Antibacterial Activity of Chitosan Nanoparticles: A Review




Abstract
:1. Introduction
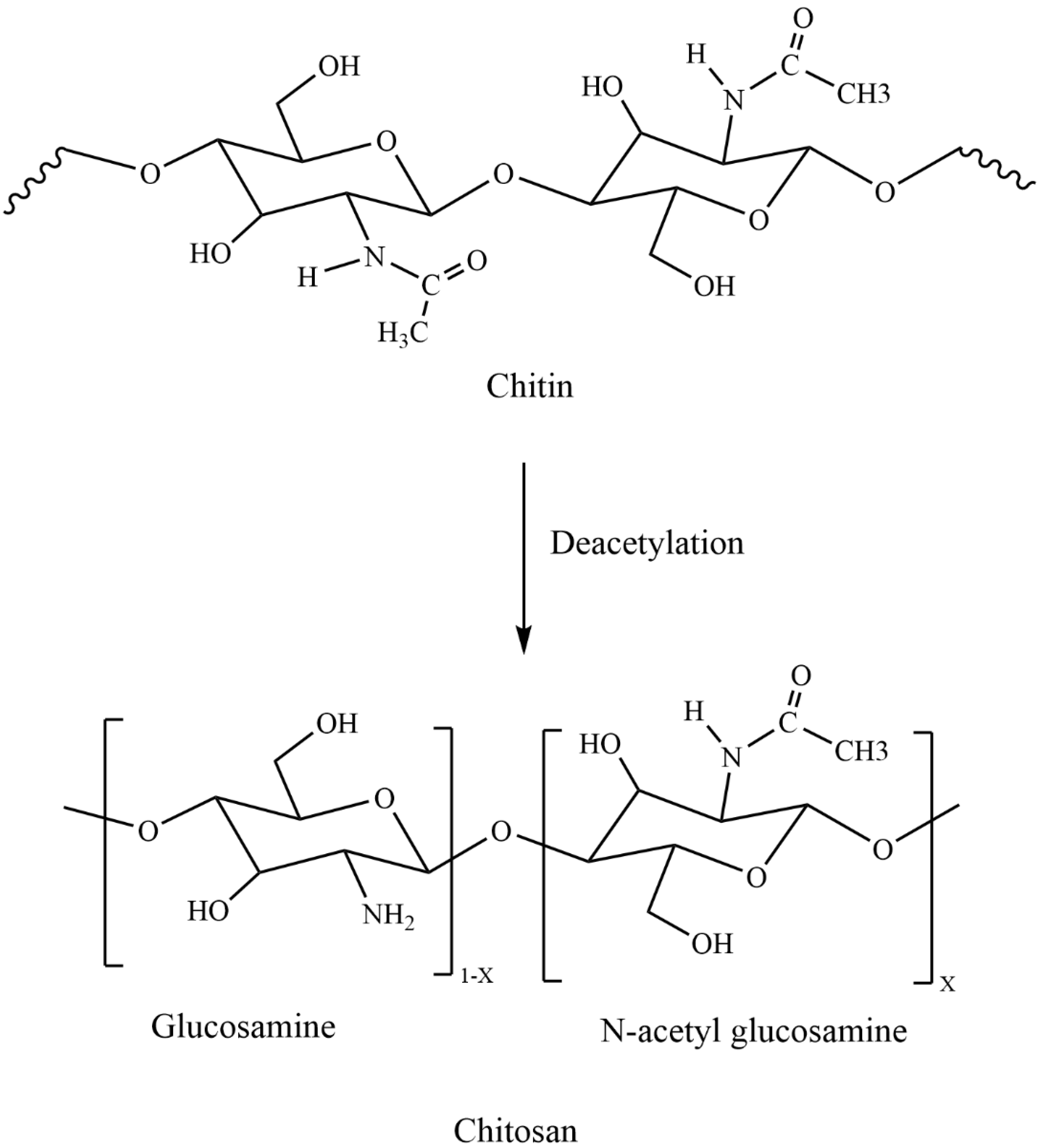
2. Chitosan
3. Chitosan Nanoparticles (CS-NPs)
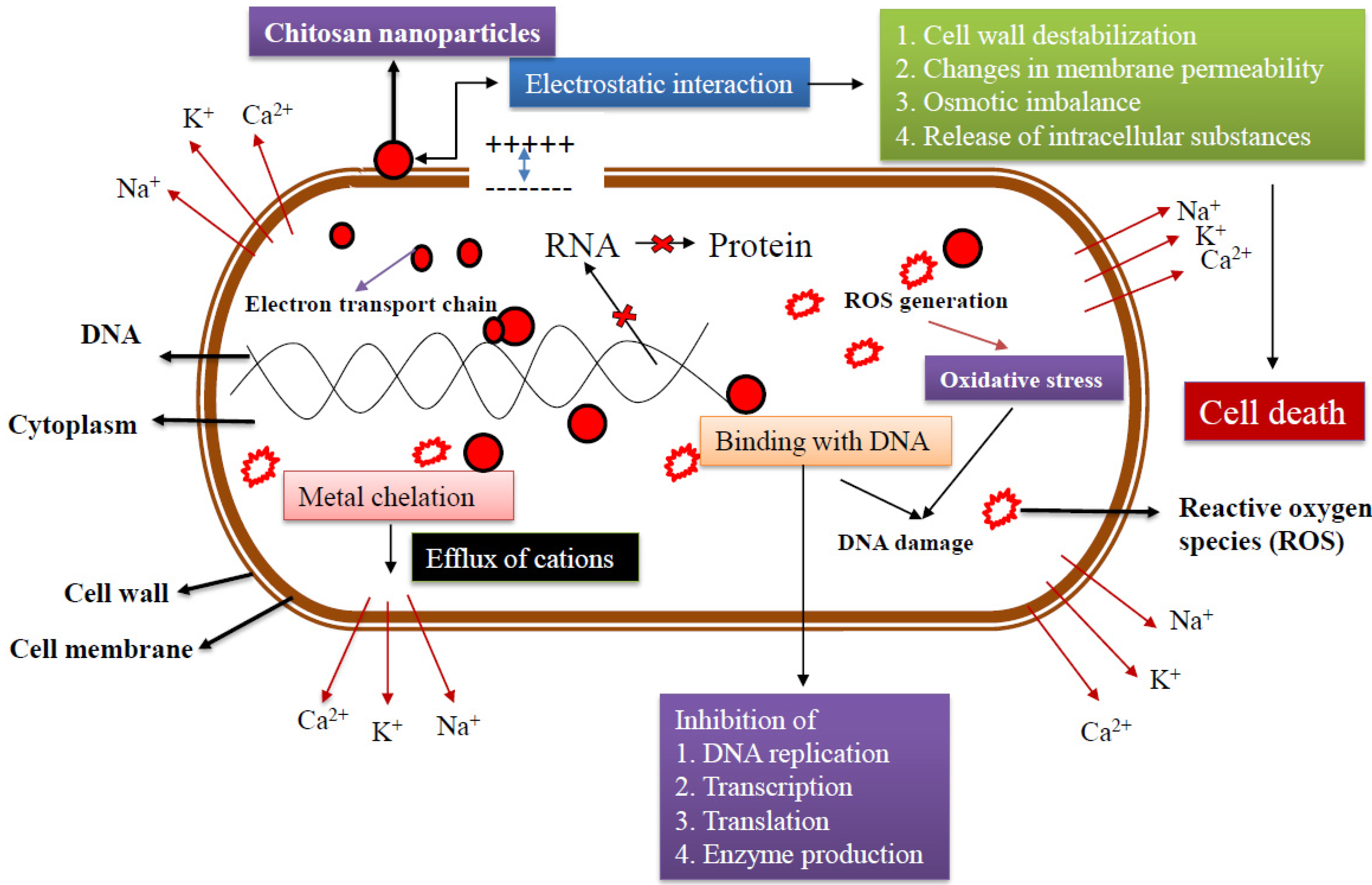
4. Mechanism of Antibacterial Action
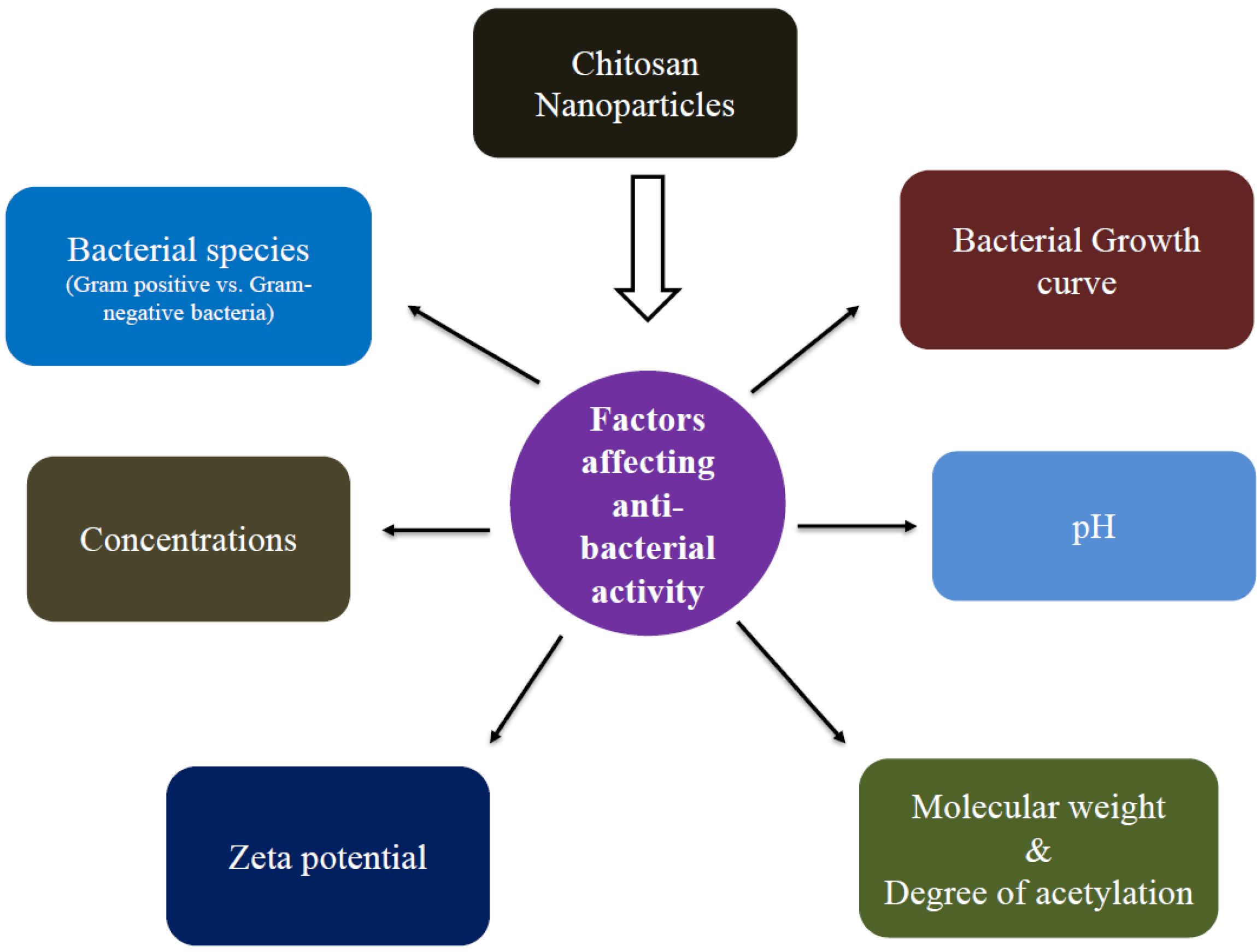
5. Factors Affecting Antibacterial Activity
5.1. Bacterial Species (Gram-Negative (G−) vs. Gram-Positive Bacteria (G+))
5.2. Bacterial Growth Stage
5.3. Zeta Potential
5.4. pH
5.5. Concentrations
5.6. Influence of Molecular Weight and Degree of Acetylation
6. Applications
6.1. Antibacterial Activity against Animal Pathogens
6.2. Antibacterial Activity against Plant Pathogens
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Goy, R.C.; De Britto, D.; Assis, O.B.G. A review of the antimicrobial activity of chitosan. Polímeros 2009, 19, 241–247. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Rabea, E.I.; Badawy, M.E.T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as antimicrobial agent: Applications and mode of action. Biomacromolecules 2003, 4, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, X.; Chen, R.; Huangfu, W.; Xie, G.-L. Antibacterial activity of chitosan solution against Xanthomonas pathogenic bacteria isolated from Euphorbia pulcherrima. Carbohydr. Polym. 2008, 72, 287–292. [Google Scholar] [CrossRef]
- Jia, Z.; Shen, D.; Xu, W. Synthesis and antibacterial activities of quaternary ammonium salt of chitosan. Carbohydr. Res. 2001, 333, 1–6. [Google Scholar] [CrossRef]
- Vinsová, J.; Vavříková, E. Chitosan derivatives with antimicrobial, antitumour and antioxidant Activities—A Review. Curr. Pharm. Des. 2011, 17, 3596–3607. [Google Scholar] [CrossRef]
- Wang, X.; Du, Y.; Fan, L.; Liu, H.; Hu, Y. Chitosan-metal complexes as antimicrobial agent: Synthesis, characterization and structure-activity study. Polym. Bull. 2005, 55, 105–113. [Google Scholar] [CrossRef]
- Yang, T.-C.; Chou, C.-C.; Li, C.-F. Antibacterial activity of N-alkylated disaccharide chitosan derivatives. Int. J. Food Microbiol. 2005, 97, 237–245. [Google Scholar] [CrossRef]
- Kammoun, M.; Haddar, M.; Kallel, T.K.; Dammak, M.; Sayari, A. Biological properties and biodegradation studies of chitosan biofilms plasticized with PEG and glycerol. Int. J. Boil. Macromol. 2013, 62, 433–438. [Google Scholar] [CrossRef]
- Ma, Z.; Garrido-Maestu, A.; Jeong, K.C. Application, mode of action, and in vivo activity of chitosan and its micro- and nanoparticles as antimicrobial agents: A review. Carbohydr. Polym. 2017, 176, 257–265. [Google Scholar] [CrossRef]
- Divya, K.; Vijayan, S.; George, T.K.; Jisha, M.S. Antimicrobial properties of chitosan nanoparticles: Mode of action and factors affecting activity. Fibers Polym. 2017, 18, 221–230. [Google Scholar] [CrossRef]
- Divya, K.; Jisha, M.S. Chitosan nanoparticles preparation and applications. Environ. Chem. Lett. 2017, 16, 101–112. [Google Scholar] [CrossRef]
- Kassem, A.; Ayoub, G.M.; Malaeb, L. Antibacterial activity of chitosan nano-composites and carbon nanotubes: A review. Sci. Total. Environ. 2019, 668, 566–576. [Google Scholar] [CrossRef]
- Rozman, N.A.S.; Tong, W.Y.; Leong, C.R.; Tan, W.N.; Hasanolbasori, M.A.; Abdullah, S.Z. Potential antimicrobial applications of chitosan nanoparticles (ChNP). J. Microbiol. Biotechnol. 2019, 29, 1009–1013. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Aljaeid, B.M. Preparation, characterization, and potential application of chitosan, chitosan derivatives, and chitosan metal nanoparticles in pharmaceutical drug delivery. Drug Des. Dev. Ther. 2016, 10, 483–507. [Google Scholar] [CrossRef] [Green Version]
- Vishwakarma, A.; Sriram, P.; Preetha, S.P.; Tirumurugaan, K.G.; Nagarajan, K.; Pandian, K. Synthesis and characterization of chitosan/TPP encapsulated curcumin nanoparticles and its antibacterial efficacy against colon bacteria. Int. J. Chem. Stud. 2019, 7, 602–606. [Google Scholar]
- Perinelli, D.R.; Fagioli, L.; Campana, R.; Lam, J.K.; Baffone, W.; Palmieri, G.F.; Bonacucina, G. Chitosan-based nanosystems and their exploited anti-microbial activity. Eur. J. Pharm. Sci. 2018, 117, 8–20. [Google Scholar] [CrossRef]
- Kravanja, G.; Primožič, M.; Knez, Ž; Leitgeb, M. Chitosan-Based (nano) materials for novel biomedical applications. Molecules 2019, 24, 1960. [Google Scholar] [CrossRef] [Green Version]
- Tokumitsu, H.; Ichikawa, H.; Fukumori, Y. Chitosan-gadopentetic acid complex nanoparticles for gadolinium neutron-capture therapy of cancer: Preparation by novel emulsion-droplet coalescence technique and characterization. Pharm. Res. 1999, 16, 1830–1835. [Google Scholar] [CrossRef]
- Supraja, N.; Thiruchenduran, S.; Prasad, T. Synthesis and characterization of chitosan nanoparticles and evaluation of antimicrobial activity antioxidant activity. Adv. Bioequiv. Bioavailab. 2018, 2, 88–93. [Google Scholar]
- MubarakAli, D.; LewisOscar, F.; Gopinath, V.; Alharbi, N.S.; Alharbi, S.A.; Thajuddin, N. An inhibitory action of chitosan nanoparticles against pathogenic bacteria and fungi and their potential applications as biocompatible antioxidants. Microb. Pathog. 2018, 114, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Xu, Z.; Jiang, X.; Hu, C.; Zou, X. Preparation and antibacterial activity of chitosan nanoparticles. Carbohydr. Res. 2004, 339, 2693–2700. [Google Scholar] [CrossRef] [PubMed]
- Maluin, F.N.; Hussein, M. Chitosan-based agronanochemicals as a sustainable alternative in crop protection. Molecules 2020, 25, 1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elieh-Ali-Komi, D.; Hamblin, M.R. Chitin and chitosan: Production and application of versatile biomedical nanomaterials. Int. J. Adv. Res. 2016, 4, 411–427. [Google Scholar]
- Hsu, S.; Chang, Y.-B.; Tsai, C.-L.; Fu, K.-Y.; Wang, S.-H.; Tseng, H.-J. Characterization and biocompatibility of chitosan nanocomposites. Colloids Surfaces B Biointerfaces 2011, 85, 198–206. [Google Scholar] [CrossRef]
- Mi, F.-L.; Tan, Y.-C.; Liang, H.-F.; Sung, H. In vivo biocompatibility and degradability of a novel injectable-chitosan-based implant. Biomaterials 2002, 23, 181–191. [Google Scholar] [CrossRef]
- Kim, H.; Tator, C.H.; Shoichet, M.S. Chitosan implants in the rat spinal cord: Biocompatibility and biodegradation. J. Biomed. Mater. Res. Part A 2011, 97, 395–404. [Google Scholar] [CrossRef]
- Shi, Z.; Neoh, K.G.; Kang, E.-T.; Wang, W. Antibacterial and mechanical properties of bone cement impregnated with chitosan nanoparticles. Biomaterials 2006, 27, 2440–2449. [Google Scholar] [CrossRef]
- Rabea, E.; Steurbaut, W. Chemically modified chitosans as antimicrobial agents against some plant pathogenic bacteria and fungi. Plant Prot. Sci. 2010, 46, 149–158. [Google Scholar] [CrossRef]
- Bernkop-Schnürch, A.; Dünnhaupt, S. Chitosan-based drug delivery systems. Eur. J. Pharm. Biopharm. 2012, 81, 463–469. [Google Scholar] [CrossRef]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, G.; Chen, X.; Yin, Y.; Zheng, H.; Xiong, X.; Du, Y. Synthesis, characterization and antibacterial activity of salicyloyl chitosan. Carbohydr. Polym. 2011, 83, 1274–1278. [Google Scholar] [CrossRef]
- Sajomsang, W.; Tantayanon, S.; Tangpasuthadol, V.; Daly, W.H. Synthesis of methylated chitosan containing aromatic moieties: Chemoselectivity and effect on molecular weight. Carbohydr. Polym. 2008, 72, 740–750. [Google Scholar] [CrossRef]
- Tan, H.-L.; Ma, R.; Lin, C.; Liu, Z.; Tang, T. Quaternized chitosan as an antimicrobial agent: Antimicrobial activity, mechanism of action and biomedical applications in orthopedics. Int. J. Mol. Sci. 2013, 14, 1854–1869. [Google Scholar] [CrossRef]
- Dutta, P.; Tripathi, S.; Mehrotra, G.; Dutta, J. Perspectives for chitosan based antimicrobial films in food applications. Food Chem. 2009, 114, 1173–1182. [Google Scholar] [CrossRef]
- Ray, S.D. Potential aspects of chitosan as pharmaceutical excipient. Acta Pol. Pharm. Drug Res. 2011, 68, 619–622. [Google Scholar]
- Upadhyaya, L.; Singh, J.; Agarwal, V.; Tewari, R.P. Biomedical applications of carboxymethyl chitosans. Carbohydr. Polym. 2013, 91, 452–466. [Google Scholar] [CrossRef]
- Cota-Arriola, O.; Cortez-Rocha, M.O.; Burgos-Hernández, A.; Ezquerra-Brauer, J.M.; Plascencia-Jatomea, M. Controlled release matrices and micro/nanoparticles of chitosan with antimicrobial potential: Development of new strategies for microbial control in agriculture. J. Sci. Food Agric. 2013, 93, 1525–1536. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef]
- Liu, H.; Du, Y.M.; Wang, X.H.; Sun, L.P. Chitosan kills bacteria through cell membrane damage. Int. J. Food Microbiol. 2004, 95, 147–155. [Google Scholar] [CrossRef]
- Calvo, P.; Remuñan-López, C.; Vila-Jato, J.L.; Alonso, M.J. Chitosan and chitosan/ethylene oxide-propylene oxide block copolymer nanoparticles as novel carriers for proteins and vaccines. Pharm. Res. 1997, 14, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Brunel, F.; Varon, L.; David, L.; Domard, A.; Delair, T. A novel syn-thesis of chitosan nanoparticles in reverse emulsion. Langmuir 2008, 24, 11370–11377. [Google Scholar] [CrossRef] [PubMed]
- Maitra, A.; Ghosh, P.K.; De, T.K.; Sahoo, S.K. Process for the Preparation of Highly Monodispersed Polymeric hydrophilic Nanoparticles. U.S. Patent 587411, 23 February 1999. [Google Scholar]
- Davis, L.I.S.S. Sustained release chitosan microspheres prepared by novel spray drying methods. J. Microencapsul. 1999, 16, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Jiang, H.; Ye, H.; Li, J.; Huang, J. Preparation, antibacterial, and antioxidant activities of silver/chitosan composites. J. Carbohydr. Chem. 2014, 33, 298–312. [Google Scholar] [CrossRef]
- Qian, J.; Pan, C.; Liang, C. Antimicrobial activity of Fe-loaded chitosan nanoparticles. Eng. Life Sci. 2017, 17, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Badawy, M.E.I.; Lotfy, T.M.R.; Shawir, S.M.S. Preparation and antibacterial activity of chitosan-silver nanoparticles for application in preservation of minced meat. Bull. Natl. Res. Cent. 2019, 43, 83. [Google Scholar] [CrossRef]
- Katas, H.; Lim, C.S.; Azlan, A.Y.H.N.; Buang, F.; Busra, M.F.M. Antibacterial activity of biosynthesized gold nanoparticles using biomolecules from Lignosus rhinocerotis and chitosan. Saudi Pharm. J. 2018, 27, 283–292. [Google Scholar] [CrossRef]
- Tamara, F.R.; Lin, C.; Mi, F.-L.; Ho, Y.-C. Antibacterial effects of chitosan/cationic peptide nanoparticles. Nanomaterials 2018, 8, 88. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Wu, C.; Fu, S.; Wang, L.; Yuan, C.; Chen, S.; Hu, Y. Integration of lysozyme into chitosan nanoparticles for improving antibacterial activity. Carbohydr. Polym. 2017, 155, 192–200. [Google Scholar] [CrossRef]
- Huang, L.; Cheng, X.; Liu, C.; Xing, K.; Zhang, J.; Sun, G.; Li, X.; Chen, X. Preparation, characterization, and antibacterial activity of oleic acid-grafted chitosan oligosaccharide nanoparticles. Front. Biol. China 2009, 4, 321–327. [Google Scholar] [CrossRef]
- Sotelo-Boyás, M.; Correa-Pacheco, Z.; Bautista-Baños, S.; Corona-Rangel, M. Physicochemical characterization of chitosan nanoparticles and nanocapsules incorporated with lime essential oil and their antibacterial activity against food-borne pathogens. LWT 2017, 77, 15–20. [Google Scholar] [CrossRef]
- Ali, M.J.; Sharafaldin, A.M.; Majid, P.; Mahdi, F.R.; Kazem, A.; Hajar, R.; Zuhair, M.H.; Mahdi, K.M.; Reza, M. Curcumin-loaded chitosan tripolyphosphate nanoparticles as a safe, natural and effective antibiotic inhibits the infection of Staphylococcus aureus and Pseudomonas aeruginosa in vivo. Iran. J. Biotechnol. 2014, 12, 1–8. [Google Scholar]
- Avadi, M.; Sadeghi, A.; Tahzibi, A.; Bayati, K.; Pouladzadeh, M.; Zohuriaan-Mehr, M.; Rafiee-Tehrani, M. Diethylmethyl chitosan as an antimicrobial agent: Synthesis, characterization and antibacterial effects. Eur. Polym. J. 2004, 40, 1355–1361. [Google Scholar] [CrossRef]
- Du, W.-L.; Niu, S.-S.; Xu, Y.-L.; Xu, Z.-R.; Fan, C.-L. Antibacterial activity of chitosan tripolyphosphate nanoparticles loaded with various metal ions. Carbohydr. Polym. 2009, 75, 385–389. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Nguyen, T.T.H.; Wang, S.-L.; Vo, T.P.K.; Nguyen, A.D. Preparation of chitosan nanoparticles by TPP ionic gelation combined with spray drying, and the antibacterial activity of chitosan nanoparticles and a chitosan nanoparticle–amoxicillin complex. Res. Chem. Intermed. 2016, 43, 3527–3537. [Google Scholar] [CrossRef]
- Ngan, L.T.K.; Wang, S.-L.; Hiep, Đ.M.; Luong, P.M.; Nguyen, V.B.; Đinh, T.M.; Dzung, N.A. Preparation of chitosan nanoparticles by spray drying, and their antibacterial activity. Res. Chem. Intermed. 2014, 40, 2165–2175. [Google Scholar] [CrossRef]
- Piras, A.M.; Maisetta, G.; Sandreschi, S.; Gazzarri, M.; Bartoli, C.; Grassi, L.; Esin, S.; Chiellini, F.; Batoni, G. Chitosan nanoparticles loaded with the antimicrobial peptide temporin B exert a long-term antibacterial activity in vitro against clinical isolates of Staphylococcus epidermidis. Front. Microbiol. 2015, 6, 372. [Google Scholar] [CrossRef] [Green Version]
- Tsai, G.-J.; Su, W.-H. Antibacterial activity of shrimp chitosan against Escherichia coli. J. Food Prot. 1999, 62, 239–243. [Google Scholar] [CrossRef]
- Sudarshan, N.; Hoover, D.; Knorr, D. Antibacterial action of chitosan. Food Biotechnol. 1992, 6, 257–272. [Google Scholar] [CrossRef]
- Shahidi, F.; Arachchi, J.K.V.; Jeon, Y.-J. Food applications of chitin and chitosans. Trends Food Sci. Technol. 1999, 10, 37–51. [Google Scholar] [CrossRef]
- Raafat, D.; Von Bargen, K.; Haas, A.; Sahl, H.-G. Insights into the mode of action of chitosan as an antibacterial compound. Appl. Environ. Microbiol. 2008, 74, 3764–3773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cava, F.; Lam, H.; De Pedro, M.A.; Waldor, M.K. Emerging knowledge of regulatory roles of d-amino acids in bacteria. Cell. Mol. Life Sci. 2010, 68, 817–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivask, A.; Elbadawy, A.; Kaweeteerawat, C.; Boren, D.; Fischer, H.; Ji, Z.; Chang, C.H.; Liu, R.; Tolaymat, T.; Telesca, D.; et al. Toxicity mechanisms in Escherichia coli vary for silver nanoparticles and differ from ionic silver. ACS Nano 2013, 8, 374–386. [Google Scholar] [CrossRef] [PubMed]
- Birsoy, K.; Wang, T.C.; Chen, W.W.; Freinkman, E.; Abu-Remaileh, M.; Sabatini, D.M. An essential role of the mitochondrial electron transport chain in cell proliferation is to enable aspartate synthesis. Cell 2015, 162, 540–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eltahlawy, K.; El-Bendary, M.A.; Elhendawy, A.; Hudson, S. The antimicrobial activity of cotton fabrics treated with different crosslinking agents and chitosan. Carbohydr. Polym. 2005, 60, 421–430. [Google Scholar] [CrossRef]
- Joshi, M.; Ali, S.W.; Purwar, R. Ecofriendly antimicrobial finishing of textile using bioactive agents based on natural products. Indian J. Fibre Text. Res. 2009, 30, 295–304. [Google Scholar]
- Wang, X.; Du, Y.; Liu, H. Preparation, characterization and antimicrobial activity of chitosan–Zn complex. Carbohydr. Polym. 2004, 56, 21–26. [Google Scholar] [CrossRef]
- Chung, Y.-C.; Su, Y.-P.; Chen, C.-C.; Jia, G.; Wang, H.-L.; Wu, J.C.G.; Lin, J.-G. Relationship between antibacterial activity of chitosan and surface characteristics of cell wall. Acta Pharmacol. Sin. 2004, 25, 932–936. [Google Scholar]
- No, H.K.; Park, N.Y.; Lee, S.H.; Meyers, S.P. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int. J. Food Microbiol. 2002, 74, 65–72. [Google Scholar] [CrossRef]
- Zhong, Z.; Xing, R.; Liu, S.; Wang, L.; Cai, S.; Li, P. Synthesis of acyl thiourea derivatives of chitosan and their antimicrobial activities in vitro. Carbohydr. Res. 2008, 343, 566–570. [Google Scholar] [CrossRef]
- Cao, X.L.; Cheng, C.; Ma, Y.L.; Zhao, C.S. Preparation of silver nanoparticles with antimicrobial activities and the researches of their biocompatibilities. J. Mater. Sci. Mater. Electron. 2010, 21, 2861–2868. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.V.; Tran, L.D.; Ba, C.T.; Vu, H.D.; Nguyen, T.N.; Pham, D.G.; Nguyen, P.X. Synthesis, characterization, antibacterial and antiproliferative activities of monodisperse chitosan-based silver nanoparticles. Colloids Surfaces A Physicochem. Eng. Asp. 2010, 360, 32–40. [Google Scholar] [CrossRef]
- Perelshtein, I.; Ruderman, E.; Perkas, N.; Tzanov, T.; Beddow, J.; Joyce, E.; Mason, T.; Blanes, M.; Mollá, K.; Patlolla, A.; et al. Chitosan and chitosan-ZnO-based complex nanoparticles: Formation, characterization, and antibacterial activity. J. Mater. Chem. B 2013, 1, 1968–1976. [Google Scholar] [CrossRef] [PubMed]
- Devlieghere, F.; Vermeulen, A.; Debevere, J. Chitosan: Antimicrobial activity, interactions with food components and applicability as a coating on fruit and vegetables. Food Microbiol. 2004, 21, 703–714. [Google Scholar] [CrossRef]
- Venkatesham, M.; Ayodhya, D.; Madhusudhan, A.; Babu, N.V.; Veerabhadram, G. A novel green one-step synthesis of silver nanoparticles using chitosan: Catalytic activity and antimicrobial studies. Appl. Nanosci. 2012, 4, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Rasaee, I.; Ghannadnia, M.; Honari, H. Antibacterial properties of biologically formed chitosan nanoparticles using aqueous leaf extract of Ocimum basilicum. Nanomed. J. 2016, 3, 147–154. [Google Scholar]
- Aleanizy, F.S.; Alqahtani, F.Y.; Shazly, G.; Alfaraj, R.; Alsarra, I.; Alshamsan, A.; Abdulhady, H.G. Measurement and evaluation of the effects of pH gradients on the antimicrobial and antivirulence activities of chitosan nanoparticles in Pseudomonas aeruginosa. Saudi Pharm. J. 2017, 26, 79–83. [Google Scholar] [CrossRef]
- Wei, D.; Sun, W.; Qian, W.; Ye, Y.; Ma, X. The synthesis of chitosan-based silver nanoparticles and their antibacterial activity. Carbohydr. Res. 2009, 344, 2375–2382. [Google Scholar] [CrossRef]
- Muthukrishnan, A.M.; Manik, A.; Sathiyabama, M. Green synthesis of copper-chitosan nanoparticles and study of its antibacterial activity. J. Nanomed. Nanotechnol. 2015, 6, 251. [Google Scholar] [CrossRef]
- Sarwar, A.; Katas, H.; Zin, N.M. Antibacterial effects of chitosan–tripolyphosphate nanoparticles: Impact of particle size molecular weight. J. Nanoparticle Res. 2014, 16, 2517. [Google Scholar] [CrossRef]
- Orellano, M.S.; Isaac, P.; Breser, M.L.; Bohl, L.P.; Conesa, A.; Falcone, R.D.; Porporatto, C. Chitosan nanoparticles enhance the antibacterial activity of the native polymer against bovine mastitis pathogens. Carbohydr. Polym. 2019, 213, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ikono, R.; Vibriani, A.; Wibowo, I.; Saputro, K.E.; Muliawan, W.; Bachtiar, B.M.; Mardliyati, E.; Bachtiar, E.W.; Rochman, N.T.; Kagami, H.; et al. Nanochitosan antimicrobial activity against Streptococcus mutans and Candida albicans dual-species biofilms. BMC Res. Notes 2019, 12, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.-C.; Li, C.-F.; Chou, C.-C. Cell age, suspending medium and metal ion influence the susceptibility of Escherichia coli O157:H7 to water-soluble maltose chitosan derivative. Int. J. Food Microbiol. 2007, 113, 258–262. [Google Scholar] [CrossRef]
- Xing, K.; Chen, X.; Li, Y.Y.; Liu, C.; Liu, C.G.; Cha, D.S.; Park, H.J. Antibacterial activity of oleoyl-chitosan nanoparticles: A novel antibacterial dispersion system. Carbohydr. Polym. 2008, 74, 114–120. [Google Scholar] [CrossRef]
- Wang, G.-H. Inhibition and inactivation of five species of foodborne pathogens by chitosan. J. Food Prot. 1992, 55, 916–919. [Google Scholar] [CrossRef]
- Gomes, L.P.; Andrade, C.T.; Del Aguila, E.M.; Alexander, C.; Paschoalin, V.M. Assessing the antimicrobial activity of chitosan nanoparticles by fluorescence-labeling. Int. J. Biotechnol. Bioeng. 2018, 12, 111–117. [Google Scholar]
- Anitha, A.; Rani, V.D.; Krishna, R.; Sreeja, V.; Selvamurugan, N.; Nair, S.; Tamura, H.; Jayakumar, R. Synthesis, characterization, cytotoxicity and antibacterial studies of chitosan, O-carboxymethyl and N,O-carboxymethyl chitosan nanoparticles. Carbohydr. Polym. 2009, 78, 672–677. [Google Scholar] [CrossRef]
- Mohammadi, A.; Hashemi, M.; Hosseini, S.M. Effect of chitosan molecular weight as micro and nanoparticles on antibacterial activity against some soft rot pathogenic bacteria. LWT 2016, 71, 347–355. [Google Scholar] [CrossRef]
- Liu, N.; Chen, X.; Park, H.J.; Liu, C.; Liu, C.; Meng, X.-H.; Yu, L.-J. Effect of MW and concentration of chitosan on antibacterial activity of Escherichia coli. Carbohydr. Polym. 2006, 64, 60–65. [Google Scholar] [CrossRef]
- Sanpui, P.; Murugadoss, A.; Prasad, P.; Ghosh, S.; Chattopadhyay, A. The antibacterial properties of a novel chitosan–Ag-nanoparticle composite. Int. J. Food Microbiol. 2008, 124, 142–146. [Google Scholar] [CrossRef]
- Alarfaj, A.A. Preparation, characterization and antibacterial effect of chitosan nanoparticles against food spoilage bacteria. J. Pure Appl. Microbiol. 2019, 13, 1273–1278. [Google Scholar] [CrossRef] [Green Version]
- Gan, Q.; Wang, T.; Cochrane, C.; McCarron, P.A. Modulation of surface charge, particle size and morphological properties of chitosan–TPP nanoparticles intended for gene delivery. Colloids Surfaces B Biointerfaces 2005, 44, 65–73. [Google Scholar] [CrossRef]
- De Paz, L.E.C.; Resin, A.; Howard, K.A.; Sutherland, D.S.; Wejse, P.L. Antimicrobial effect of chitosan nanoparticles on Streptococcus mutans biofilms. Appl. Environ. Microbiol. 2011, 77, 3892–3895. [Google Scholar] [CrossRef] [Green Version]
- Yildirim-Aksoy, M.; Beck, B.H. Antimicrobial activity of chitosan and a chitosan oligomer against bacterial pathogens of warmwater fish. J. Appl. Microbiol. 2017, 122, 1570–1578. [Google Scholar] [CrossRef]
- Garrido-Maestu, A.; Ma, Z.; Paik, S.-Y.-R.; Chen, N.; Ko, S.-H.; Tong, Z.; Jeong, K.C. Engineering of chitosan-derived nanoparticles to enhance antimicrobial activity against foodborne pathogen Escherichia coli O157:H7. Carbohydr. Polym. 2018, 197, 623–630. [Google Scholar] [CrossRef]
- Alqahtani, F.Y.; Aleanizy, F.S.; El Tahir, E.; AlQuadeib, B.; Alsarra, I.; Alanazi, J.S.; Abdelhady, H.G. Preparation, characterization, and antibacterial activity of diclofenac-loaded chitosan nanoparticles. Saudi Pharm. J. 2018, 27, 82–87. [Google Scholar] [CrossRef]
- Honary, S.; Ghajar, K.; Khazaeli, P.; Shalchian, P. Preparation, characterization and antibacterial properties of silver-chitosan nanocomposites using different molecular weight grades of chitosan. Trop. J. Pharm. Res. 2011, 10, 74. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.; Liu, C.; Meng, X.H.; Yu, L.J.; Liu, C.G. Antibacterial mechanism of chitosan microspheres in a solid dispersing system against Escherchia coli. Colloids Surfaces B Biointerfaces 2008, 65, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Regiel, A.; Irusta, S.; Kyzioł, A.; Arruebo, M.; Santamaria, J. Preparation and characterization of chitosan–silver nanocomposite films and their antibacterial activity against Staphylococcus aureus. Nanotechnology 2012, 24, 015101. [Google Scholar] [CrossRef]
- Covarrubias, C.; Trepiana, D.; Corral, C. Synthesis of hybrid copper-chitosan nanoparticles with antibacterial activity against cariogenic Streptococcus mutans. Dent. Mater. J. 2018, 37, 379–384. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Cristóbal, L.F.; Martinez-Castañon, G.A.; Martínez-Martínez, R.; Loyola-Rodríguez, J.P.; Patiño-Marín, N.; Reyes-Macías, J.; Ruiz, F. Antibacterial effect of silver nanoparticles against Streptococcus mutans. Mater. Lett. 2009, 63, 2603–2606. [Google Scholar] [CrossRef]
- Besinis, A.; De Peralta, T.; Handy, R.D. The antibacterial effects of silver, titanium dioxide and silica dioxide nanoparticles compared to the dental disinfectant chlorhexidine on Streptococcus mutans using a suite of bioassays. Nanotoxicology 2013, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliasghari, A.; Khorasgani, M.R.; Vaezifar, S.; Rahimi, F.; Younesi, H.; Khoroushi, M. Evaluation of antibacterial efficiency of chitosan and chitosan nanoparticles on cariogenic streptococci: An in vitro study. Iran. J. Microbiol. 2016, 8, 93–100. [Google Scholar] [PubMed]
- Costa, E.; Silva, S.; Tavaria, F.; Pintado, M. Study of the effects of chitosan upon Streptococcus mutans adherence and biofilm formation. Anaerobe 2013, 20, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Hipalaswins, W.M.; Balakumaran, M.D.; Jagadeeswari, S. Synthesis, characterization and antibacterial activity of chitosan nanoparticles and its impact on seed germination. J. Acad. Ind.Res. 2016, 5, 65–71. [Google Scholar]
- Taponen, S.; Pyörälä, S. Coagulase-negative staphylococci as cause of bovine mastitis—Not so different from Staphylococcus aureus? Veter. Microbiol. 2009, 134, 29–36. [Google Scholar] [CrossRef]
- Cardozo, V.F.; Lancheros, C.A.; Narciso, A.M.; Valereto, E.C.; Kobayashi, R.K.; Seabra, A.B.; Nakazato, G. Evaluation of antibacterial activity of nitric oxide-releasing polymeric particles against Staphylococcus aureus and Escherichia coli from bovine mastitis. Int. J. Pharm. 2014, 473, 20–29. [Google Scholar] [CrossRef]
- Li, B.; Zhang, Y.; Yang, Y.; Qiu, W.-J.; Wang, X.; Liu, B.; Wang, Y.; Sun, G. Synthesis, characterization, and antibacterial activity of chitosan/TiO2 nanocomposite against Xanthomonas oryzae pv. oryzae. Carbohydr. Polym. 2016, 152, 825–831. [Google Scholar] [CrossRef]
- Esyanti, R.R.; Farah, N.; Bajra, B.D.; Nofitasari, D.; Martien, R.; Sunardi, S.; Safitri, R. Comparative study of nano-chitosan and synthetic bactericide application on chili pepper (Capsicum annuum L.) infected by Xanthomonas campestris. AGRIVITA J. Agric. Sci. 2020, 42, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.-W.; Chun, S.C.; Chandrasekaran, M. Preparation and in vitro characterization of chitosan nanoparticles and their broad-spectrum antifungal action compared to antibacterial activities against phytopathogens of tomato. Agronomy 2019, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Santiago, T.R.; Bonatto, C.C.; Rossato, M.; Lopes, C.A.P.; Lopes, C.A.; Mizubuti, E.S.G.; Silva, L.P. Green synthesis of silver nanoparticles using tomato leaf extract and their entrapment in chitosan nanoparticles to control bacterial wilt. J. Sci. Food Agric. 2019, 99, 4248–4259. [Google Scholar] [CrossRef] [PubMed]
- Pilon, L.; Spricigo, P.C.; Miranda, M.; De Moura, M.R.; Assis, O.B.G.; Mattoso, L.H.C.; Ferreira, M.D. Chitosan nanoparticle coatings reduce microbial growth on fresh-cut apples while not affecting quality attributes. Int. J. Food Sci. Technol. 2014, 50, 440–448. [Google Scholar] [CrossRef]
- Li, B.; Liu, B.; Shan, C.-L.; Ibrahim, M.; Lou, Y.; Wang, Y.; Xie, G.; Li, H.-Y.; Sun, G. Antibacterial activity of two chitosan solutions and their effect on rice bacterial leaf blight and leaf streak. Pest Manag. Sci. 2012, 69, 312–320. [Google Scholar] [CrossRef]
- Mansilla, A.Y.; Albertengo, L.; Rodriguez, M.S.; Debbaudt, A.; Zúñiga, A.; Casalongué, C.A. Evidence on antimicrobial properties and mode of action of a chitosan obtained from crustacean exoskeletons on Pseudomonas syringae pv. tomato DC3000. Appl. Microbiol. Biotechnol. 2013, 97, 6957–6966. [Google Scholar] [CrossRef]
- El Ghaouth, A. Effect of chitosan on cucumber plants: Suppression of Pythium aphanidermatum and induction of defense reactions. Phytopathology 1994, 84, 313. [Google Scholar] [CrossRef]
- Amborabé, B.-E.; Bonmort, J.; Fleurat-Lessard, P.; Roblin, G. Early events induced by chitosan on plant cells. J. Exp. Bot. 2008, 59, 2317–2324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badawy, M.E.I.; Rabea, E.I.; Taktak, N.E. Antimicrobial and inhibitory enzyme activity of N-(benzyl) and quaternary N-(benzyl) chitosan derivatives on plant pathogens. Carbohydr. Polym. 2014, 111, 670–682. [Google Scholar] [CrossRef]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev. 2014, 35, 569–588. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, R.C.; Kumaraswamy, R.V.; Kumari, S.; Sharma, S.S.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Cu-chitosan nanoparticle boost defense responses and plant growth in maize (Zea mays L.). Sci. Rep. 2017, 7, 9754. [Google Scholar] [CrossRef]
- Algam, S.A.E.; Xie, G.; Li, B.; Yu, S.; Su, T.; Larsen, J. Effects of Paenibacillus strains and chitosan on plant growth promotion and control of Ralstonia wilt in tomato. J. Plant Pathol. 2010, 92, 593–600. [Google Scholar]
- Li, B.; Shi, Y.; Shan, C.; Zhou, Q.; Ibrahim, M.; Wang, Y.; Wu, G.; Li, H.; Xie, G.; Sun, G. Effect of chitosan solution on the inhibition of Acidovorax citrulli causing bacterial fruit blotch of watermelon. J. Sci. Food Agric. 2013, 93, 1010–1015. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Li, B.; Wu, G.; Tang, Q.; Ibrahim, M.; Li, H.; Xie, G.; Sun, G. Action of chitosan against Xanthomonas pathogenic bacteria isolated from Euphorbia pulcherrima. Molecules 2012, 17, 7028–7041. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanoparticles | Size of Nanoparticles (nm) | Zeta Potential (mV) | Antibacterial Activity | References |
|---|---|---|---|---|
| CS-Fe2+NPs | 206.4 | +28.82 | Escherichia coli, Staphylococcus aureus, Candida albicans | [46] |
| CS-Fe3+-NPs | 195.2 | +28.26 | ||
| CS-AgNP1 | 76.86 | −0.2250 | Escherichia coli and Salmonella typhimurium | [47] |
| CS-AgNP2 | 63.03 | −0.1930 | ||
| CS-AgNP3 | 61.57 | −0.0695 | ||
| CS-NP1 | 78.4 | +33.77 | Escherichia coli and Bacillus cereus | [49] |
| CS-NP2 | 150.67 | +33.63 | ||
| CS-NP3 | 201 | +32 | ||
| Protamine-CS-NP1 | 114.33 | +32.33 | ||
| Protamine-CS-NP2 | 84.8 | +30.27 | ||
| Protamine-CS-NP3 | 79.4 | +27.67 | ||
| CS-NPs | 476.2 | +17.20 | Escherichia coli and Staphylococcus aureus | [50] |
| CS-NPs | 548.1 | +12.50 | ||
| CS-Lys-NPs | 488.8 | +21.10 | ||
| CS-Lys-NPs | 613.5 | +13.6 | ||
| CS-NPs | 217 | +37.6 | Escherichia coli and Bacillus subtilis | [51] |
| Carvacrol Modified CS-NPs | 26 | +18.8 | ||
| Eugenol Modified-CS-NPs | 235 | +28.4 | ||
| CS-NPs | 53.99 | +51.37 | Escherichia coli, Salmonella choleraesuis and Staphylococcus aureus | [55] |
| CS-Ag-NPs | 90.29 | +92.05 | ||
| CS-Cu2+-NPs | 121.9 | +88.69 | ||
| CS-Zn2+-NPs | 210.9 | +86.65 | ||
| CS-Mn2+-NPs | 102.3 | +75.74 | ||
| CS-Fe2+-NPs | 95.81 | +71.42 | ||
| CS-NPs | 166.7 | +54.7 | Streptococcus pneumoniae | [56] |
| CS-NPs | 189.7 | +47.7 | ||
| CS-NPs | 334.6 | +38.6 | ||
| CS-NPs | 1230 | +34.9 | ||
| CS-NPs | 95.4 | +45.7 | Staphylococcus aureus | [57] |
| CS-NPs | 271.5 | +35.1 | ||
| CS-NPs | 335.9 | +29.6 | ||
| CS-NPs | 124 | +21.9 | Staphylococcus epidermidis | [58] |
| Temporin-B Loaded-CS-NPs | 185 | +8.8 |
| Nanoparticles | Gram-Negative/Positive Bacteria | Bacterial Species | MIC | MBC | References |
|---|---|---|---|---|---|
| Chitosan nanoparticles (CS-NPs) | G− bacteria | Escherichia coli | 1/16 | 1 | [22] |
| Escherichia coli | 1/32 | 2 | |||
| Salmonella choleraesuis | 1/16 | 2 | |||
| Salmonella typhimurium | 1/8 | 4 | |||
| G+ bacteria | Staphylococcus aureus | 1/8 | 4 | ||
| CS-NPs | G− bacteria | Escherichia coli | 1/32 | 1 | [22] |
| Escherichia coli | 1/32 | 1 | |||
| Salmonella choleraesuis | 1/32 | 1 | |||
| Salmonella typhimurium | 1/16 | 2 | |||
| G+ bacteria | Staphylococcus aureus | 1/6 | 2 | ||
| Ag-CS-NPs | G− bacteria | Escherichia coli | 32 | 128 | [45] |
| Salmonella choleraesuis | 64 | 256 | |||
| G+ bacteria | Staphylococcus aureus | 64 | 256 | ||
| Bacillus subtilis | 32 | 128 | |||
| CS-NPs | G− bacteria | Escherichia coli | 625 | 2500 | [50] |
| G+ bacteria | Bacillus subtilis | 312 | 1250 | ||
| CS-NPs | G− bacteria | Escherichia coli | 117 | 187 | [55] |
| Salmonella choleraesuis | 117 | 187 | |||
| G+ bacteria | Staphylococcus aureus | 234 | 281 | ||
| CS-Ag+NPs | G− bacteria | Escherichia coli | 3 | 6 | [55] |
| Salmonella choleraesuis | 3 | 6 | |||
| G+ bacteria | Staphylococcus aureus | 6 | 12 | ||
| CS-Cu2+NPs | G− bacteria | Escherichia coli | 9 | 12 | [55] |
| Salmonella choleraesuis | 9 | 12 | |||
| G+ bacteria | Staphylococcus aureus | 21 | 24 | ||
| CS-Zn2+NPs | G− bacteria | Escherichia coli | 18 | 24 | [55] |
| Salmonella choleraesuis | 18 | 24 | |||
| G+ bacteria | Staphylococcus aureus | 36 | 48 | ||
| CS-Mn2+NPs | G− bacteria | Escherichia coli | 73 | 97 | [55] |
| Salmonella choleraesuis | 73 | 97 | |||
| G+ bacteria | Staphylococcus aureus | 85 | 97 | ||
| CS-Fe2+NPs | G− bacteria | Escherichia coli | 121 | 195 | [55] |
| Salmonella choleraesuis | 121 | 195 | |||
| G+ bacteria | Staphylococcus aureus | 146 | 195 | ||
| Low MW CS-NPs | G− bacteria | Escherichia coli | 12.5 | 25 | [81] |
| Acinetobacter schindleri | 12.5 | 25 | |||
| Pseudomonas aeruginosa | 3.13 | 6.25 | |||
| G+ bacteria | Staphylococcus aureus | 6.25 | 12.5 | ||
| Bacillus cereus | 3.13 | 6.25 | |||
| Bacillus subtilis | 1.56 | 3.13 | |||
| Low MW CS-NPs | G− bacteria | Escherichia coli | 12.5 | 50 | [81] |
| Acinetobacter schindleri | 12.5 | 50 | |||
| Pseudomonas aeruginosa | 3.13 | 12.5 | |||
| G+ bacteria | Staphylococcus aureus | 12.5 | 25 | ||
| Bacillus cereus | 6.25 | 6.25 | |||
| Bacillus subtilis | 3.13 | 6.25 | |||
| High MW CS-NPs | G− bacteria | Escherichia coli | 25 | 50 | [81] |
| Acinetobacter schindleri | 25 | 50 | |||
| Pseudomonas aeruginosa | 12.5 | 25 | |||
| G+ bacteria | Staphylococcus aureus | 12.5 | 25 | ||
| Bacillus cereus | 6.25 | 25 | |||
| Bacillus subtilis | 6.25 | 25 | |||
| High MW CS-NPs | G− bacteria | Escherichia coli | 25 | 50 | [81] |
| Acinetobacter schindleri | 25 | 50 | |||
| Pseudomonas aeruginosa | 12.5 | 25 | |||
| G+ bacteria | Staphylococcus aureus | 12.5 | 25 | ||
| Bacillus cereus | 12.5 | 0.05 | |||
| Bacillus subtilis | 6.25 | 25 |
| Nanoparticles | Pathogen | Disease/Infection | References |
|---|---|---|---|
| CS-NPs-essential oil | Staphylococcus aureus, Listeria monocytogenes, Shigella dysenteriae, and Escherichia coli | Food-borne pathogens | [52] |
| CS-NPs-amoxicillin | Streptococcus pneumoniae | Respiratory infection | [56] |
| CS-NPs | Staphylococcus epidermidi | Opportunistic infections | [58] |
| CS-NPs | Staphylococcus aureus, Staphylococcus xylosus, Staphylococcus chromogenes, Staphylococcus haemolyticus | Bovine mastitis | [82] |
| CS-NPs | Streptococcus mutans, Candida albicans | Dental caries | [83] |
| Cu-CS-NPs | Streptococcus mutans | Dental caries | [94] |
| CS-NPs | Streptococcus mutans, Streptococcus sobrinus, Streptococcus sanguis, and Streptococcus salivarius | Cariogenic bacteria | [101] |
| CS-NPs | Pseudomonas fluorescens, Proteus mirabilis, Staphylococcus aureus, Klebsiella pneumoniae, Escherichia coli and Enterobacter aerogenes | Clinical pathogens | [106] |
| CS-NPs | Staphylococcus aureus | Bovine mastitis | [107] |
| CS-NPs | Staphylococcus aureus, Escherichia coli | Bovine mastitis | [108] |
| CS/TiO2 NPs | Xanthomonas oryzae pv. oryzae | Bacterial spot of tomato | [109] |
| CS-NPs | Xanthomonas campestris | Bacterial spot of chili-pepper | [110] |
| CS-NPs | Xanthomonas campestris and Erwinia carotovora | Bacterial spot and soft rot in tomato | [111] |
| CS-NPs | Ralstonia solanacearum | Bacterial wilt of tomato | [112] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chandrasekaran, M.; Kim, K.D.; Chun, S.C. Antibacterial Activity of Chitosan Nanoparticles: A Review. Processes 2020, 8, 1173. https://doi.org/10.3390/pr8091173
Chandrasekaran M, Kim KD, Chun SC. Antibacterial Activity of Chitosan Nanoparticles: A Review. Processes. 2020; 8(9):1173. https://doi.org/10.3390/pr8091173
Chicago/Turabian StyleChandrasekaran, Murugesan, Ki Deok Kim, and Se Chul Chun. 2020. "Antibacterial Activity of Chitosan Nanoparticles: A Review" Processes 8, no. 9: 1173. https://doi.org/10.3390/pr8091173
APA StyleChandrasekaran, M., Kim, K. D., & Chun, S. C. (2020). Antibacterial Activity of Chitosan Nanoparticles: A Review. Processes, 8(9), 1173. https://doi.org/10.3390/pr8091173