
Immune Redox Modulation Effects of Non-Electrolyzed Hypochlorous Acid Water on Helicobacter pylori-Infected C57BL/6 Mouse Model

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
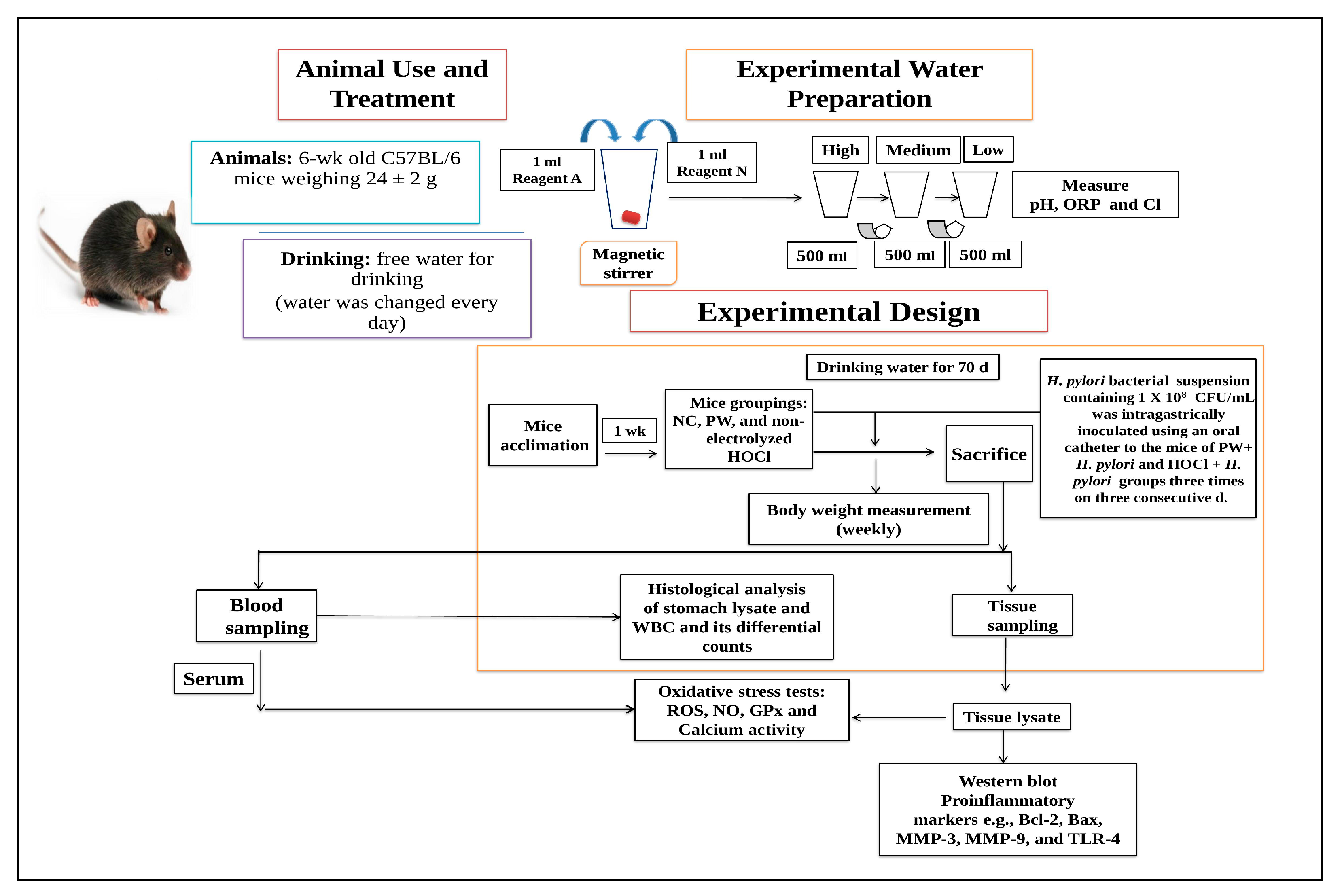
2.1. Animal Groupings and Treatment Procedure
2.2. Water Preparation and Their Properties
2.3. H. pylori Infection in Mice
2.4. Serum and Stomach Tissue Lysates Sample Preparation
2.5. Measurement of Body Weight
2.6. Histological Examination by Giemsa Staining
2.7. Total and Differential White Blood Cell (WBC) Counts
2.8. Measurement of Total Reactive Oxygen Species (ROS)
2.9. Measurement of Nitric Oxide (NO) Levels
2.10. Measurement of Glutathione Peroxidase (GPx) Activity
2.11. Detection of Intracellular Ca2+ Activity
2.12. Measurement of Proinflammatory Cytokines
2.13. Western Blot Analysis
2.14. Data Management and Statistical Analysis
3. Results
3.1. Effects of Non-Electrolyzed HOCl Water on the Body Weight of H. pylori-Infected C57BL/6 Mice
3.2. Histological Observation of the Stomach of H. pylori-Infected C57BL/6 Mice
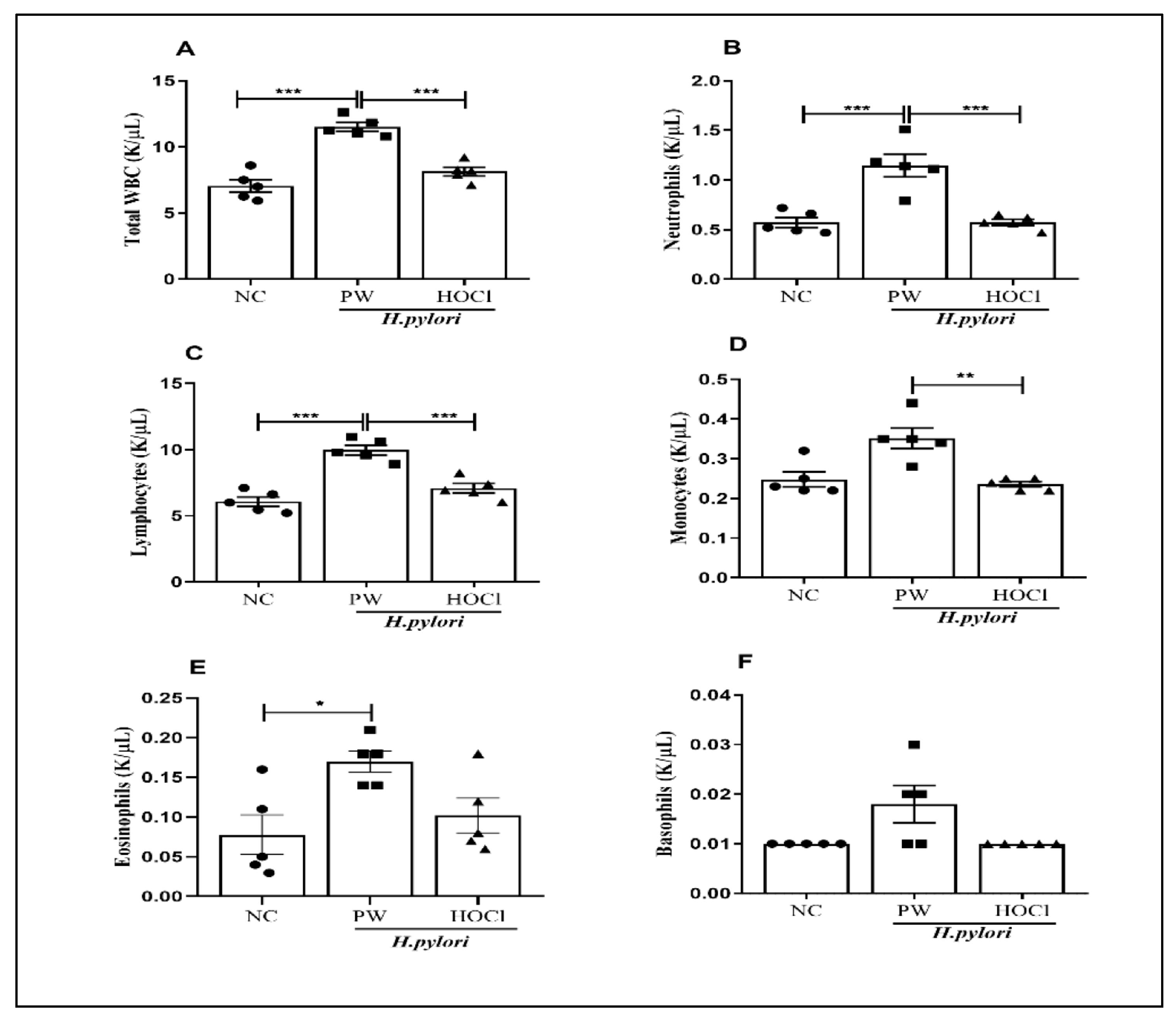
3.3. Effects of Non-Electrolyzed HOCl Water in H. pylori-Infected C57BL/6 Mice on Total WBC and Its Differential Counts
3.4. Effects of Non-Electrolyzed HOCl Water on OS Markers in H. pylori-Infected
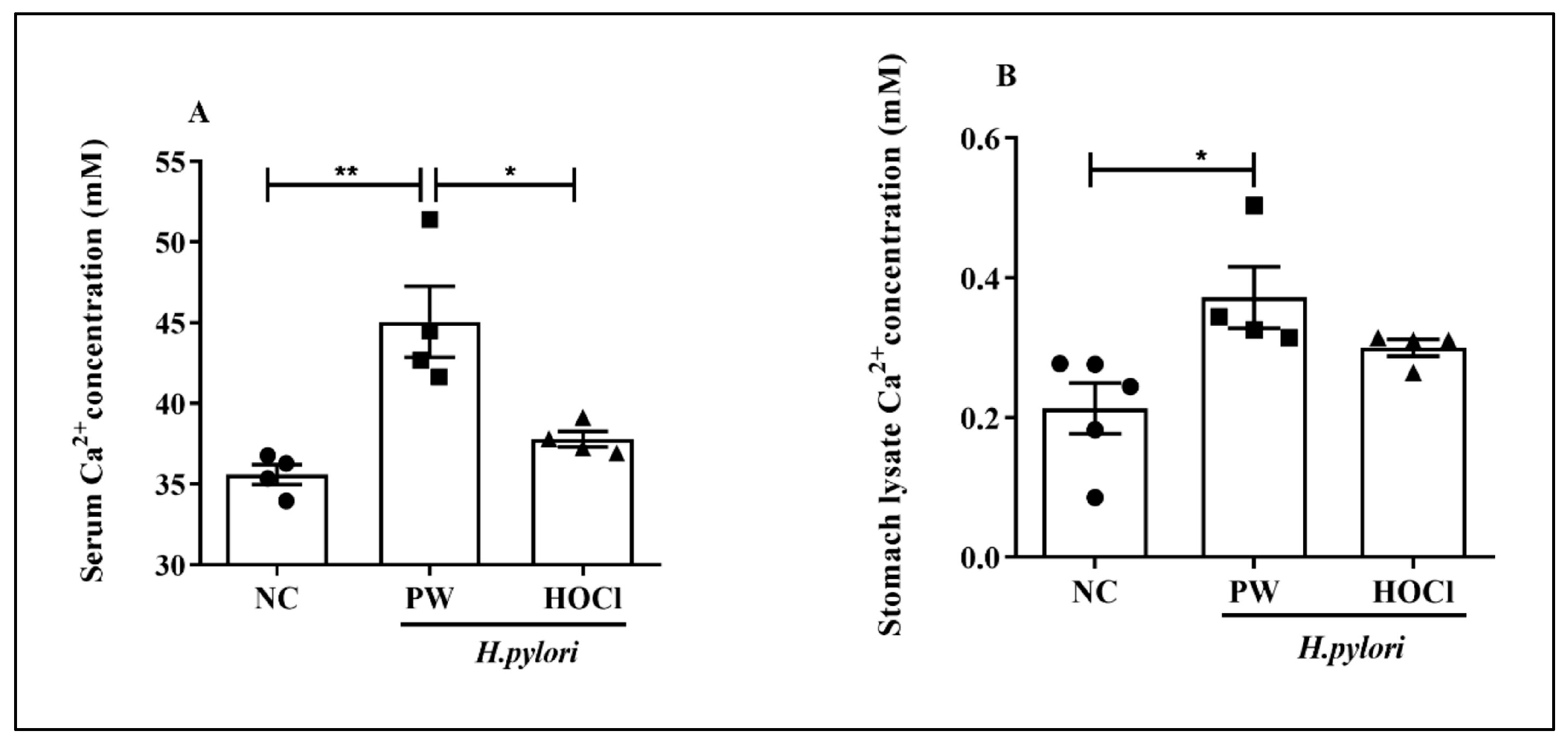
3.5. Effects of Non-Electrolyzed HOCl Water on the Intracellular Ca2+ Activity in H. pylori-Infected C57BL/6 Mice
3.6. Effects of Non-Electrolyzed HOCl on the Level of Inflammatory Cytokines in H. pylori-Infected C57BL/6 Mice
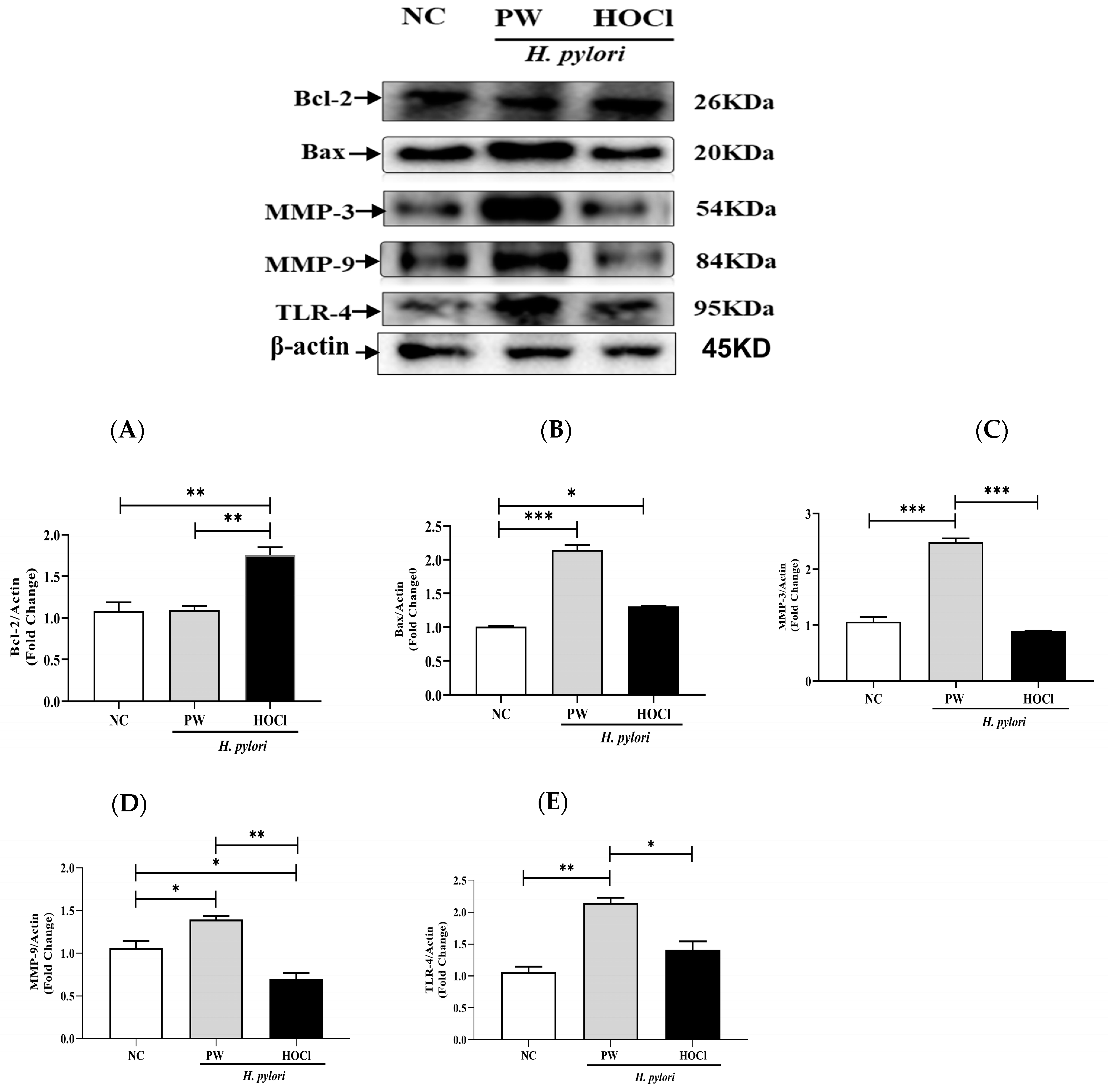
3.7. Effects of Non-Electrolyzed HOCl on Bcl-2, Bax, MMP-3, MMP-9, and TLR-4 in Stomach Lysates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Young, S.B.; Setlow, P. Mechanisms of Killing of Bacillus Subtilis Spores by Hypochlorite and Chlorine Dioxide. J. Appl. Microbiol. 2003, 95, 54–67. [Google Scholar] [CrossRef]
- Hori, T.; Yamamoto, H.; Harada, H.; Yamamoto, M.; Yamada, M.; Yazawa, T.; Sasaki, B.; Tani, M.; Sato, A.; Katsura, H.; et al. Multiple Gastric Carcinomas Associated with Epstein-Barr Virus and Helicobacter Pylori: A Thought-Provoking Case. Am. J. Case Rep. 2021, 22, e931668. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Atherton, J.C. The Spectrum of Helicobacter—Mediated Diseases. Annu. Rev. Pathol. Mech. Dis. 2021, 16, 123–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, L.; Deng, H.; Li, H.; Tang, W.; Guan, L.; Qiu, Y.; Donovan, M.J.; Chen, Z.; Tan, W. In Vivo Activation of PH-Responsive Oxidase-like Graphitic Nanozymes for Selective Killing of Helicobacter Pylori. Nat. Commun. 2021, 12, 2002. [Google Scholar] [CrossRef]
- Suzuki, T.; Kato, K.; Ohara, S.; Noguchi, K.; Sekine, H.; Nagura, H.; Shimosegawa, T. Localization of Antigen-Presenting Cells in Helicobacter Pylori-Infected Gastric Mucosa. Pathol. Int. 2002, 52, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Sipowicz, M.A.; Chomarat, P.; Diwan, B.A.; Anver, M.A.; Awasthi, Y.C.; Ward, J.M.; Rice, J.M.; Kasprzak, K.S.; Wild, C.P.; Anderson, L.M. Increased Oxidative DNA Damage and Hepatocyte Overexpression of Specific Cytochrome P450 Isoforms in Hepatitis of Mice Infected with Helicobacter Hepaticus. Am. J. Pathol. 1997, 151, 933–941. [Google Scholar] [PubMed]
- Ding, S.Z.; Minohara, Y.; Xue, J.F.; Wang, J.; Reyes, V.E.; Patel, J.; Dirden-Kramer, B.; Boldogh, I.; Ernst, P.B.; Crowe, S.E. Helicobacter Pylori Infection Induces Oxidative Stress and Programmed Cell Death in Human Gastric Epithelial Cells. Infect. Immun. 2007, 75, 4030–4039. [Google Scholar] [CrossRef]
- Farinati, F.; Cardin, R.; Degan, P.; Rugge, M.; Di Mario, F.; Bonvicini, P.; Naccarato, R. Oxidative DNA Damage Accumulation in Gastric Carcinogenesis. Gut 1998, 42, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Drake, I.M.; Mapstone, N.P.; Schorah, C.J.; White, K.L.M.; Chalmers, D.M.; Dixon, M.F.; Axon, A.T.R. Reactive Oxygen Species Activity and Lipid Peroxidation in Helicobacter Pylori Associated Gastritis: Relation to Gastric Mucosal Ascorbic Acid Concentrations and Effect of H Pylori Eradication. Gut 1998, 42, 768–771. [Google Scholar] [CrossRef]
- Davies, G.R.; Simmonds, N.J.; Stevens, T.R.J.; Sheaff, M.T.; Banatvala, N.; Laurenson, I.F.; Blake, D.R.; Rampton, D.S. Helicobacter Pylori Stimulates Antral Mucosal Reactive Oxygen Metabolite Production in vivo. Gut 1994, 35, 179–185. [Google Scholar] [CrossRef]
- Zhang, Q.B.; Dawodu, J.B.; Husain, A.; Etolhi, G.; Gemmell, C.G.; Russell, R.I. Association of Antral Mucosal Levels of Interleukin 8 and Reactive Oxygen Radicals in Patients Infected with Helicobacter Pylori. Clin. Sci. 1997, 92, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Argent, R.H.; Atherton, J.C. The Inflammatory and Immune Response to Helicobacter Pylori Infection. Best Pract. Res. Clin. Gastroenterol. 2007, 21, 237–259. [Google Scholar] [CrossRef]
- Petersen, O.H. Localization and Regulation of Ca2+ Entry and Exit Pathways in Exocrine Gland Cells. Cell Calcium 2003, 33, 337–344. [Google Scholar] [CrossRef]
- Pallagi, P.; Madácsy, T.; Varga, Á.; Maléth, J. Intracellular Ca2+ Signalling in the Pathogenesis of Acute Pancreatitis: Recent Advances and Translational Perspectives. Int. J. Mol. Sci 2020, 21, 4005. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.H.; Wong, J.H.; Chang, C.M.; Chen, B.K.; Tsai, Y.T.; Chen, W.C.; Wang, E.T.; Hsu, W.L.; Chang, W.C. Involvement of Intracellular Calcium Mobilization in IL-8 Activation in Human Retinal Pigment Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2015, 56, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Pierzchalski, P.; Konturek, S.J.; Meixner, H.; Faller, G.; Kirchner, T.; Hahn, E.G. Helicobacter Pylori Induces Apoptosis in Gastric Mucosa through an Upregulation of Bax Expression in Humans. Scand. J. Gastroenterol. 1999, 34, 375–383. [Google Scholar] [CrossRef]
- Konturek, P.C.; Konturek, S.J.; Brzozowski, T. Helicobacter Pylori Infection in Gastric Cancerogenesis. J. Physiol. Pharmacol. 2009, 60, 3–21. [Google Scholar]
- Lima, V.P.; de Lima, M.A.P.; André, A.R.; Ferreira, M.V.P.; Barros, M.A.P.; Rabenhorst, S.H.B. H Pylori (CagA) and Epstein-Barr Virus Infection in Gastric Carcinomas: Correlation With P53 Mutation and c-Myc, Bcl-2 and Bax Expression. World J. Gastroenterol. 2008, 14, 884–891. [Google Scholar] [CrossRef]
- Sokolova, O.; Naumann, M. Matrix Metalloproteinases in Helicobacter Pylori—Associated Gastritis and Gastric Cancer. Int. J. Mol. Sci. 2022, 23, 1883. [Google Scholar] [CrossRef]
- Smith, S.M. Role of Toll-like Receptors in Helicobacter Pylori Infection and Immunity. World J. Gastrointest. Pathophysiol. 2014, 5, 133–146. [Google Scholar] [CrossRef]
- O’Connor, A.; Molina-Infante, J.; Gisbert, J.P.; O’Morain, C. Treatment of Helicobacter Pylori Infection 2013. Helicobacter 2013, 18, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Medina-Tamayo, J.; Sánchez-Miranda, E.; Balleza-Tapia, H.; Ambriz, X.; Cid, M.E.; González-Espinosa, D.; Gutiérrez, A.A.; González-Espinosa, C. Super-Oxidized Solution Inhibits IgE-Antigen-Induced Degranulation and Cytokine Release in Mast Cells. Int. Immunopharmacol. 2007, 7, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Del Rosso, J.Q.; Bhatia, N. Status Report on Topical Hypochlorous Acid: Clinical Relevance of Specific Formulations, Potential Modes of Action, and Study Outcomes. J. Clin. Aesthetic Dermatol. 2018, 11, 36–39. [Google Scholar]
- De-Oliveira, R.B.B.; Kato, E.E.; Soares de Lima, T.; Alba-Loureiro, T.C.; Curi, R.; Cirillo, M.C.; Sampaio, S.C. Crotoxin-Treated Macrophages Stimulate ROS Production and Killing Activity in Co-Cultured Neutrophils Article Information. J. Appl. Microbiol. Res. 2018, 1, 2018050322. [Google Scholar]
- Mentis, A.F.A.; Boziki, M.; Grigoriadis, N.; Papavassiliou, A.G. Helicobacter Pylori Infection and Gastric Cancer Biology: Tempering a Double-Edged Sword. Cell. Mol. Life Sci. 2019, 76, 2477–2486. [Google Scholar] [CrossRef] [PubMed]
- Kubota, A.; Goda, T.; Tsuru, T.; Yonekura, T.; Yagi, M.; Kawahara, H.; Yoneda, A.; Tazuke, Y.; Tani, G.; Ishii, T.; et al. Efficacy and Safety of Strong Acid Electrolyzed Water for Peritoneal Lavage to Prevent Surgical Site Infection in Patients with Perforated Appendicitis. Surg. Today 2015, 45, 876–879. [Google Scholar] [CrossRef]
- Wang, L.; Bassiri, M.; Najafi, R.; Najafi, K.; Yang, J.; Khosrovi, B.; Hwong, W.; Barati, E.; Belisle, B.; Celeri, C.; et al. Hypochlorous Acid as a Potential Wound Care Agent: Part I. Stabilized Hypochlorous Acid: A Component of the Inorganic Armamentarium of Innate Immunity. J. Burn. Wounds 2007, 6, e5. [Google Scholar]
- Takeda, Y.; Matsuda, S.; Jamsransuren, D.; Ogawa, H. Comparison of the SARS-CoV-2-Inactivating Activities of the Differently Manufactured Hypochlorous Acid Water Products with Various PH. J. Water Health 2021, 19, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.S.; Bajgai, J.; You, I.S.; Rahman, M.H.; Fadriquela, A.; Sharma, S.; Kwon, H.U.; Lee, S.Y.; Kim, C.S.; Lee, K.J. Therapeutic Effects of Hydrogen Gas Inhalation on Trimethyltin-Induced Neurotoxicity and Cognitive Impairment in the C57BL/6 Mice Model. Int. J. Mol. Sci. 2021, 22, 13313. [Google Scholar] [CrossRef]
- Jiménez, F.T.; Bayona, C.T. Molecular Pathophysiology in Infection by Helicobacter Pylori. Salud Uninorte 2021, 32, 500–512. [Google Scholar] [CrossRef]
- Peek, R.M.; Fiske, C.; Wilson, K.T. Role of Innate Immunity in Helicobacter Pylori-Induced Gastric Malignancy. Physiol Rev. 2010, 90, 831–858. [Google Scholar] [CrossRef]
- Iram, A.; Wang, X.; Demirci, A. Electrolyzed Oxidizing Water and Its Applications as Sanitation and Cleaning Agent. Food Eng. Rev. 2021, 13, 411–427. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, Y.; Gao, L.; Huang, J.; Yang, X.; Pei, L.; Ye, Z.; Zhu, L. Acidic Electrolyzed Water Inhibits the Viability of Gardnerella Spp. via Oxidative Stress Response. Front. Med. 2022, 9, 817957. [Google Scholar] [CrossRef]
- Lucio-Sauceda, D.G.; Urrutia-Baca, V.H.; Gomez-Flores, R.; De La Garza-Ramos, M.A.; Tamez-Guerra, P.; Orozco-Flores, A. Antimicrobial and Anti-Biofilm Effect of an Electrolyzed Superoxidized Solution at Neutral-PH against Helicobacter pylori. BioMed Res. Int. 2019, 2019, 6154867. [Google Scholar] [CrossRef]
- Liu, C.; Li, X. Stage-Dependent Changes in Albumin, NLR, PLR, and AFR Are Correlated with Shorter Survival in Patients with Gastric Cancer. Clin. Lab. 2019, 65. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Suzuki, H.; Saya, H.; Hatakeyama, M.; Hirayama, T.; Hirata, K.; Nagano, O.; Matsuzaki, J.; Hibi, T. Reactive Oxygen Species-Induced Autophagic Degradation of Helicobacter Pylori CagA Is Specifically Suppressed in Cancer Stem-like Cells. Cell Host Microbe 2012, 12, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Gmünder, H.; Dröge, W. Differential Effects of Glutathione Depletion on T Cell Subsets. Cell. Immunol. 1991, 138, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Sheh, A.; Lee, C.W.; Masumura, K.; Rickman, B.H.; Nohmi, T.; Wogan, G.N.; Fox, J.G.; Schauer, D.B. Mutagenic Potency of Helicobacter Pylori in the Gastric Mucosa of Mice Is Determined by Sex and Duration of Infection. Proc. Natl. Acad. Sci. USA 2010, 107, 15217–15222. [Google Scholar] [CrossRef]
- Peek, R.M.; Blaser, M.J. Helicobacter Pylori and Gastrointestinal Tract Adenocarcinomas. Nat. Rev. Cancer 2002, 2, 28–37. [Google Scholar] [CrossRef]
- Jabeen, R.; Jami, A.; Shahab, A.; Shahab, A. Relationship of Halitosis with Gastric Helicobacter Pylori Infection. Pak. J. Med. Health Sci. 2021, 15, 2285–2288. [Google Scholar] [CrossRef]
- Kidane, D.; Murphy, D.L.; Sweasy, J.B. Accumulation of Abasic Sites Induces Genomic Instability in Normal Human Gastric Epithelial Cells during Helicobacter Pylori Infection. Oncogenesis 2014, 3, e128. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Yoshida, H.; Mitsuno, Y.; Hirata, Y.; Ogura, K.; Shiratori, Y.; Omata, M. Analysis of Apoptotic and Antiapoptotic Signalling Pathways Induced by Helicobacter Pylori. Gut 2002, 50, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Bhattacharya, G.; Stohs, S.J. Production of Reactive Oxygen Species by Gastric Cells in Association with Helicobacter Pylori. Free Radic. Res. 1996, 24, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Beswick, E.J.; Pinchuk, I.V.; Suarez, G.; Sierra, J.C.; Reyes, V.E. Helicobacter Pylori CagA-Dependent Macrophage Migration Inhibitory Factor Produced by Gastric Epithelial Cells Binds to CD74 and Stimulates Procarcinogenic Events. J. Immunol. 2006, 176, 6794–6801. [Google Scholar] [CrossRef]
- El-Omar, E.M.; Oien, K.; El-Nujumi, A.; Gillen, D.; Wirz, A.; Dahill, S.; Williams, C.; Ardill, J.E.S.; McColl, K.E.L. Helicobacter Pylori Infection and Chronic Gastric Acid Hyposecretion. Gastroenterology 1997, 113, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.Y. Helicobacter Pylori Infection in the Pathogenesis of Duodenal Ulcer and Gastric Cancer: A Model. Gastroenterology 1997, 113, 1983–1991. [Google Scholar] [CrossRef]
- Crabtree, J.E.; Court, M.; Aboshkiwa, M.A.; Jeremy, A.H.T.; Dixon, M.F.; Robinson, P.A. Gastric Mucosal Cytokine and Epithelial Cell Responses to Helicobacter Pylori Infection in Mongolian Gerbils. J. Pathol. 2004, 202, 197–207. [Google Scholar] [CrossRef]
- Rébé, C.; Ghiringhelli, F. Interleukin-1β and Cancer. Cancers 2020, 12, 1791. [Google Scholar] [CrossRef]
- Beales, I.L.P.; Calam, J. Interleukin 1β and Tumour Necrosis Factor α Inhibit Acid Secretion in Cultured Rabbit Parietal Cells by Multiple Pathways. Gut 1998, 42, 227–234. [Google Scholar] [CrossRef]
- Outlioua, A.; Badre, W.; Desterke, C.; Echarki, Z.; El Hammani, N.; Rabhi, M.; Riyad, M.; Karkouri, M.; Arnoult, D.; Khalil, A.; et al. Gastric IL-1β, IL-8, and IL-17A Expression in Moroccan Patients Infected with Helicobacter Pylori May Be a Predictive Signature of Severe Pathological Stages. Cytokine 2020, 126, 154893. [Google Scholar] [CrossRef]
- Basso, D.; Scrigner, M.; Toma, A.; Navaglia, F.; Di Mario, F.; Rugge, M.; Plebani, M. Helicobacter Pylori Infection Enhances Mucosal Interleukin-1β, Interleukin-6, and the Soluble Receptor of Interleukin-2. Int. J. Clin. Lab. Res. 1996, 26, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Fang, D.C.; Wang, R.Q.; Yang, S.M.; Liu, H.F.; Luo, Y.H. Effect of Helicobacter Pylori Infection on Expression of Bcl-2 Family Members in Gastric Adenocarcinoma. World J. Gastroenterol. 2004, 10, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Konturek, S.J.; Pierzchalski, P.; Bielański, W.; Duda, A.; Marlicz, K.; Starzyńska, T.; Hahn, E.G. Cancerogenesis in Helicobacter Pylori Infected Stomach—Role of Growth Factors, Apoptosis and Cyclooxygenases. Med. Sci. Monit. 2001, 7, 1092–1107. [Google Scholar]
- Li, S.L.; Zhao, J.R.; Ren, X.Y.; Xie, J.P.; Ma, Q.Z.; Rong, Q.H. Increased Expression of Matrix Metalloproteinase-9 Associated with Gastric Ulcer Recurrence. World J. Gastroenterol. 2013, 19, 4590–4595. [Google Scholar] [CrossRef]
- Minyaylo, O.; Ponomarenko, I.; Reshetnikov, E.; Dvornyk, V.; Churnosov, M. Functionally Significant Polymorphisms of the MMP-9 Gene Are Associated with Peptic Ulcer Disease in the Caucasian Population of Central Russia. Sci. Rep. 2021, 11, 13515. [Google Scholar] [CrossRef] [PubMed]
- Bergin, P.J.; Raghavan, S.; Svensson, H.; Starckx, S.; Van Aelst, I.; Gjertsson, I.; Opdenakker, G.; Quiding-Järbrink, M. Gastric Gelatinase B/Matrix Metalloproteinase-9 Is Rapidly Increased in Helicobacter Felis-Induced Gastritis. FEMS Immunol. Med. Microbiol. 2008, 52, 88–98. [Google Scholar] [CrossRef]
- Chichirau, B.E.; Scheidt, T.; Diechler, S.; Neuper, T.; Horejs-Hoeck, J.; Huber, C.G.; Posselt, G.; Wessler, S. Dissecting the Helicobacter Pylori-Regulated Transcriptome of b Cells. Pathog. Dis. 2020, 78, ftaa049. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | PW | HOCl |
|---|---|---|
| pH | 7.5 | 6.1 |
| ORP (mV) | 310 | 931 |
| ACC (ppm) | 0 | 51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.H.; Bajgai, J.; Cho, Y.; Fadriquela, A.; Sharma, S.; Thuy, T.T.; Cho, S.H.; Jeong, Y.J.; Goh, S.H.; Kim, Y.; et al. Immune Redox Modulation Effects of Non-Electrolyzed Hypochlorous Acid Water on Helicobacter pylori-Infected C57BL/6 Mouse Model. Processes 2023, 11, 1474. https://doi.org/10.3390/pr11051474
Rahman MH, Bajgai J, Cho Y, Fadriquela A, Sharma S, Thuy TT, Cho SH, Jeong YJ, Goh SH, Kim Y, et al. Immune Redox Modulation Effects of Non-Electrolyzed Hypochlorous Acid Water on Helicobacter pylori-Infected C57BL/6 Mouse Model. Processes. 2023; 11(5):1474. https://doi.org/10.3390/pr11051474
Chicago/Turabian StyleRahman, Md. Habibur, Johny Bajgai, Yoojin Cho, Ailyn Fadriquela, Subham Sharma, Trinh Thi Thuy, Syung Hyun Cho, Yun Ju Jeong, Seong Hoon Goh, Yundeok Kim, and et al. 2023. "Immune Redox Modulation Effects of Non-Electrolyzed Hypochlorous Acid Water on Helicobacter pylori-Infected C57BL/6 Mouse Model" Processes 11, no. 5: 1474. https://doi.org/10.3390/pr11051474
APA StyleRahman, M. H., Bajgai, J., Cho, Y., Fadriquela, A., Sharma, S., Thuy, T. T., Cho, S. H., Jeong, Y. J., Goh, S. H., Kim, Y., Kim, C. -S., & Lee, K. -J. (2023). Immune Redox Modulation Effects of Non-Electrolyzed Hypochlorous Acid Water on Helicobacter pylori-Infected C57BL/6 Mouse Model. Processes, 11(5), 1474. https://doi.org/10.3390/pr11051474