin); systemic drug influence; AKT3; NRG1; NRG2; RPS6KB1; PAK31. Introduction
Rapamycin, which possesses immunosuppressive and anti-proliferative properties, was discovered in 1964 as an antifungal metabolite produced by Streptomyces hygroscopicus [1]. Rapamycin, also known as sirolimus, has been reported to have many effects on cancer, diabetes, obesity, and genetic disorders [1,2]. Sirolimus was discovered to be an inhibitor of S6K1 activation [1,2,3,4,5] and identified as a mammalian target of rapamycin (mTOR) in animal cells and yeast [6,7]. It also downregulates cancer metabolism by inhibiting the oncogenic activation of the mTOR signaling pathway. In the mTOR signaling pathway, some genes, including phosphatidylinositol3-kinase (PI3K), protein kinase B (AKT), and phosphatase and tensin homologue (PTEN), play role as triggers for the activation of cell survival, proliferation and cell growth [8]. Therefore, its therapeutic potential against certain cancers, including gastric cancer, non-small cell lung cancer, and breast cancer [8,9,10], has been evaluated.
Since the AKT/mTOR and epidermal growth factor receptor (ERBB) signaling pathways are growth factor receptor cascades in cancer, many studies have demonstrated their expected crosstalk mechanisms [2,11,12]. For example, the drug gefitinib was reported as an inhibitor of the crosstalk between the mTOR and ERBB signaling pathways in breast cancer [13]. Sirolimus also induced the transactivation of the EGFR signaling pathway [14]. Since some co-treatments restored the resistance of drugs, inferring systemic gene interaction in an enriched pathway by an objective drug is a novel therapeutic method. The information of the pathway as gene sets was curated and a peer-reviewed knowledge base of biomolecular pathways is provided from several open sources, such as the Kyoto encyclopedia of genes (KEGG) [15], Biocarta [16], and reactome [17,18]. In this study, we perform all pathway analyses based on KEGG. The mTOR signaling pathway consists of 48 genes, and the ERBB signaling pathway consists of 87 genes introduced from KEGG [15]. There are 18 overlapping genes between the two pathways (AKT1/2/3, BRAF, EIF4EBP1, FRAP1, MAPK1/3, PIK3CA/B/D/G, PIK3R1/2/3/5, and RPS6KB1/2) that could act as mediators.
More than USD 1.8 billion have been invested in drug development [19] over more than 12 years, but the FDA approval rate is only 2.01% [20,21]. Drug repositioning, which refers to reuse of existing approved drugs to treat different diseases, is one cost-effective alternative to drug development [22]. One of the proposed methods is computational-based drug repositioning, which computes the similarity of chemical components, physical binding of proteins, and topologically and differentially expressed gene information [23,24,25,26]. Recently, network-based computational models have revealed drug–disease interactions and drug–target relationships for drug repositioning; those methods require a priori information, such as protein–protein interactions [27].
In this study, we examine the systematic effects of sirolimus through gene–gene interactions in the ERBB signaling pathway using computational methods with no a priori information.
2. Materials and Methods
2.1. Materials
We downloaded two sets of MCF7 breast cancer cell line data for computational approaches. First, we downloaded a dataset, GSE5258, updated in 2017 from the gene expression omnibus (GEO) [28]. The datasets included a total of 218 samples divided into 182 drug-induced cancer cell lines and 36 control samples from the Affymetrix HT human genome U133A array platform. Datasets were preprocessed by elimination of unknown gene names and averaging of duplicate gene names, resulting in 13,235 unique gene names from 22,268 probes. To test our new method, we selected sirolimus-induced datasets with a sample size of seven. Another dataset was extracted from a different cohort, GSE116447 updated in 2018 from GEO, which was generated from the Affymetrix Human Genome U133A2.0 array platform with 22,277 probes [29]. We extracted 79 ERBB signaling genes from the GSE116447 dataset to validate the proposed method.
2.2. Computational Methods for Obtaining Pathway and Gene Interactions
Computational methods include two parts: obtaining enrichment pathways and inferring gene–gene interactions. First, to obtain the enrichment pathway affected by a given drug, we adopted gene set enrichment analysis (GSEA) [30], which sorted pathways by enrichment score. The enrichment score was computed by the maximum deviation from zero encountered in the random walk corresponding to the weighted Kolmogorov–Smirnov-like statistic for comparing drug-induced datasets to normal datasets.
Next, an inverse algorithm was adopted to discover gene–gene interactions using only drug-induced datasets. The inverse algorithm [31] consisted of four subroutines. First, to perform an optimization approach for search direction, a quasi-Newton method based on an approximated Hessian matrix minimizing quadratic Newton steps (BFGS) was employed. The performance of BFGS in optimization was proven in previous studies [32]. Second, line search was used to determine the subsequent direction for the parameters. Next, an objective function provided the criterion for determining either termination or iteration of the algorithm based on error integration between real and synthetic datasets. The objective function used for this algorithm was norm, which can be rewritten as . Finally, a mathematical drug model was used in matrix form, following the works reported in [32,33,34].
The method used in this study was examined and validated in our previous research [32,35]. We used cancer datasets only for identifying gene–gene networks (parameters in the inverse algorithm) in the enrichment pathways; however, parameter prediction with an inverse algorithm requires both real and synthetic datasets. To generate synthetic datasets, noise was added to the original microarray datasets using the following equation:
, where . In each iteration, is newly updated as the synthetic data for .
To validate our computational results, we used another dataset, GSE116447, whose description and validation dataset are shown in Figure 1.
3. Results
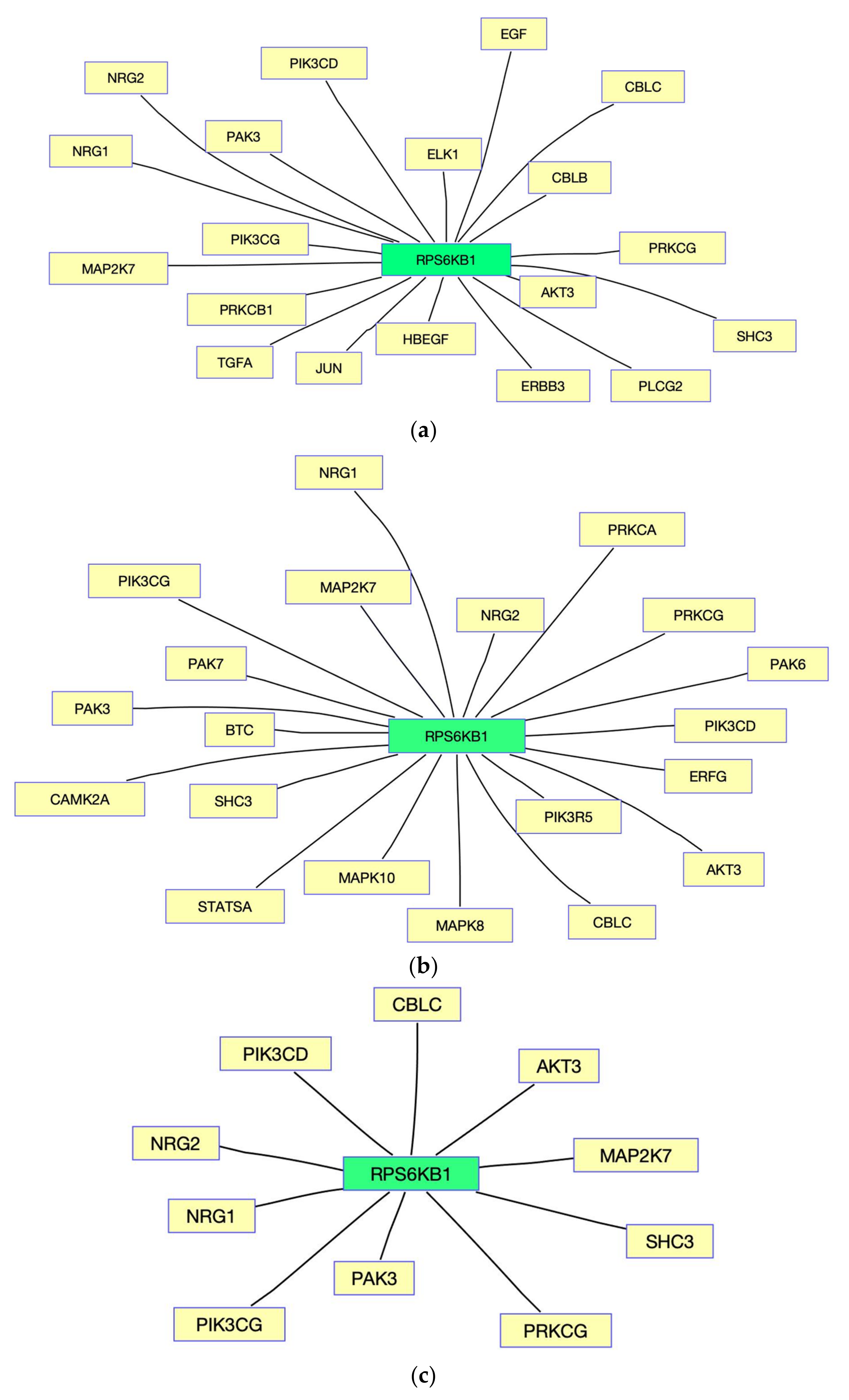
Both the ERBB signaling pathway and the mTOR signaling pathway are associated with cancer development. GSEA was conducted by using MCF7 breast cancer datasets comparing samples with sirolimus treatment and samples without sirolimus treatment. We obtained the top three pathways, which were cellular homeostasis-related cysteine metabolism, prion disease, and the ERBB signaling pathway. Even though the mTOR signaling pathway is a well-known target of sirolimus, it was not included in top enrichment pathways from our GSEA performance. Instead of the mTOR signaling pathway, therefore, we considered the ERBB signaling pathway as the pathway most affected by sirolimus. Additionally, we observed 18 overlapping genes between the mTOR pathway, which consisted of 48 genes, and the ERBB signaling pathway, which consisted of 87 genes, based on the KEGG pathway [15]. Among the 18 genes (AKT1/2/3, BRAF, EIF4EBP1, FRAP, MAPK1/3, PIK3CA/B/D/G, PIK3R1/2/3/5, and RPS6KB1/2), 3 genes, including PIK3, AKT, and RPS6KB1, are known targets of rapamycin. With 82 genes, after filtering in the ERBB signaling pathway, we performed an inverse algorithm by using cancer datasets only. The relationships between RPS6KB1 and 19 genes, as shown in Figure 2a, were derived from the inverse algorithm. For GSE116447 datasets, an inverse algorithm was applied to 79 genes (after filtering) in the ERBB signaling pathway, as in Figure 2b. There were 20 connections between the genes, with RPS6KB1 as a hub gene.
Interaction analyses among the affected genes were conducted to determine how the drug exerts systemic effects through gene interaction. Our conclusion was that RPS6KB1 is a hub gene for each 20 genes of the two datasets (Figure 2a,b) and shared 10 genes, including AKT3, CBLC, MAP2K7, NRG1/2, PAK3, PIK3CD/G, PRKCG, and SHC3, as shown in Figure 2c.
4. Discussion
In breast cancer therapy, it is well established that rapamycin, a cytostatic agent, regulates mTOR signaling networks via its downstream target, called RPS6KB1 [34]. Additionally, the AKT and ERBB signaling pathways are also involved in cancer cell proliferation, cell survival and cell growth. Yet, little is known regarding systemic gene interactions in the pathways. In this paper, we suggest a systemic effect of sirolimus (rapamycin) in the ERBB signaling pathway that is obtained from GSEA. By conducting the computational analysis of two independent datasets, GSE5258 and GSE116447, we identified a hub gene RPS6KB1 and its related genes, including AKT3, CBLC, MAP2K7, NRG1, NRG2, PAK3, PIK3CD, PIK3CG, PRKCG, and SHC3, involved in breast cancer.
PIK3 and AKT are upstream targets of the mTOR pathway. The PIK3/AKT/mTOR pathway plays a critical role in mediating multiple cellular functions, including cell growth, proliferation, metabolism, survival and angiogenesis via its downstream target RPS6KB1 [36]. NRG1 is known as a candidate oncogene and a tumor suppressor gene in breast cancer [37]. Novel to the study is that the effect of sirolimus may take place by the ERBB signaling pathway and its downstream target RPS6KB1. Furthermore, we discovered that sirolimus significantly affects the expression of genes, such as PAK3, AKT3, MAP2K7 and NRG1, which are regulated by RPS6KB1 via the ERBB signaling pathway, implying crosstalk with the mTOR pathway.
P21-activated kinases (PAKs) have roles in cell cycle, cell motility, and angiogenesis, and are mostly overexpressed in several cancers, such as glioma, lung, breast and colorectal [38]. Yet, the mechanism of PAKs is controversial. For example, in one study, p21-activiated kinase 3 (PAK3) was overexpressed in neuroendocrine cancer [38,39], while another study suggested that significantly decreased PAK3 expression was detected in gastric cancer cells [40]. In our study, PAK3 was upregulated by rapamycin treatment and by the inhibition of RPS6KB1. In addition, sirolimus affects CBLC, MAP2K7, NRG1/2, PAK3, SHC3, and PRKCG, which are not involved in the mTOR pathway, through RPS6KB1 in the ERBB pathway, implying that sirolimus is also involved in the ERBB signaling pathway.
This study has the following limitation: the current findings of our study remain to be experimentally confirmed. Caution is necessary in interpretating or determining the functional roles of rapamycin for the pathology of breast cancers.
5. Conclusions
In this study, we explored the systemic role of sirolimus, also known as rapamycin, via the ERBB signaling pathway, which plays an important role in cancer development. In computational analysis with MCF7 breast cancer cells, we found the 10 most effected genes through RPS6KB1 within the ERBB signaling pathway. The current findings of the study suggest that the therapeutic effect of rapamycin for breast cancer is mediated by RPS6KB1 and its interaction with the ERBB signaling pathway. In addition, the proposed computational methodology for determining systemic drug influences may be a useful approach to determine drug repositioning for diseases that have the same pattern in the future.
Funding
This work was supported by Sangmyung University (2021-A000-0364).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data can be freely downloaded from GEO datasets. (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE5258, accessed on 17 June 2021 and https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE116447, accessed on 17 June 2021).
Conflicts of Interest
The author declares no conflict of interest.
References
- Li, J.; Kim, S.G.; Blenis, J. Rapamycin: One drug, many effects. Cell Metab. 2014, 19, 373–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 169, 361–371. [Google Scholar] [CrossRef]
- Chung, J.; Kuo, C.J.; Crabtree, G.R.; Blenis, J. Rapamycin-FKBP specifically blocks growth-dependent activation of and signaling by the 70 kd S6 protein kinases. Cell 1992, 69, 1227–1236. [Google Scholar] [CrossRef]
- Kuo, C.J.; Chung, J.; Fiorentino, D.F.; Flanagan, W.M.; Blenis, J.; Crabtree, G.R. Rapamycin selectively inhibits interleukin-2 activation of p70 S6 kinase. Nature 1992, 358, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Price, D.J.; Grove, J.R.; Calvo, V.; Avruch, J.; Bierer, B.E. Rapamycin-induced inhibition of the 70-kilodalton S6 protein kinase. Science 1992, 257, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [Green Version]
- Loewith, R.; Hall, M.N. Target of rapamycin (TOR) in nutrient signaling and growth control. Genetics 2011, 189, 1177–1201. [Google Scholar] [CrossRef] [Green Version]
- Malaguti, P.; Vari, S.; Cognetti, F.; Fabi, A. The Mammalian target of rapamycin inhibitors in breast cancer: Current evidence and future directions. Anticancer Res. 2013, 33, 21–28. [Google Scholar]
- Wander, S.A.; Hennessy, B.T.; Slingerland, J.M. Next-generation mTOR inhibitors in clinical oncology: How pathway complexity informs therapeutic strategy. J. Clin. Investig. 2011, 121, 1231–1241. [Google Scholar] [CrossRef] [Green Version]
- Heinonen, H.; Nieminen, A.; Saarela, M.; Kallioniemi, A.; Klefstrom, J.; Hautaniemi, S.; Monni, O. Deciphering downstream gene targets of PI3K/mTOR/p70S6K pathway in breast cancer. BMC Genom. 2008, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hare, S.H.; Harvey, A.J. mTOR function and therapeutic targeting in breast cancer. Am. J. Cancer Res. 2017, 7, 383–404. [Google Scholar] [PubMed]
- Frogne, T.; Benjaminsen, R.V.; Sonne-Hansen, K.; Sorensen, B.S.; Nexo, E.; Laenkholm, A.V.; Rasmussen, L.M.; Riese, D.J., 2nd; de Cremoux, P.; Stenvang, J.; et al. Activation of ErbB3, EGFR and Erk is essential for growth of human breast cancer cell lines with acquired resistance to fulvestrant. Breast Cancer Res. Treat. 2009, 114, 263–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, M.; Grundker, C.; Fister, S.; Kubin, J.; Wilkens, L.; Mueller, M.D.; Hemmerlein, B.; Emons, G.; Gunthert, A.R. Inhibition of the AKT/mTOR and erbB pathways by gefitinib, perifosine and analogs of gonadotropin-releasing hormone I and II to overcome tamoxifen resistance in breast cancer cells. Int. J. Oncol. 2012, 41, 1845–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaturvedi, D.; Gao, X.; Cohen, M.S.; Taunton, J.; Patel, T.B. Rapamycin induces transactivation of the EGFR and increases cell survival. Oncogene 2009, 28, 1187–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Rouillard, A.D.; Gundersen, G.W.; Fernandez, N.F.; Wang, Z.; Monteiro, C.D.; McDermott, M.G.; Ma’ayan, A. The harmonizome: A collection of processed datasets gathered to serve and mine knowledge about genes and proteins. Database 2016, 2016, 1–16. [Google Scholar] [CrossRef]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Forner, O.; Marin-Garcia, P.; Arnau, V.; D’Eustachio, P.; Stein, L.; Hermjakob, H. Reactome pathway analysis: A high-performance in-memory approach. BMC Bioinform. 2017, 18, 142. [Google Scholar] [CrossRef] [Green Version]
- Fabregat, A.; Sidiropoulos, K.; Garapati, P.; Gillespie, M.; Hausmann, K.; Haw, R.; Jassal, B.; Jupe, S.; Korninger, F.; McKay, S.; et al. The Reactome pathway Knowledgebase. Nucleic Acids Res. 2016, 44, D481–D487. [Google Scholar] [CrossRef] [Green Version]
- Ashburn, T.T.; Thor, K.B. Drug repositioning: Identifying and developing new uses for existing drugs. Nat. Rev. Drug Discov. 2004, 3, 673–683. [Google Scholar] [CrossRef]
- Pammolli, F.; Magazzini, L.; Riccaboni, M. The productivity crisis in pharmaceutical R&D. Nat. Rev. Drug Discov. 2011, 10, 428–438. [Google Scholar]
- Yeu, Y.; Yoon, Y.; Park, S. Protein localization vector propagation: A method for improving the accuracy of drug repositioning. Mol. Biosyst. 2015, 11, 2096–2102. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, S.; Zhang, X.; Li, J. Drug repositioning by integrating target information through a heterogeneous network model. Bioinformatics 2014, 30, 2923–2930. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, F.; Carrella, D.; Mandriani, B.; Pisonero-Vaquero, S.; Sirci, F.; Medina, D.L.; Brunetti-Pierri, N.; di Bernardo, D. gene2drug: A computational tool for pathway-based rational drug repositioning. Bioinformatics 2018, 34, 1498–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laenen, G.; Thorrez, L.; Bornigen, D.; Moreau, Y. Finding the targets of a drug by integration of gene expression data with a protein interaction network. Mol. Biosyst. 2013, 9, 1676–1685. [Google Scholar] [CrossRef]
- Isik, Z.; Baldow, C.; Cannistraci, C.V.; Schroeder, M. Drug target prioritization by perturbed gene expression and network information. Sci. Rep. 2015, 5, 17417. [Google Scholar] [CrossRef] [PubMed]
- Kim, S. A New Computational Approach to Evaluating Systemic Gene–Gene Interactions in a Pathway Affected by Drug LY294002. Processes 2020, 8, 1230. [Google Scholar] [CrossRef]
- Badkas, A.; De Landtsheer, S.; Sauter, T. Topological network measures for drug repositioning. Brief. Bioinform. 2021, 22, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lamb, J.; Crawford, E.D.; Peck, D.; Modell, J.W.; Blat, I.C.; Wrobel, M.J.; Lerner, J.; Brunet, J.P.; Subramanian, A.; Ross, K.N.; et al. The Connectivity Map: Using gene-expression signatures to connect small molecules, genes, and disease. Science 2006, 313, 1929–1935. [Google Scholar] [CrossRef] [Green Version]
- Monks, A.; Zhao, Y.; Hose, C.; Hamed, H.; Krushkal, J.; Fang, J.; Sonkin, D.; Palmisano, A.; Polley, E.C.; Fogli, L.K.; et al. The NCI Transcriptional Pharmacodynamics Workbench: A Tool to Examine Dynamic Expression Profiling of Therapeutic Response in the NCI-60 Cell Line Panel. Cancer Res. 2018, 78, 6807–6817. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Press, W.H. Numerical Recipes in C: The Art of Scientific Computing, 2nd ed.; Cambridge University Press: Cambridge, UK, 1992; pp. 425–430. [Google Scholar]
- Kim, S.; Kim, J.; Cho, K.H. Inferring gene regulatory networks from temporal expression profiles under time-delay and noise. Comput. Biol. Chem. 2007, 31, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kreider, K. Parameter identification for nonlinear elastic and viscoelastic plates. Appl. Numer. Math. 2006, 56, 1538–1554. [Google Scholar] [CrossRef]
- Kim, S. Identifying dynamic pathway interactions based on clinical information. Comput. Biol. Chem. 2017, 68, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Kim, S. A Computational Approach for Pathway-Based Systemic Drug Influence. Processes 2021, 9, 1063. [Google Scholar] [CrossRef]
- Vignot, S.; Faivre, S.; Aguirre, D.; Raymond, E. mTOR-targeted therapy of cancer with rapamycin derivatives. Ann. Oncol. 2005, 16, 525–537. [Google Scholar] [CrossRef]
- Chua, Y.L.; Ito, Y.; Pole, J.C.; Newman, S.; Chin, S.F.; Stein, R.C.; Ellis, I.O.; Caldas, C.; O’Hare, M.J.; Murrell, A.; et al. The NRG1 gene is frequently silenced by methylation in breast cancers and is a strong candidate for the 8p tumour suppressor gene. Oncogene 2009, 28, 4041–4052. [Google Scholar] [CrossRef] [Green Version]
- Ye, D.Z.; Field, J. PAK signaling in cancer. Cell Logist. 2012, 2, 105–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.-X.; Wang, W.-Q.; Ye, L.; Bi, Y.-F.; Fang, H.; Cui, B.; Zhou, W.-W.; Dai, M.; Zhang, J.; Li, X.-Y. p21-activated kinase 3 is overexpressed in thymic neuroendocrine tumors (carcinoids) with ectopic ACTH syndrome and participates in cell migration. Endocrine 2010, 38, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.J.; Yu, T.Y.; Li, Y.R.; Liu, Y.J.; Deng, B.B. Circ_0000190 suppresses gastric cancer progression potentially via inhibiting miR-1252/PAK3 pathway. Cancer Cell Int. 2020, 20, 351. [Google Scholar] [CrossRef]
Figure 1.
Overall schematic flowchart. Red arrows represent validation results using GSE116447; black arrows represent the results of GSE5258.
Figure 1.
Overall schematic flowchart. Red arrows represent validation results using GSE116447; black arrows represent the results of GSE5258.

Figure 2.
(a) Results of the inverse algorithm using the GSE3921 dataset; (b) GSE116447 dataset; (c) Overlapping gene interactions from both results A and B.
Figure 2.
(a) Results of the inverse algorithm using the GSE3921 dataset; (b) GSE116447 dataset; (c) Overlapping gene interactions from both results A and B.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, S. Systemic Efficacy of Sirolimus via the ERBB Signaling Pathway in Breast Cancer. Processes 2022, 10, 552. https://doi.org/10.3390/pr10030552
AMA Style
Kim S. Systemic Efficacy of Sirolimus via the ERBB Signaling Pathway in Breast Cancer. Processes. 2022; 10(3):552. https://doi.org/10.3390/pr10030552
Chicago/Turabian StyleKim, Shinuk. 2022. "Systemic Efficacy of Sirolimus via the ERBB Signaling Pathway in Breast Cancer" Processes 10, no. 3: 552. https://doi.org/10.3390/pr10030552
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.