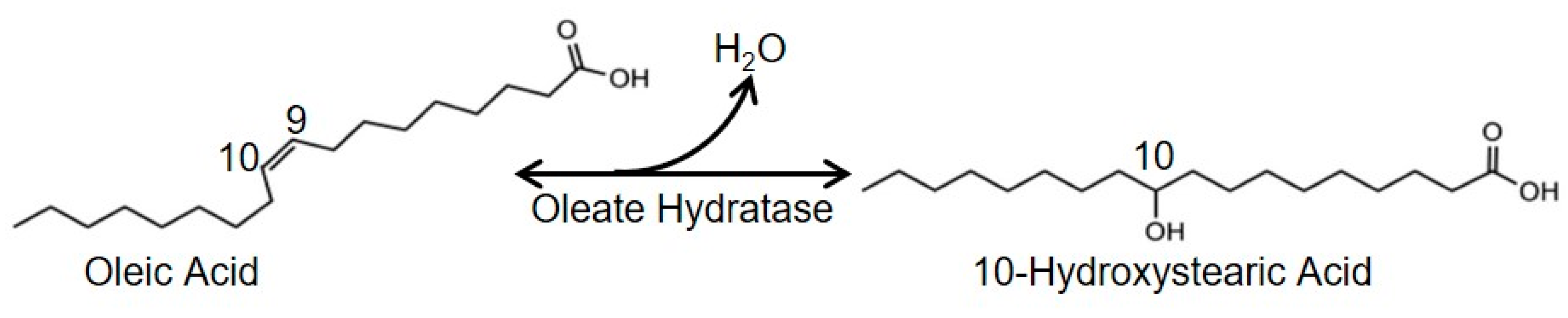
Sustainable Biotransformation of Oleic Acid to 10-Hydroxystearic Acid by a Recombinant Oleate Hydratase from Lactococcus garvieae

 ,
,  and
and 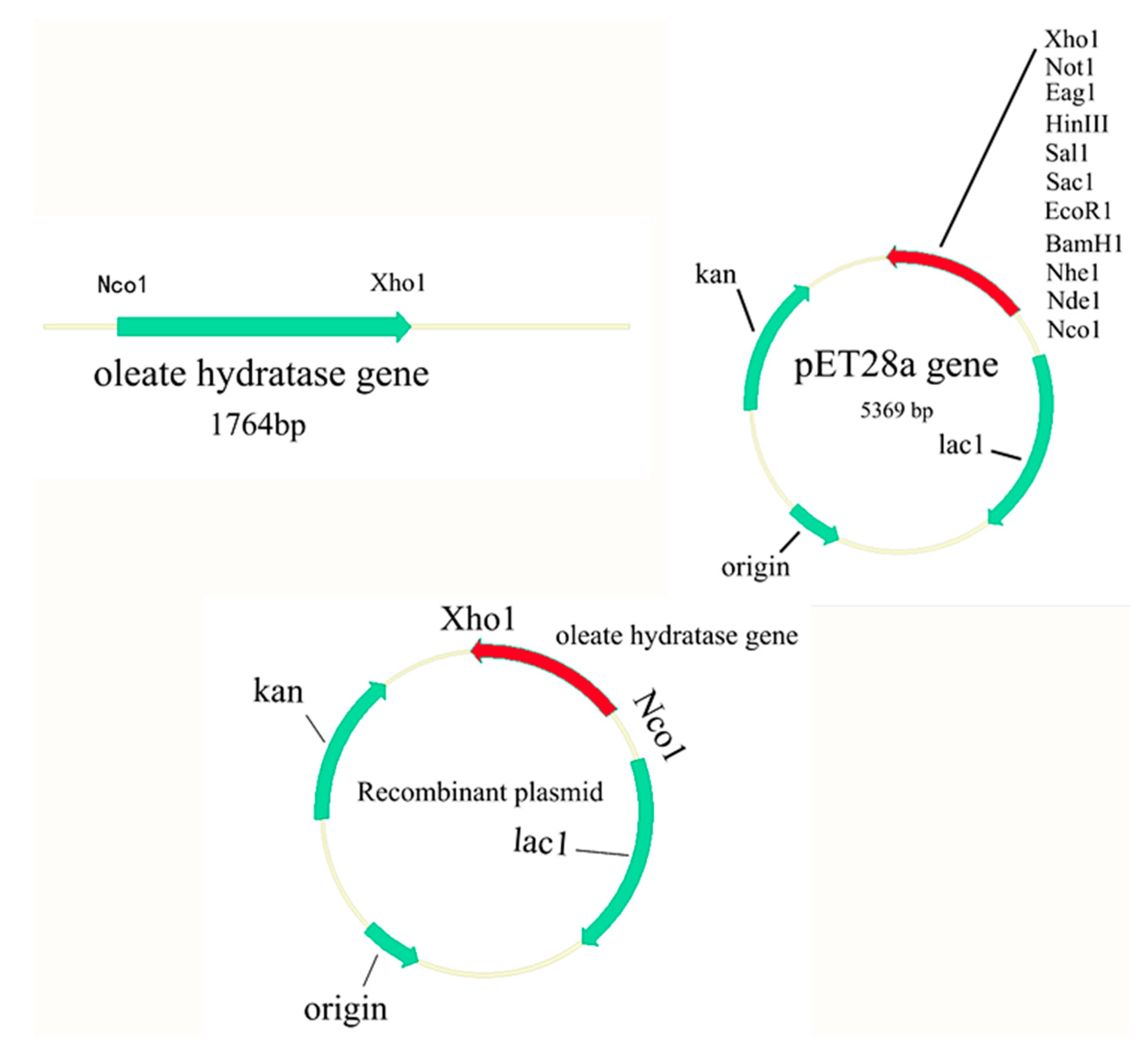{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Vectors, Growth Conditions and Reagents
2.2. Gene Cloning
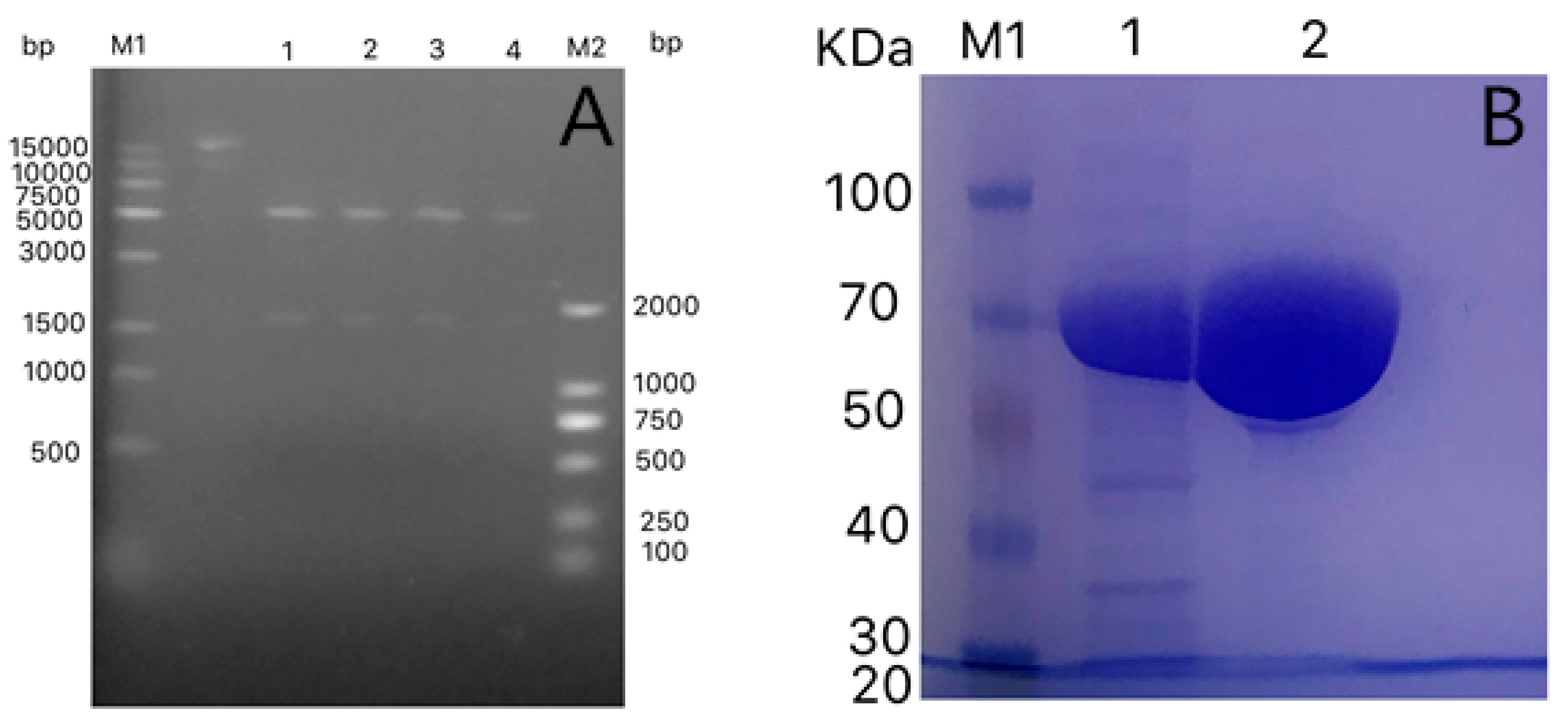
2.3. Enzyme Purification
2.4. SDS-PAGE
2.5. Optimization of Reaction Conditions for 10-HSA Production
2.6. Analytical Methods
2.7. Statistical Analysis
3. Results and Discussion
3.1. Cloning and Molecular Characterization of Fatty Acid Hydratase from Lactococcus garvieae
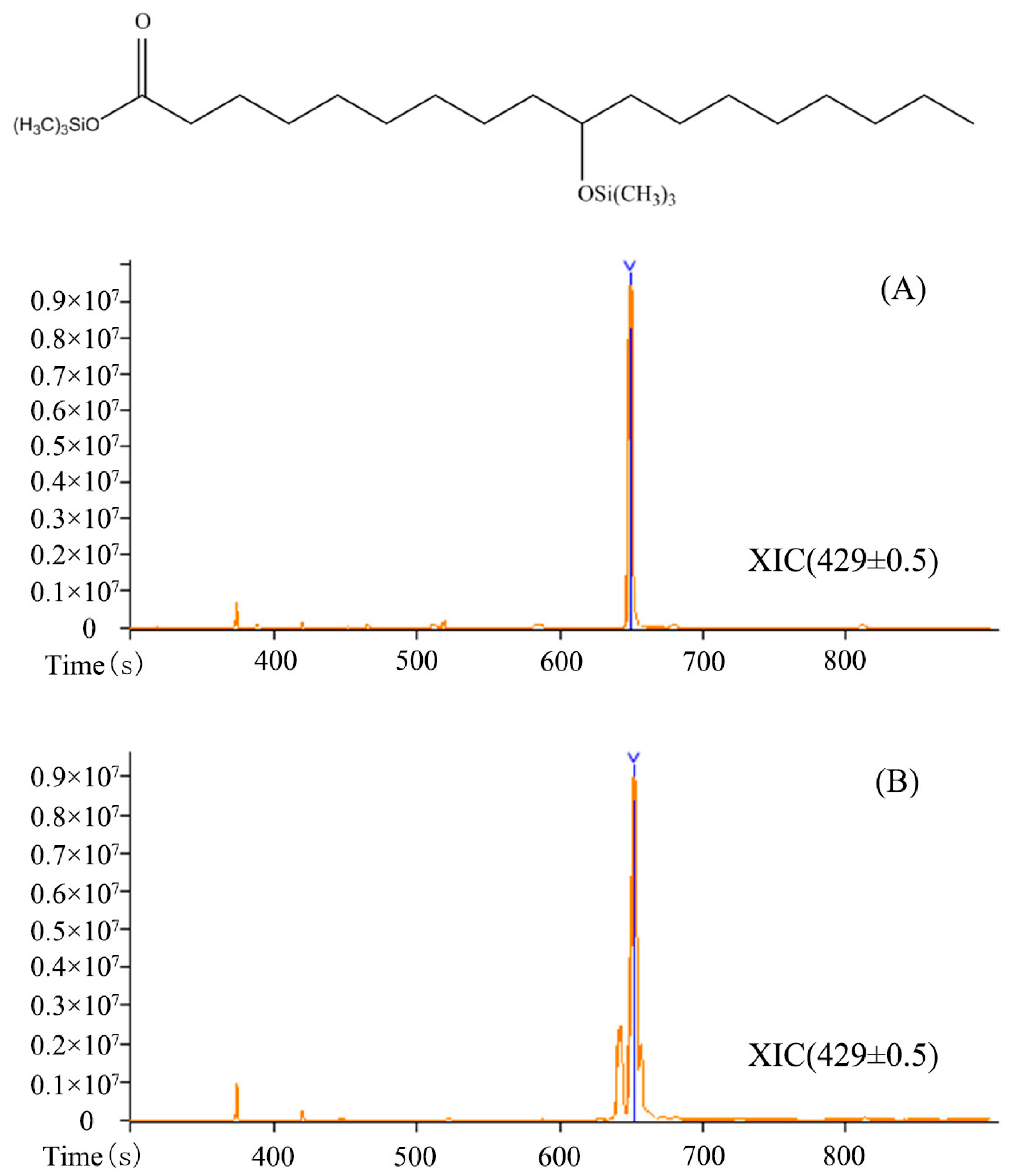
3.2. Identification of Products by GC-MS Analysis
3.3. Optimization of Reaction Conditions
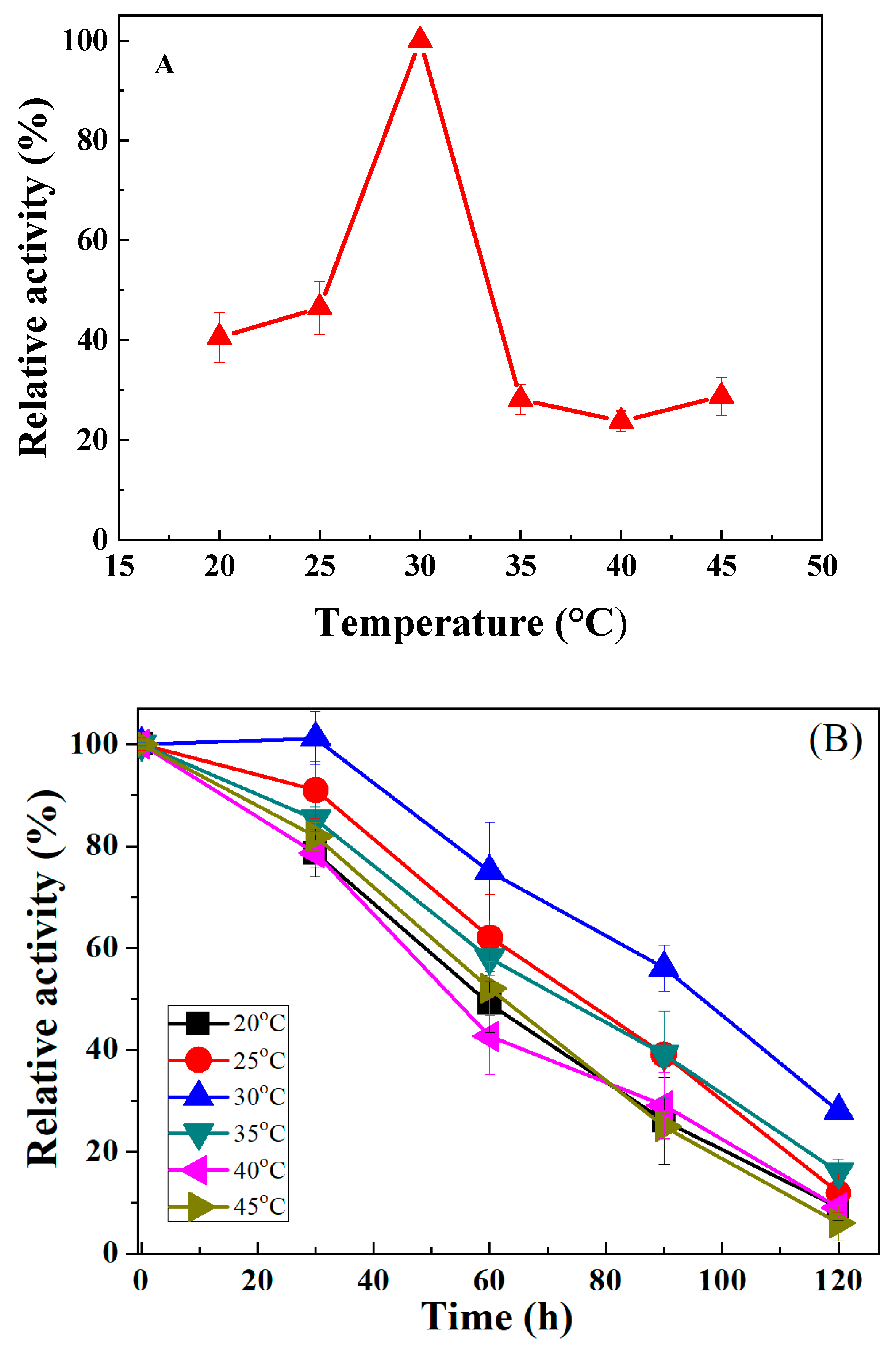
3.3.1. Influence of Temperature
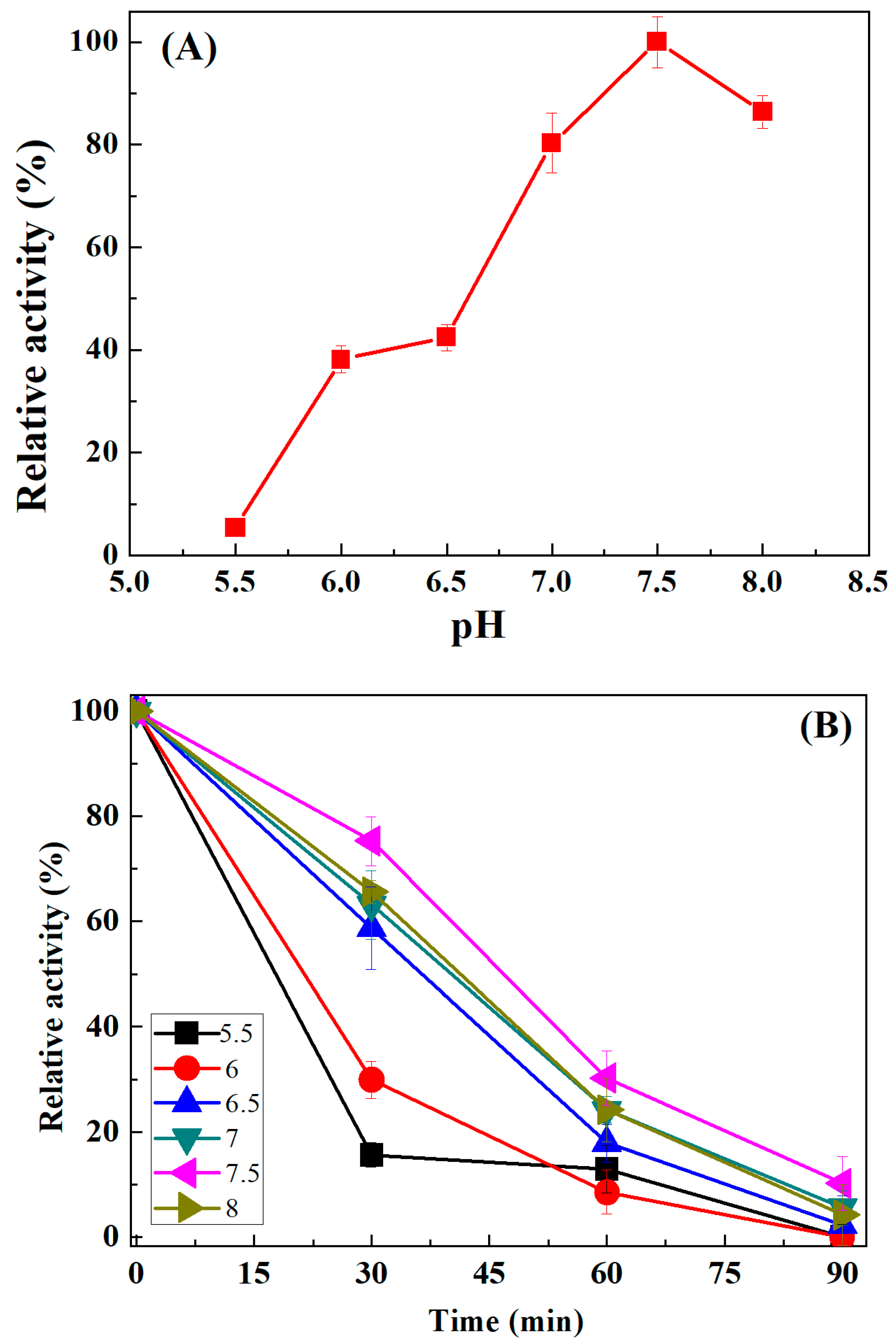
3.3.2. Influence of pH
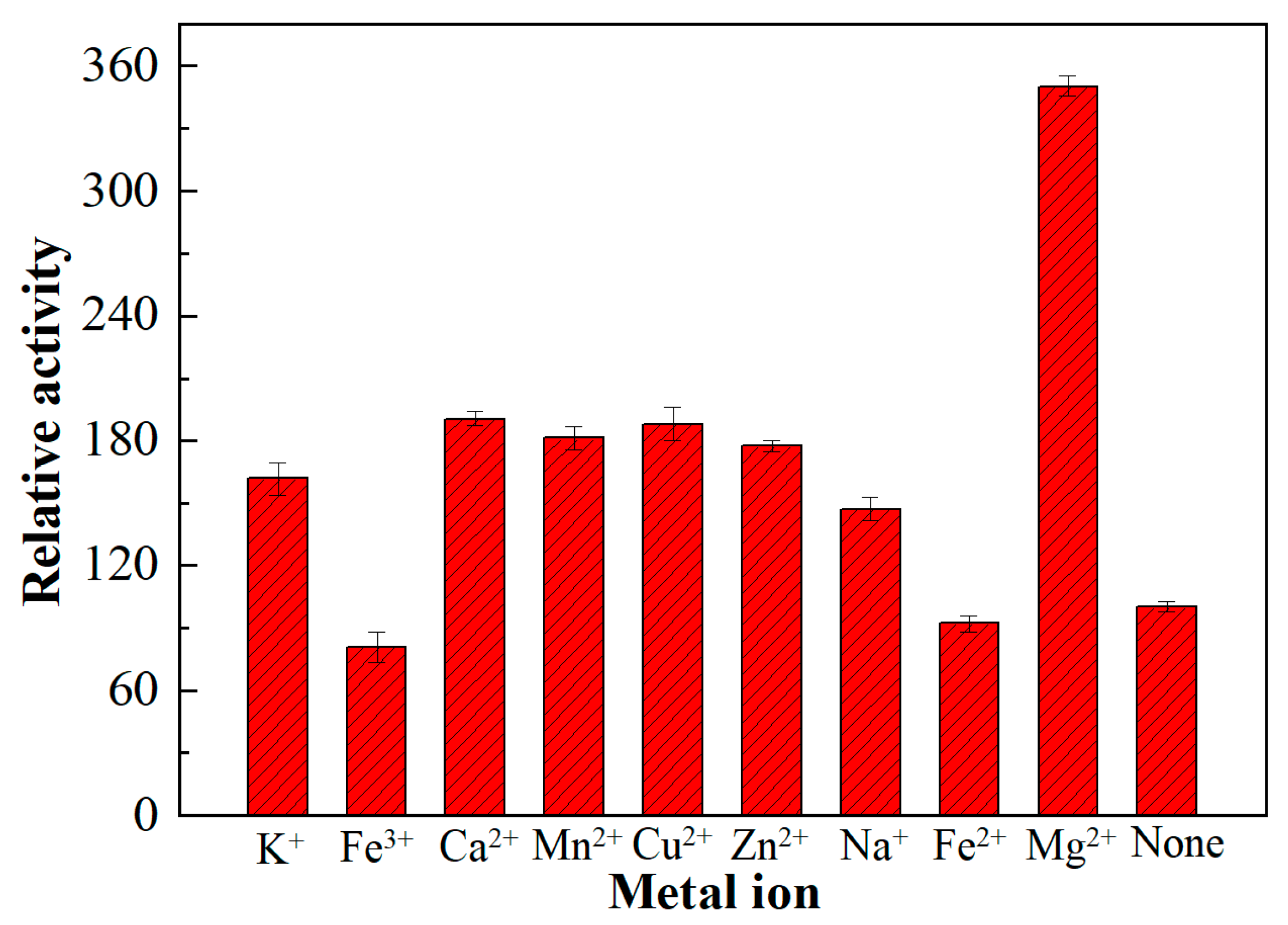
3.3.3. Influence of Metal Ions
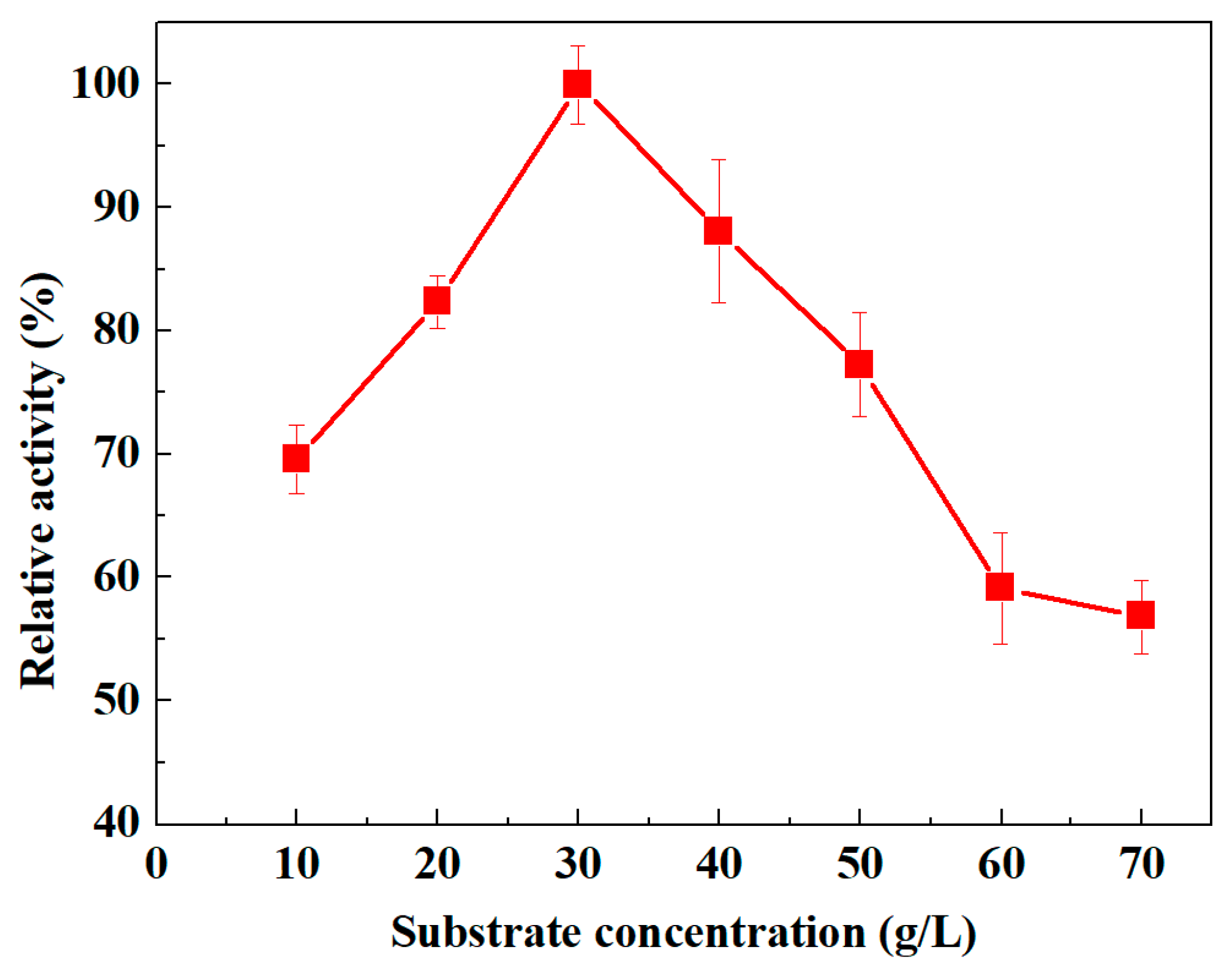
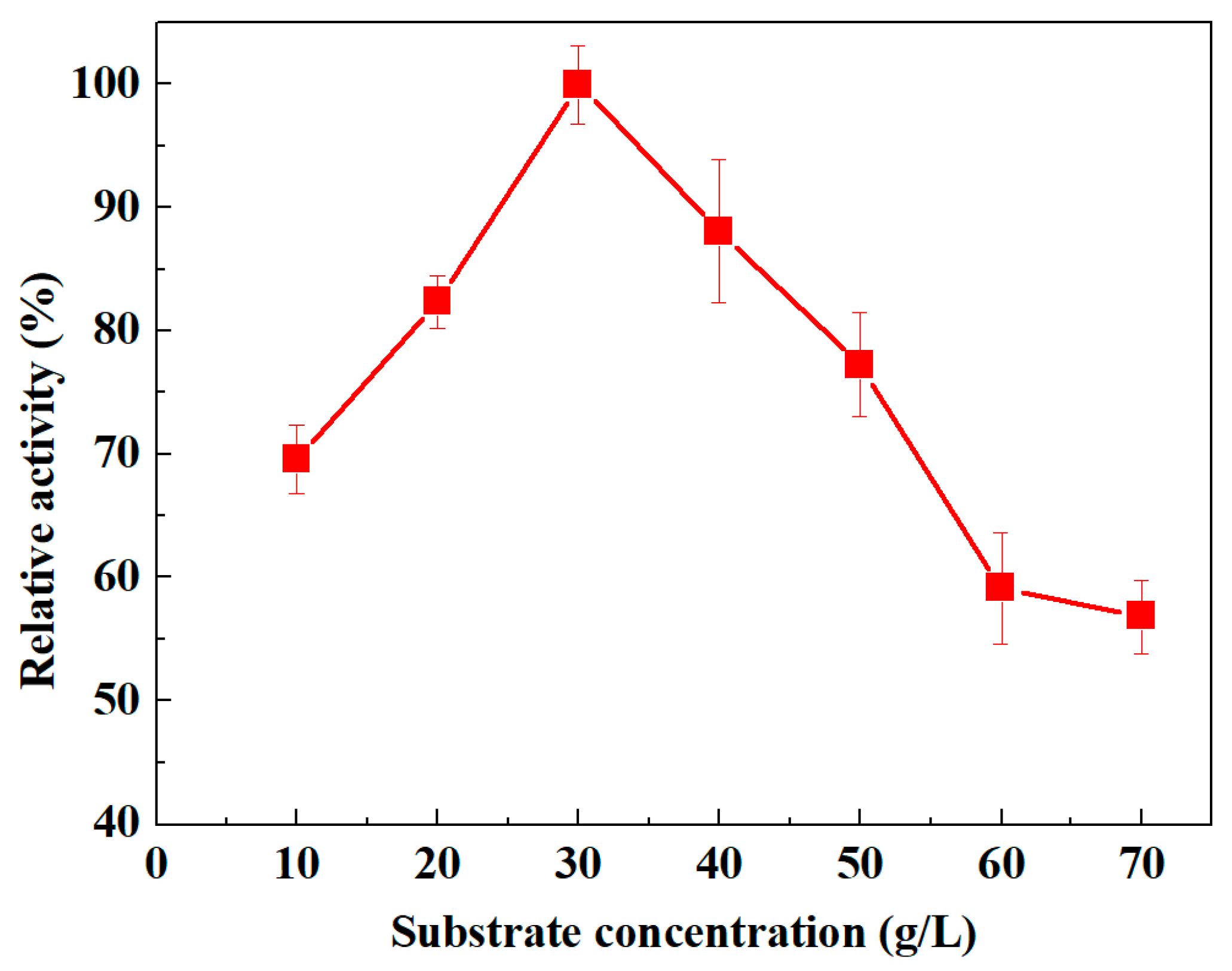
3.3.4. Influence of Substrate Concentration
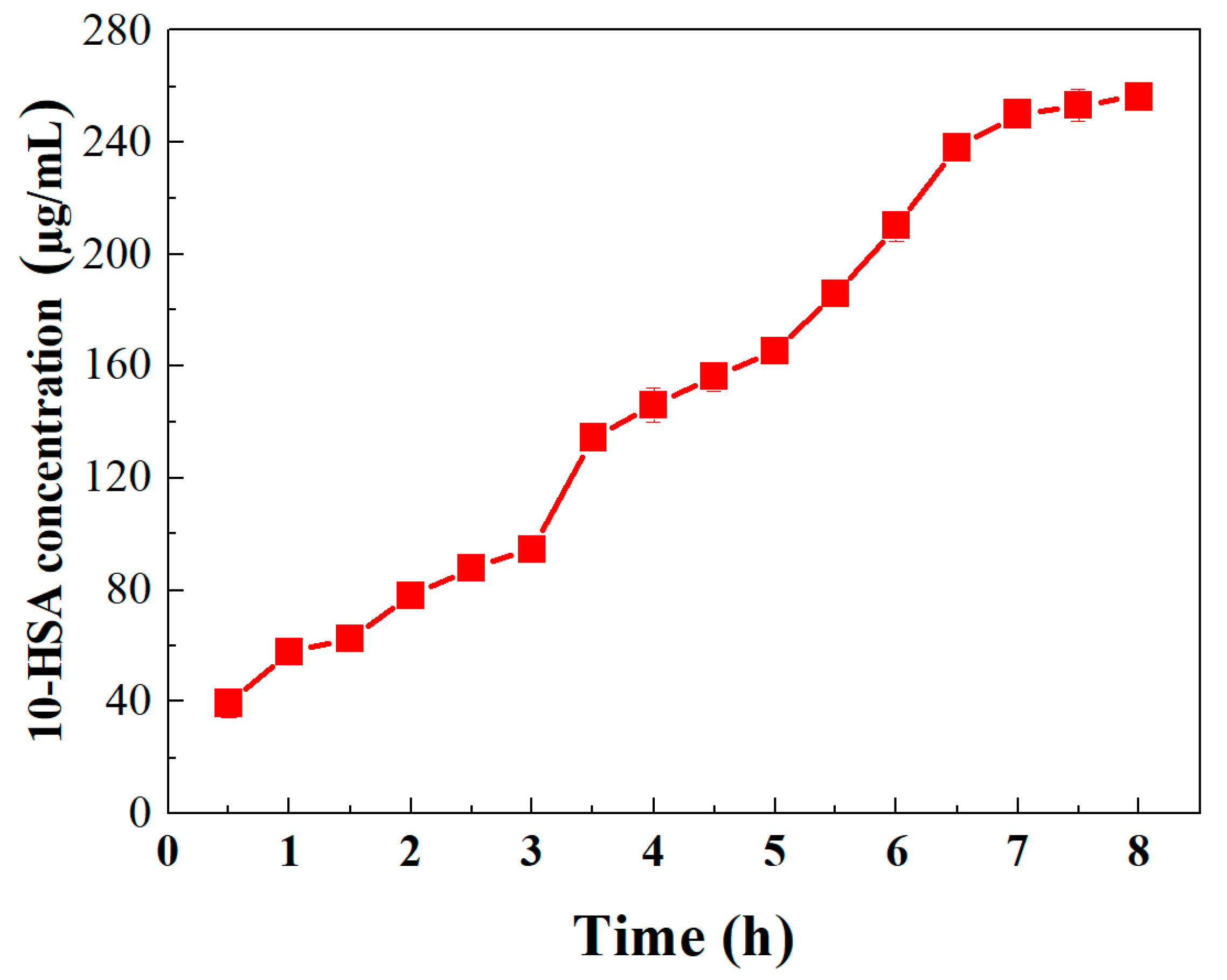
3.3.5. Influence of Reaction Time on Oleic Acid Biotransformation to 10-HSA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, B.S.; Otten, L.G.; Hanefeld, U. Stereochemistry of enzymatic water addition to c=c bonds. Biotechnol. Adv. 2015, 33, 526–546. [Google Scholar] [CrossRef] [PubMed]
- Park, A.K.; Lee, G.H.; Kim, D.W.; Jang, E.H.; Kwon, H.T.; Chi, Y.M. Crystal structure of oleate hydratase from, Stenotrophomonas, sp. kctc 12332 reveals conformational plasticity surrounding the fad-binding site. Biochem. Biophys. Res. Commun. 2018, 499, 772–776. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Anaya, J.; Hernández-Santoyo, A. Functional characterization of a fatty acid double-bond hydratase from lactobacillus plantarum and its interaction with biosynthetic membranes. Biochim. Biophys. Acta-Biomembr. 2015, 1848, 3166–3174. [Google Scholar] [CrossRef] [PubMed]
- Chance, D.L.; Gerhardt, K.O.; Mawhinney, T.P. Gas-liquid chromatography-mass spectrometry of hydroxy fatty acids as their methyl esters tert.-butyldimethylsilyl ethers. J. Chromatogr. A 1998, 793, 91–98. [Google Scholar] [CrossRef]
- Naughton, F.C. Production, chemistry and commercial applica-tions of various chemicals from castor oil. J. Am. Oil Chem. Soc. 1974, 51, 65–71. [Google Scholar] [CrossRef]
- Kim, K.R.; Oh, D.K. Production of hydroxy fatty acids by microbial fatty acid-hydroxylation enzymes. Biotechnol. Adv. 2013, 31, 1473–1485. [Google Scholar] [CrossRef] [PubMed]
- Latrasse, A.; Paitier, S.; Lachot, B.; Bonnarme, P.; Feron, G.; Durand, A.; Le, Q.J. Conversion of oleic acid to 10-hydroxystearic acid by Nocardia paraffinae. Biotechnol. Lett. 1997, 19, 715–718. [Google Scholar] [CrossRef]
- Koritala, S.; Hou, C.; Hesseltine, C.; Bagby, M. Microbial conversion of oleic acid to 10-hydroxystearic acid. Appl. Microbiol. Biotechnol. 1989, 32, 299–304. [Google Scholar] [CrossRef]
- Hudson, J.; MacKenzie, C.; Joblin, K. Conversion of oleic acid to 10-hydroxystearic acid by two species of ruminal bacteria. Appl. Microbiol. Biotechnol. 1995, 44, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.J.; Kim, S.U.; Song, J.W.; Lee, J.H.; Kang, W.R.; Jo, Y.S. Biotransformation of linoleic acid into hydroxy fatty acids and carboxylic acids using a linoleate double bond hydratase as key enzyme. Adv. Synth. Catal. 2015, 357, 408–416. [Google Scholar] [CrossRef]
- Kim, B.N.; Yeom, S.J.; Oh, D.K. Conversion of oleic acid to 10-hydroxystearic acid by whole cells of Stenotrophomonas nitritireducens. Biotechnol. Lett. 2011, 33, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Kitazume, T.; Yamazaki, Y.; Matsuyama, S.; Shoun, H.; Takaya, N. Production of hydroxy-fatty acid derivatives from waste oil by Escherichia coli cells producing fungal cytochrome P450foxy. Appl. Microbiol. Biotechnol. 2008, 79, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.B.; Zhang, Z.; Doherty, W.O.; O’Hara, I.M. A multi-criteria analysis approach for ranking and selection of microorganisms for the production of oils for biodiesel production. Bioresour. Technol. 2015, 190, 264–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.R.; An, J.U.; Lee, S.H.; Oh, D.K. Selective production of 9R-hydroxy-10E,12Z,15Z-octadecatrienoic acid from alpha-linolenic acid in perilla seed oilhydrolyzate by a lipoxygenase from Nostoc Sp. SAG 25.82. PLoS ONE 2015, 10, e0137785. [Google Scholar]
- Fadel, M. Production physiology of cellulases and β-glucosidase enzymes of Aspergillus niger grown under solid state fermentation conditions. Online J. Biol. Sci. 2000, 1, 401–411. [Google Scholar]
- Takeuchi, M.; Kishino, S.; Hirata, A.; Park, S.B.; Kitamura, N.; Ogawa, J. Characterization of the linoleic acid Δ9 hydratase catalyzing the first step of polyunsaturated fatty acid saturation metabolism in Lactobacillus plantarum AKU 1009a. J. Biosci. Bioeng. 2015, 119, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Meng, H.; Jiang, L.; Wang, S. Fatty-Acid–Metal-Ion Complexes as Multicolor Superhydrophobic Coating Materials. Chem. Asian J. 2011, 6, 1757–1760. [Google Scholar] [CrossRef] [PubMed]
- Joo, Y.C.; Jeong, K.W.; Yeom, S.J.; Kim, Y.S.; Kim, Y.; Oh, D.K. Biochemical characterization and FAD-binding analysis of oleate hydratase from Macrococcus caseolyticus. Biochimie 2012, 94, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Bhagavan, N.V.; Ha, C.E. Enzymes and Enzyme Regulation. In Essentials of Medical Biochemistry; Academic Press: Cambridge, MA, USA, 2011; pp. 47–58. [Google Scholar]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Bilal, M.; Liu, S.; Zhang, J.; Lu, H.; Luo, H.; Luo, C.; Shi, H.; Iqbal, H.M.N.; Zhao, Y. Sustainable Biotransformation of Oleic Acid to 10-Hydroxystearic Acid by a Recombinant Oleate Hydratase from Lactococcus garvieae. Processes 2019, 7, 326. https://doi.org/10.3390/pr7060326
Zhang J, Bilal M, Liu S, Zhang J, Lu H, Luo H, Luo C, Shi H, Iqbal HMN, Zhao Y. Sustainable Biotransformation of Oleic Acid to 10-Hydroxystearic Acid by a Recombinant Oleate Hydratase from Lactococcus garvieae. Processes. 2019; 7(6):326. https://doi.org/10.3390/pr7060326
Chicago/Turabian StyleZhang, Jing, Muhammad Bilal, Shuai Liu, Jiaheng Zhang, Hedong Lu, Hongzhen Luo, Chuping Luo, Hao Shi, Hafiz M. N. Iqbal, and Yuping Zhao. 2019. "Sustainable Biotransformation of Oleic Acid to 10-Hydroxystearic Acid by a Recombinant Oleate Hydratase from Lactococcus garvieae" Processes 7, no. 6: 326. https://doi.org/10.3390/pr7060326
APA StyleZhang, J., Bilal, M., Liu, S., Zhang, J., Lu, H., Luo, H., Luo, C., Shi, H., Iqbal, H. M. N., & Zhao, Y. (2019). Sustainable Biotransformation of Oleic Acid to 10-Hydroxystearic Acid by a Recombinant Oleate Hydratase from Lactococcus garvieae. Processes, 7(6), 326. https://doi.org/10.3390/pr7060326