Maximization of Siderophores Production from Biocontrol Agents, Pseudomonas aeruginosa F2 and Pseudomonas fluorescens JY3 Using Batch and Exponential Fed-Batch Fermentation

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial and Fungal Isolates
2.1.1. Isolation of Fluorescent Pseudomonas Isolates
2.1.2. Bacterial and Fungal Isolates from Culture Collection
2.2. Screening for Siderophores Production Using CAS Assay
2.2.1. Qualitative Assay
2.2.2. Quantitative Assay
2.3. Antagonistic Effect of Fluorescent Pseudomonad Isolates (Dual Culture Method)
2.4. Molecular Identification of Bacterial Isolates
2.5. Fermentation Experiments
2.5.1. Bioreactor
2.5.2. Batch Fermentation
2.5.3. Fed-Batch Fermentation
2.6. Analytical Procedures
2.6.1. Relative Level of Siderophores
2.6.2. Biomass Estimation
2.6.3. Glucose Estimation
2.6.4. Glycerol Estimation
2.7. Application of Siderophores-Producing Pseudomonas Isolates as Biocontrol Agents
2.7.1. Formulation Experiment
2.7.2. Fungal Inoculum Preparation
2.7.3. Soil Infestation
2.7.4. Disease Assessment
2.7.5. Greenhouse Experiment
2.8. Statistical Analysis
3. Results
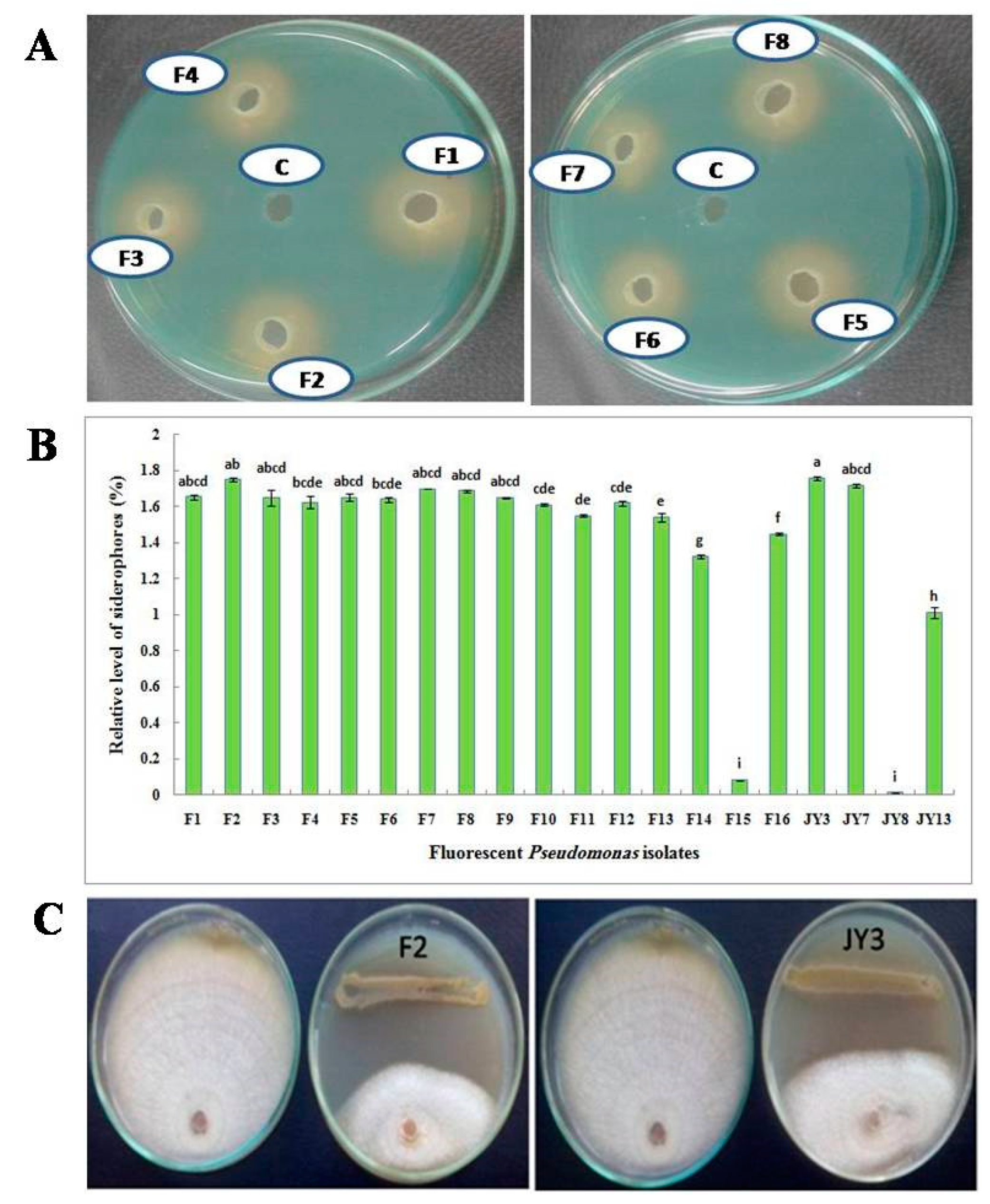
3.1. Isolation of Fluorescent Pseudomonas Isolates
3.2. Screening for Siderophores Production
3.3. Antagonistic Effect of Fluorescent Pseudomonad Isolates (Dual Culture Method)
3.4. Molecular Identification of Bacterial Isolates
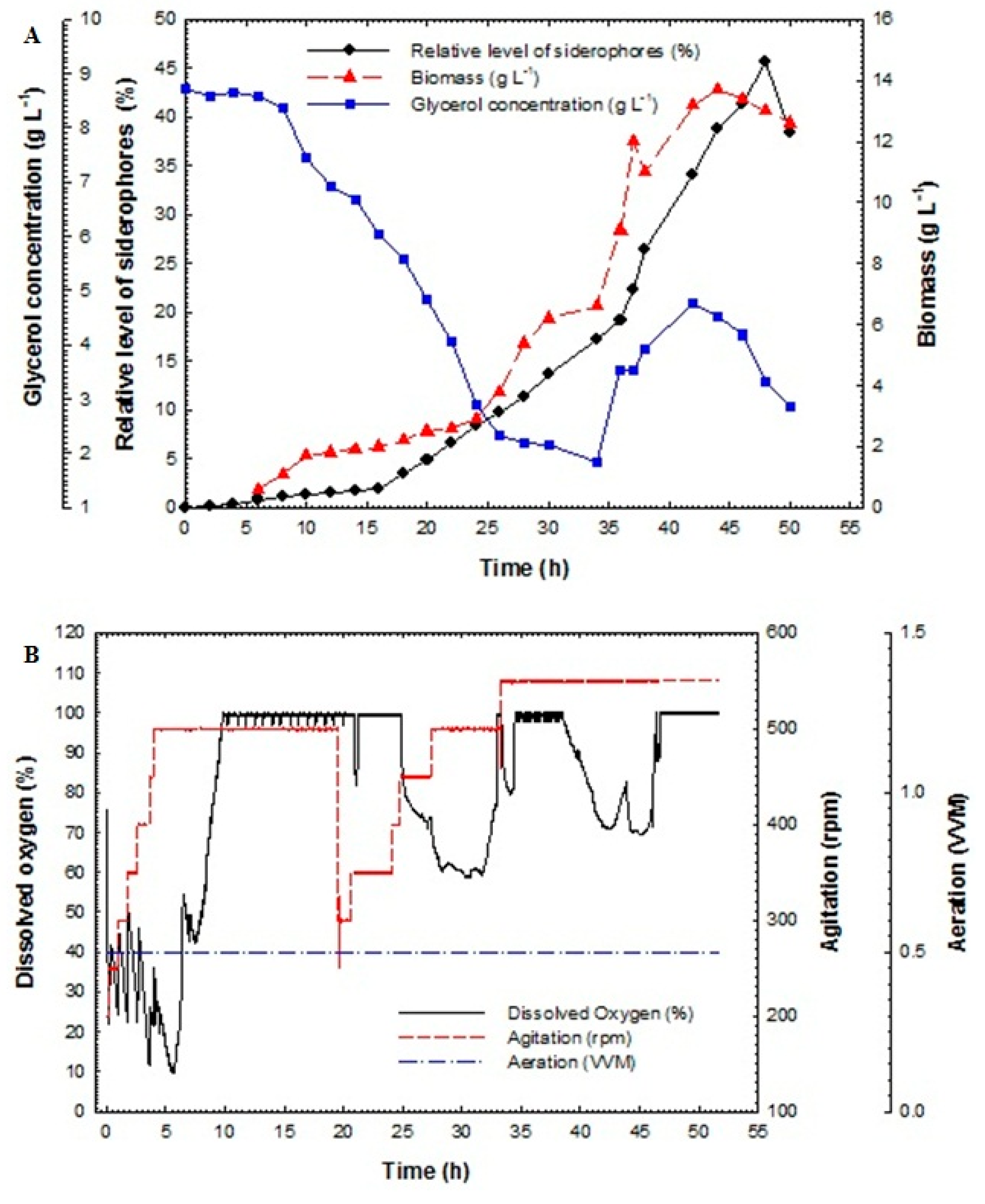
3.5. Batch Fermentation
3.6. Fed-Batch Fermentation
3.7. In-Vivo Antagonistic Effect of P. aeruginosa F2 and P. fluorescens JY3 against Some Phytopathogenic Fungi
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Neilands, J.B. Microbial iron compounds. Annu. Rev. Biochem. 1981, 50, 715–731. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Kurth, C.; Kage, H.; Nett, M. Siderophores as molecular tools in medical and environmental applications. Org. Biomol. Chem. 2016, 14, 8212–8227. [Google Scholar] [CrossRef] [PubMed]
- Kloepper, J.W.; Leong, J.; Teintze, M.; Schroth, M.N. Enhancing plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature 1980, 286, 885–886. [Google Scholar] [CrossRef]
- Arya, N.; Rana, A.; Rajwar, A.; Sahgal, M.; Sharma, A.K. Biocontrol efficacy of siderophore producing indigenous pseudomonas strains against fusarium wilt in tomato. Natl. Acad. Sci. Lett. 2018, 41, 133–136. [Google Scholar] [CrossRef]
- Gupta, C.P.; Dubey, R.C.; Maheshwari, D.K. Plant growth enhancement and suppression of Macrophomina phaseolina causing charcoal rot of peanut by fluorescent Pseudomonas. Biol. Fertil. Soils 2002, 35, 399–405. [Google Scholar]
- Yu, S.; Teng, C.; Liang, J.; Song, T.; Dong, L.; Bai, X.; Jin, Y.; Qu, J. Characterization of siderophore produced by Pseudomonas syringae BAF.1 and its inhibitory effects on spore germination and mycelium morphology of Fusarium oxysporum. J. Microbiol. 2017, 55, 877–884. [Google Scholar] [CrossRef]
- Sayyed, R.Z.; Badgujar, M.D.; Sonawane, H.M.; Mhaske, M.M.; Chincholkar, S.B. Production of microbial iron chelators (Siderophores) by fluorescent Pseudomonads. Ind. J. Biotechnol. 2005, 4, 484–490. [Google Scholar]
- Sarma, M.; Saharan, K.; Kumar, L.; Gautam, A.; Kapoor, A.; Srivastava, N.; Sahai, V.; Bisaria, V. Process optimization for enhanced production of cell biomass and metabolites of fluorescent pseudomonad r81. Int. J. Biomed. Biol. Eng. 2010, 4, 388–392. [Google Scholar]
- Shaikh, S.; Wani, S.; Sayyed, R. Statistical-based optimization and scale-up of siderophore production process on laboratory bioreactor. 3 Biotech 2016, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Paulová, L.; Patáková, P.; Brányik, T. Advanced fermentation processes. In Engineering Aspects of Food Biotechnology; Teixeira, J., Vincente, A.A., Eds.; Taylor & Francis Group: Milton Park, UK, 2016; pp. 90–99. [Google Scholar]
- Abo-Zaid, G.A.; Wagih, E.E.; Matar, S.M.; Ashmawy, N.A.; Hafez, E.E. Scaling-up production of pyocyanin from Pseudomonas aeruginosa JY21 as biocontrol agent against certain plant pathogenic fungi. Int. J. ChemTech Res. 2015, 8, 213–224. [Google Scholar]
- Shuler, M.L.; Kargi, F. Bioprocess Engineering Basic Concepts, 2nd ed.; Prentice-Hall PTR: Upper Saddle River, NJ, USA, 2002. [Google Scholar]
- Matar, S.M.; El-Kazzaz, S.A.; Wagih, E.E.; El-Diwany, A.I.; Moustafa, H.E.; El-Saadani, M.A.; Abo-Zaid, G.A.; Hafez, E.E. Bioprocessing and scaling-up cultivation of Bacillus subtilis as a potential antagonist to certain plant pathogenic fungi, III. Biotechnology 2009, 8, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Ridgway, D.; Moo-Young, T.G.M. Enhanced amylase production by Bacillus subtilis using a dual exponential feeding strategy. Bioprocess Biosyst. Eng. 2004, 27, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Luli, G.W. The Influence of Glucose Metabolism on High Cell Density Fermentation. Ph.D. Thesis, Ohio State University, Columbus, OH, USA, 1988. [Google Scholar]
- Lee, J.H.; Hong, J.; Lim, H.C. Experimental optimization of fed-batch culture for poly-β-hydroxybutyric acid production. Biotechnol. Bioeng. 1997, 56, 697–705. [Google Scholar] [CrossRef]
- Salwa, M.S.; Asshifa, M.N.N.; Amirul, A.A.; Yahya, A.R.M. Different feeding strategy for the production of biosurfactant from Pseudomonas aeruginosa USM AR2 in modified bioreactor. Biotechnol. Bioprocess Eng. 2009, 14, 763–768. [Google Scholar] [CrossRef]
- Lotfabad, T.B.; Tayyebi, S.; Roostaazad, R. Kinetic Measurements for Pseudomonas aeruginosa MR01 during biosurfactant production in two-phase system and developing a double-exponential model for viable cell profile. World Appl. Sci. J. 2013, 22, 809–816. [Google Scholar]
- Ju, X.; Yu, H.L.; Pan, J.; Xu, J.H. Improved production of Pseudomonas sp. ECU1011 acetyl esterase by medium design and fed-batch fermentation. Bioprocess Biosyst. Eng. 2012, 35, 323–331. [Google Scholar] [CrossRef]
- Radha, S.; Prasad, T.H.; Devi, N.S.; Mamata, S. Fed batch and batch submerged fermentation for alkaline protease production from mixed consortium of Pseudomonas putida and Staphylococcus aureus. Adv. Appl. Sci. Res. 2014, 5, 315–324. [Google Scholar]
- Buyer, J.S.; Leong, J. Iron transport-mediated antagonism between plant growth-promoting and plant-deleterious Pseudomonas strains. J. Biol. Chem. 1986, 261, 791–794. [Google Scholar]
- Kenawy, A.; Dailin, D.J.; Abo-Zaid, G.A.; Abd Malek, R.; Ambehabati, K.K.; Zakaria, K.H.; Sayyed, R.Z.; Enshasy, H.A. Biosynthesis of antibiotics by PGPR and their roles in biocontrol of plant diseases. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Sayyed, R.Z., Ed.; Springer: Singapore, 2019; pp. 1–36. [Google Scholar]
- De Boer, M.; Bom, P.; Kindt, F.; Keurentjes, J.J.B.; van der Sluis, I.; Van Loon, L.C.; Bakker, P.A. Control of Fusarium wilt of radish by combining Pseudomonas putida strains that have different disease-suppressive mechanisms. Phytopathology 2003, 93, 626–632. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, W. Mechanisms and regulation of reduction-based iron uptake in plants. New Phytol. 1999, 141, 1–26. [Google Scholar] [CrossRef]
- Vansuyt, G.; Robin, A.; Briat, J.F.; Curie, C.; Lemanceau, P. Iron acquisition from Fe-pyoverdine by Arabidopsis thaliana. Mol. Plant Microbe Interact. 2007, 20, 441–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schywn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Meyer, J.M.; Abdallah, M.A. The fluorescent pigment of Pseudomonas fluorescens: Biosynthesis, purification and physicochemical properties. J. Gen. Microbiol. 1978, 107, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Toure, Y.; Ongena, M.; Jacques, P.; Guiro, A.; Thonart, P. Role of lipopeptides produced by Bacillus subtilis GA1 in the reduction of grey mould disease caused by Botrytis cinerea on apple. J. Appl. Microbiol. 2004, 96, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- Istock, C.A.; Ferguson, N.; Istock, N.L.; Duncan, K.E. Geographical diversity of genomic lineages in Bacillus subtilis (Ehrenberg) Cohn sensulato. Org. Divers. Evol. 2001, 1, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Matar, S.M.; El-Kazzaz, S.A.; Wagih, E.E.; El-Diwany, A.I.; Hafez, E.E.; Moustafa, H.E.; Abo-Zaid, G.A.; Serour, E.A. Molecular characterization and batch fermentation of Bacillus subtilis as biocontrol agent II. Biotechnology 2009, 8, 35–43. [Google Scholar] [CrossRef]
- Abo-Zaid, G.A.; Abdullah, A.S.; Soliman, N.A.; El-Sharouny, E.E.; Sabry, S.A. Optimization of siderophores production from fluorescent pseudomonads using statistical experimental designs. Biosci. Res. 2018, 15, 3040–3051. [Google Scholar]
- Van Dam-Mieras, M.C.E.; Jeu, W.H.; Vries, J.; Currell, B.R.; James, J.W.; Leach, C.K.; Patmore, R.A. Techniques Used in Bioproduct Analysis; Butterworth-Heinemann Ltd.: Oxford, UK, 1992. [Google Scholar]
- Bok, S.H.; Demain, A.L. An Improved Calorimetric Assay for Polyols. Anal. Biochem. 1977, 81, 18–20. [Google Scholar] [CrossRef]
- Vidhyasekaran, P.; Muthamilan, M. Development of formulation of Pseudomonas fluorescens for control of chickpea wilt. Plant Dis. 1995, 79, 782–786. [Google Scholar] [CrossRef]
- Hussien, Z.N.; Mahmoud, E.Y.; Metwaly, A.H.; Sobhy, H.M. Effect of some antagonistic bacteria in reducing of peanut damping-off, root and pod rot incidence caused by Rhizoctonia solani. J. Plant Prot. Pathol. 2012, 3, 1173–1187. [Google Scholar]
- Omidvari, M.; Sharifi, R.A.; Ahmadzadeh, M.; Dahaji, P.A. Role of fluorescent pseudomonads siderophore to increase bean growth factors. J. Agric. Sci. 2010, 2, 242–247. [Google Scholar] [CrossRef] [Green Version]
- Sulochana, M.B.; Jayachandra, S.Y.; Kumar, S.K.A.; Dayanand, A. Antifungal attributes of siderophore produced by the Pseudomonas aeruginosa JAS-25. J. Basic Microbiol. 2014, 54, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Páez, M.; Martínez-Nieto, P.; Bernal-Castillo, J. Siderophores producing Pseudomonas as pathogenic Rhizoctonia solani and Botrytis cinerea antagonistis. Univ. Sci. 2005, 10, 65–74. [Google Scholar]
- Sasirekha, B.; Srividya, S. Siderophore production by Pseudomonas aeruginosa FP6, a biocontrol strain for Rhizoctonia solani and Colletotrichum gloeosporioides causing diseases in chilli. Agric. Nat. Res. 2016, 50, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.A.; Nain, Z.; Alam, M.K.; Banu, N.A.; Islam, M.R. In vitro study of biocontrol potential of rhizospheric Pseudomonas aeruginosa against Fusarium oxysporum f. sp. cucumerinum. Egypt. J. Biol. Pest Control 2018, 28, 90. [Google Scholar] [CrossRef]
- Ahmadzadeh, M.; Afsharmanesh, H.; Nikkhah, M.J.; Tehrani, A.S. Identification of some molecular traits in fluorescent pseudomonads with antifungal activity. Iran. J. Biotechnol. 2006, 4, 245–253. [Google Scholar]
- Sabra, W.; Kim, E.J.; Zeng, A.P. Physiological responses of Pseudomonas aeruginosa PAO1 to oxidative stress in controlled microaerobic and aerobic cultures. Microbiology 2002, 148, 3195–3202. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; Kai, K.; Lijima, S.; Kobayashi, T. Enhanced β-galactosidase production by high cell-density culture of recombinant Bacillus subtilis with glucose concentration control. Biotechnol. Bioeng. 1992, 40, 686–696. [Google Scholar] [CrossRef]
- Yee, L.; Blanch, H.W. Recombinant trypsin production in high cell density fed-batch cultures in Escherichia coli. Biotechnol. Bioeng. 1993, 41, 781–790. [Google Scholar] [CrossRef]
- Lee, S.Y. High cell-density culture of Escherichia coli. Trends Biotechnol. 1996, 14, 98–105. [Google Scholar] [CrossRef]
- Chosewood, L.C.; Wilson, D.E. Biosafety in Microbiological and Biomedical Laboratories, 5th ed.; HHS Publication: Washington, DC, USA, 2009; pp. 9–19. [Google Scholar]
- OECD. Bacteria: Pathogenicity factors. In Safety Assessment of Transgenic Organisms in the Environment; OECD Consensus Documents; OECD Publishing: Paris, France, 2016; Volume 5, pp. 27–79. [Google Scholar]
- Blacksell, S.D.; Robinson, M.T.; Newton, P.N.; Ruanchaimun, S.; Salje, J.; Wangrangsimakul, T.; Wegner, M.D.; Abdad, M.Y.; Bennett, A.M.; Richards, A.L.; et al. Biosafety and biosecurity requirements for Orientia spp. diagnosis and research: Recommendations for risk-based biocontainment, work practices and the case for reclassification to risk group 2. BMC Infect. Dis. 2019, 19, 1044. [Google Scholar] [CrossRef] [PubMed]
- Solans, M.; Scervino, J.M.; Messuti, M.I.; Vobis, G.; Wall, L.G. Potential biocontrol actinobacteria: Rhizospheric isolates from the argentine pampas lowlands legumes. J. Basic Microbiol. 2016, 56, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Leeman, M.; Den Ouden, F.M.; Van Pelt, J.A.; Dirkx, F.P.M.; Steijl, H.; Bakker, P.A. Iron availability affects induction of systemic resistance to Fusarium wilt of radish by Pseudomonas fluorescens. Phytopathology 1996, 86, 149–155. [Google Scholar] [CrossRef]
- Sharma, A.; Johri, B.N.; Sharma, A.K.; Glick, B.R. Plant growth-promoting bacterium Pseudomonassp. strain GRP3 influences iron acquisition in mung bean (Vigna radiataL. Wilzeck). Soil Biol. Biochem. 2003, 35, 887–894. [Google Scholar] [CrossRef]
- Manwar, A.V.; Vaigankar, P.D.; Bhonge, L.S.; Chincholker, S.B. In vitro suppression of plant pathogens by siderophores of fluorescent Pseudomonads. Ind. J. Microbiol. 2000, 40, 109–112. [Google Scholar]
- Saharan, K.; Sarma, M.V.; Srivastava, R.; Sharma, A.K.; Johri, B.N.; Prakash, A.; Sahai, V.; Bisaria, V.S. Development of non-sterile inorganic carrier-based formulations of fluorescent pseudomonad R62 and R81 and evaluation of their efficacy on agricultural crops. Appl. Soil Ecol. 2010, 46, 251–258. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fluorescent Pseudomonas Isolates Isolated in the Current Study | |||||
| Isolate Code | Plant Rhizosphere | Isolation Governorate | GPS Coordinates | GenBank Accession Number | |
| Latitude | Longitude | ||||
| F1 | Corn | Monufia | 30.5972455 | 30.9876321 | |
| F2 | Eggplant | Monufia | 30.5972455 | 30.9876321 | MG210480 |
| F3 | Pepper | Alexandria | 31.200092 | 29.918739 | |
| F4 | Pepper | Alexandria | 31.200092 | 29.918739 | |
| F5 | Eggplant | Alexandria | 31.200092 | 29.918739 | |
| F6 | Eggplant | Alexandria | 31.200092 | 29.918739 | |
| F7 | Cotton | Sohag | 26.549999 | 31.700001 | MG076939 |
| F8 | Corn | Sohag | 26.549999 | 31.700001 | MG210481 |
| F9 | Cotton | Sohag | 26.549999 | 31.700001 | |
| F10 | Corn | Sohag | 26.549999 | 31.700001 | |
| F11 | Corn | Sohag | 26.549999 | 31.700001 | |
| F12 | Corn | Sohag | 26.549999 | 31.700001 | |
| F13 | Wheat | Sohag | 26.549999 | 31.700001 | |
| F14 | Clover | Sohag | 26.549999 | 31.700001 | |
| F15 | Wheat | Sohag | 26.549999 | 31.700001 | |
| F16 | Clover | Sohag | 26.549999 | 31.700001 | |
| Pseudomonas fluorescens Isolates Provided from Culture Collection | |||||
| Isolate Code | GenBank Accession Number | ||||
| JY3 | KF922490 | ||||
| JY7 | KF922494 | ||||
| JY8 | KF922495 | ||||
| JY13 | KF922500 | ||||
| Pseudomonas Isolates | Vertical Distance of Inhibition Diameter (cm) | |||||
|---|---|---|---|---|---|---|
| Alternaria Sp | F. culmorum | F. oxysporum Isolate A | F. oxysporum Isolate B | F. solani | R. solani | |
| F1 | *2.00 ± 0.17 bcd** | 2.13 ± 0.45 abcd | 2.20 ± 0.10 a | 1.70 ± 0.10 a | 2.07 ± 0.21 abc | 1.63 ± 0.15 e |
| F2 | 2.23 ± 0.21 ab | 2.47 ± 0.06 a | 2.10 ± 0.29 abc | 1.70 ± 0.20 a | 2.23 ± 0.30 a | 2.23 ± 0.21 a |
| F3 | 1.37 ± 0.32 e | 1.87 ± 021 de | 1.60 ± 0.10 fg | 1.43 ± 0.06 abc | 1.37 ± 0.32 gh | 1.77 ± 0.21 de |
| F4 | 1.80 ± 0.26 d | 1.90 ± 0.26 de | 1.80 ± 0.20 def | 1.60 ± 0.10 ab | 1.80 ± 0.26 cde | 2.10 ± 0.36 abc |
| F5 | 2.27 ± 0.25 a | 2.10 ± 0.17 bcd | 1.80 ± 0.10 def | 1.60 ± 0.00 ab | 2.13 ± 0.25 ab | 2.10 ± 0.15 abc |
| F6 | 1.43 ± 0.12 e | 1.80 ± 0.10 de | 1.67 ± 0.15 efg | 1.53 ± 0.15 abc | 1.47 ± 0.15 fg | 2.10 ± 0.10 abc |
| F7 | 2.13 ± 0.06 abc | 2.27 ± 0.15 abc | 2.17 ± 0.15 ab | 1.57 ± 0.11 abc | 2.13 ± 0.21 ab | 2.10 ± 0.35 abc |
| F8 | 2.00 ± 0.10 bcd | 2.13 ± 0.15 abcd | 1.97 ± 0.12 bcd | 1.57 ± 0.49 abc | 2.00 ± 0.10 abcd | 2.00 ± 0.36 abcd |
| F9 | 2.17 ± 0.25 ab | 2.33 ± 0.25 ab | 1.63 ± 0.12 efg | 0.87 ± 0.32 d | 2.07 ± 0.25 abc | 2.07 ± 0.15 abc |
| F10 | 2.23 ± 0.12 ab | 2.00 ± 0.10 bcde | 1.93 ± 0.15 cd | 1.43 ± 0.15 abc | 2.23 ± 0.12 a | 2.17 ± 0.20 ab |
| F11 | 2.10 ± 0.26 abc | 2.07 ± 0.21 bcd | 1.53 ± 0.15 gh | 1.23 ± 0.06 c | 2.10 ± 0.10 ab | 2.23 ± 0.21 a |
| F12 | 1.90 ± 0.10 cd | 1.97 ± 0.06 cde | 1.27 ± 0.21 ij | 1.27 ± 0.59 bc | 1.90 ± 0.10 bcde | 1.90 ± 0.26 bcde |
| F13 | 2.10 ± 0.26 abc | 1.70 ± 0.36 e | 1.83 ± 0.21 de | 1.50 ± 0.10 abc | 2.10 ± 0.26 ab | 1.90 ± 0.20 bcde |
| F14 | 0.00 ± 0.00 g | 2.10 ± 0.10 bcd | 1.60 ± 0.10 fg | 0.00 ± 0.00 e | 0.00 ± 0.00 j | 1.63 ± 0.15 e |
| F15 | 0.00 ± 0.00 g | 0.00 ± 0.00 h | 1.33 ± 0.15 hi | 0.00 ± 0.00 e | 0.00 ± 0.00 j | 0.00 ± 0.00 h |
| F16 | 0.00 ± 0.00 g | 1.33 ± 0.42 f | 0.00 ± 0.00 k | 0.00 ± 0.00 e | 0.67 ± 0.31 i | 1.83 ± 0.25 cde |
| P. fluorescens JY3 | 0.63 ± 0.15 f | 1.07 ± 0.15 fg | 1.07 ± 0.15 j | 1.23 ± 0.15 c | 1.77 ± 0.38 de | 0.63 ± 0.10 g |
| P. fluorescens JY7 | 0.10 ± 0.00 g | 0.77 ± 0.21 g | 0.00 ± 0.00 k | 0.30 ± 0.10 e | 1.17 ± 0.10 h | 0.00 ± 0.00 h |
| P. fluorescens JY8 | 0.13 ± 0.00 g | 0.00 ± 0.00 h | 0.00 ± 0.00 k | 0.00 ± 0.00 e | 1.40 ± 0.10 fgh | 1.07 ± 0.06 f |
| P. fluorescens JY13 | 0.00 ± 0.00 g | 0.93 ± 0.25 g | 0.00 ± 0.00 k | 0.00 ± 0.00 e | 1.67 ± 0.15 ef | 0.00 ± 0.00 h |
| L.S.D | 0.266 | 0.360 | 0.222 | 0.342 | 0.276 | 0.296 |
| Treatment | Damping Off (%) | Reduction W (%) | Reduction X (%) |
|---|---|---|---|
| 1. F. oxysporum (check) | *33.33 ± 11.55 ab** | 0 | n.d. |
| 2. R. solani (check) | 53.33 ± 23.09 a | n.d. | 0 |
| 3. Control | 20.00 ± 00.00 b | 40 | 62.50 |
| 4. P. aeruginosa F2 | 13.33 ± 11.55 b | 60 | 75 |
| 5. P. fluorescens JY3 | 13.33 ± 11.55 b | 60 | 75 |
| 6. F2 + F. oxysporum | 06.67 ± 11.55 b | 80 | n.d. |
| 7. JY3 + F. oxysporum | 06.67 ± 11.55 b | 80 | n.d. |
| 8. F2 + R. solani | 06.67 ± 11.55 b | n.d. | 87.49 |
| 9. JY3 + R. solani | 20.00 ± 00.00 b | n.d. | 62.50 |
| L.S.D | 28.78 |
| Treatment | Fresh Weight of Shoots (g) | Increase Y (%) | Increase Z (%) | Fresh Weight of Roots (g) | Increase Y (%) | Increase Z (%) |
| 1. F. oxysporum (check) | *2.07 ± 0.06 c** | 0 | n.d. | 1.67 ± 0.21 c | 0 | n.d. |
| 2. R. solani (check) | 0.97 ± 0.25 d | n.d. | 0 | 0.6 ± 0.10 d | n.d. | 0 |
| 3. Control | 3.63 ± 0.21 ab | 42.98 | 73.28 | 3.23 ± 0.38 ab | 48.30 | 81.42 |
| 4. P. aeruginosa F2 | 4.17 ± 0.15 a | 50.36 | 76.74 | 3.73 ± 0.12 a | 55.23 | 83.91 |
| 5. P. fluorescens JY3 | 3.60 ± 0.25 ab | 42.50 | 73.06 | 3.10 ± 0.17 ab | 46.13 | 80.65 |
| 6. F2 + F. oxysporum | 3.77 ± 0.15 ab | 45.09 | n.d. | 3.30 ± 0.20 ab | 49.39 | n.d. |
| 7. JY3 + F. oxysporum | 3.33 ± 0.12 b | 37.84 | n.d. | 2.93 ± 0.15 b | 43 | n.d. |
| 8. F2 + R. solani | 3.53 ± 0.29 ab | n.d. | 72.52 | 3.10 ± 0.10 ab | n.d. | 80.65 |
| 9. JY3 + R. solani | 3.27 ± 0.31 b | n.d. | 70.34 | 2.83 ± 0.25 b | n.d. | 78.80 |
| L.S.D | 0.75 | 0.78 | ||||
| Treatment | Dry Weight of Shoots (g) | Increase Y (%) | Increase Z (%) | Dry Weight of Roots (g) | Increase Y (%) | Increase Z (%) |
| 1. F. oxysporum (check) | 1.03 ± 0.15 c | 0 | n.d. | 1.03 ± 0.12 c | 0 | n.d. |
| 2. R. solani (check) | 0.33 ± 0.12 d | n.d. | 0 | 0.18 ± 0.03 d | n.d. | 0 |
| 3. Control | 2.67 ± 0.23 ab | 61.42 | 87.64 | 2.1 ± 0.06 ab | 50.95 | 91.43 |
| 4. P. aeruginosa F2 | 3.30 ± 0.10 a | 68.79 | 90 | 2.67 ± 0.15 a | 61.42 | 93.26 |
| 5. P. fluorescens JY3 | 2.73 ± 0.21 ab | 62.27 | 87.91 | 2.07 ± 0.10 ab | 50.24 | 91.3 |
| 6. F2 + F. oxysporum | 2.83 ± 0.21 ab | 63.60 | n.d. | 2.30 ± 0.10 ab | 55.22 | n.d. |
| 7. JY3 + F. oxysporum | 2.47 ± 0.29 b | 58.30 | n.d. | 2.03 ± 0.21 ab | 49.26 | n.d. |
| 8. F2 + R. solani | 2.63 ± 0.06 ab | n.d. | 87.45 | 2.20 ± 0.17 ab | n.d. | 91.82 |
| 9. JY3 + R. solani | 2.37 ± 0.15 b | n.d. | 86.08 | 1.83 ± 0.06 b | n.d. | 90.16 |
| L.S.D | 0.68 | 0.66 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abo-Zaid, G.A.; Soliman, N.A.-M.; Abdullah, A.S.; El-Sharouny, E.E.; Matar, S.M.; Sabry, S.A.-F. Maximization of Siderophores Production from Biocontrol Agents, Pseudomonas aeruginosa F2 and Pseudomonas fluorescens JY3 Using Batch and Exponential Fed-Batch Fermentation. Processes 2020, 8, 455. https://doi.org/10.3390/pr8040455
Abo-Zaid GA, Soliman NA-M, Abdullah AS, El-Sharouny EE, Matar SM, Sabry SA-F. Maximization of Siderophores Production from Biocontrol Agents, Pseudomonas aeruginosa F2 and Pseudomonas fluorescens JY3 Using Batch and Exponential Fed-Batch Fermentation. Processes. 2020; 8(4):455. https://doi.org/10.3390/pr8040455
Chicago/Turabian StyleAbo-Zaid, Gaber Attia, Nadia Abdel-Mohsen Soliman, Ahmed Salah Abdullah, Ebaa Ebrahim El-Sharouny, Saleh Mohamed Matar, and Soraya Abdel-Fattah Sabry. 2020. "Maximization of Siderophores Production from Biocontrol Agents, Pseudomonas aeruginosa F2 and Pseudomonas fluorescens JY3 Using Batch and Exponential Fed-Batch Fermentation" Processes 8, no. 4: 455. https://doi.org/10.3390/pr8040455
APA StyleAbo-Zaid, G. A., Soliman, N. A. -M., Abdullah, A. S., El-Sharouny, E. E., Matar, S. M., & Sabry, S. A. -F. (2020). Maximization of Siderophores Production from Biocontrol Agents, Pseudomonas aeruginosa F2 and Pseudomonas fluorescens JY3 Using Batch and Exponential Fed-Batch Fermentation. Processes, 8(4), 455. https://doi.org/10.3390/pr8040455