
The Effect of Paraburkholderia on the Nutrients in Eutrophic Lakes

Abstract
:1. Introduction
2. Material and Methods
2.1. Bacteria and Cyanobacteria
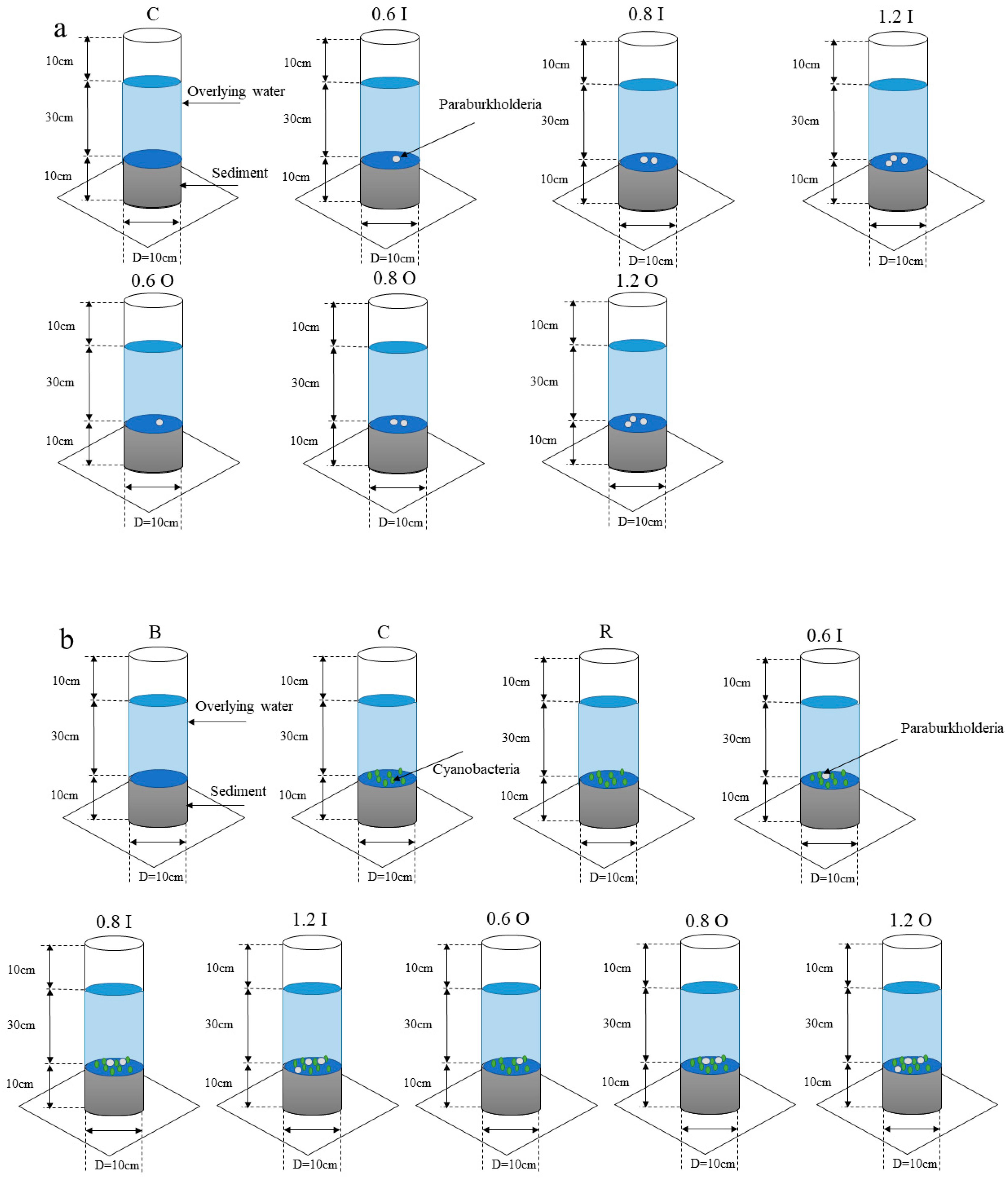
2.2. Constructing an SWI with Different Concentrations of Bacteria
2.3. Construction of the Bacteria–Algae System at the SWI
2.4. Analytical Methods
3. Results and Discussion
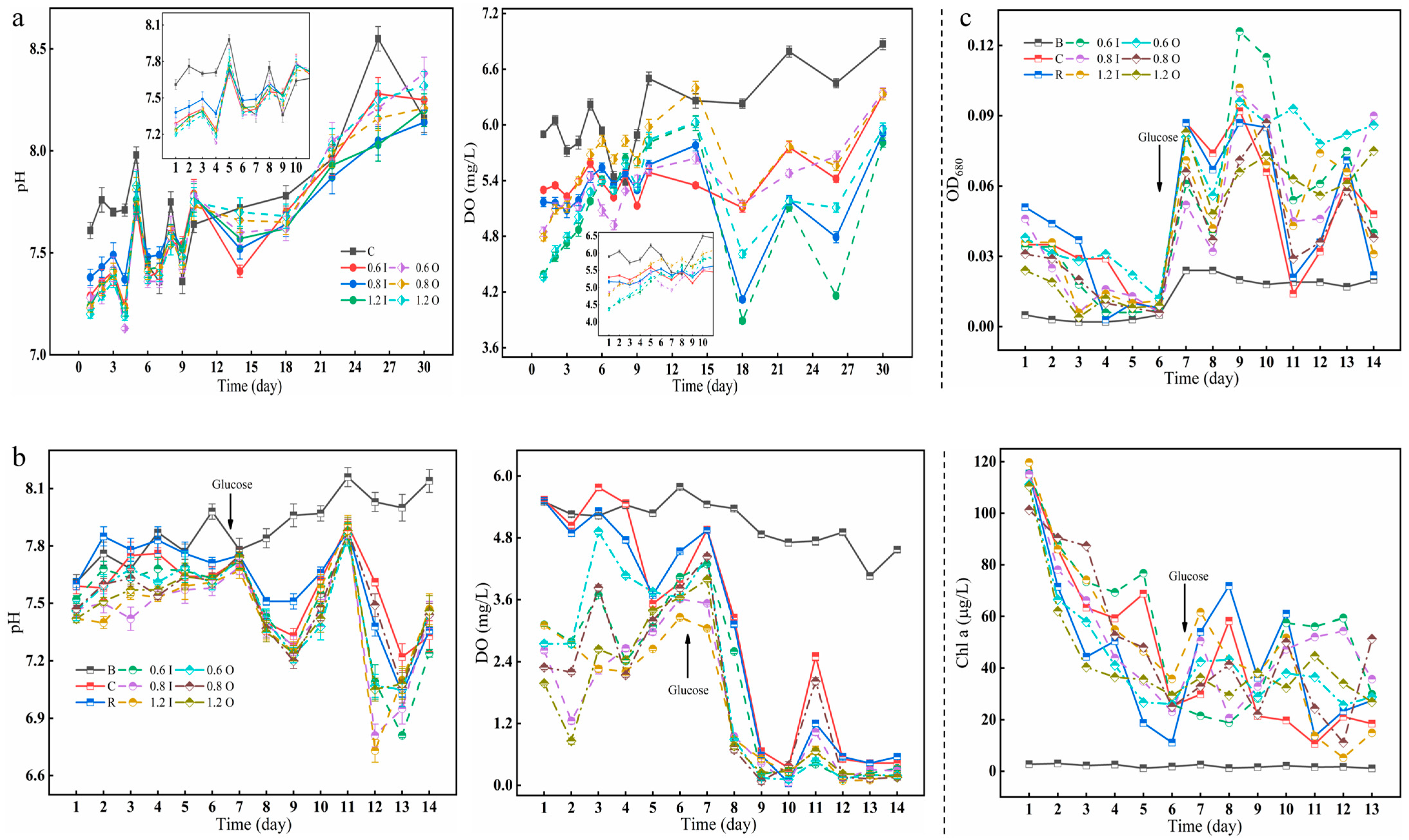
3.1. Changes in Overlying Water pH, DO, and Cyanobacterial Growth
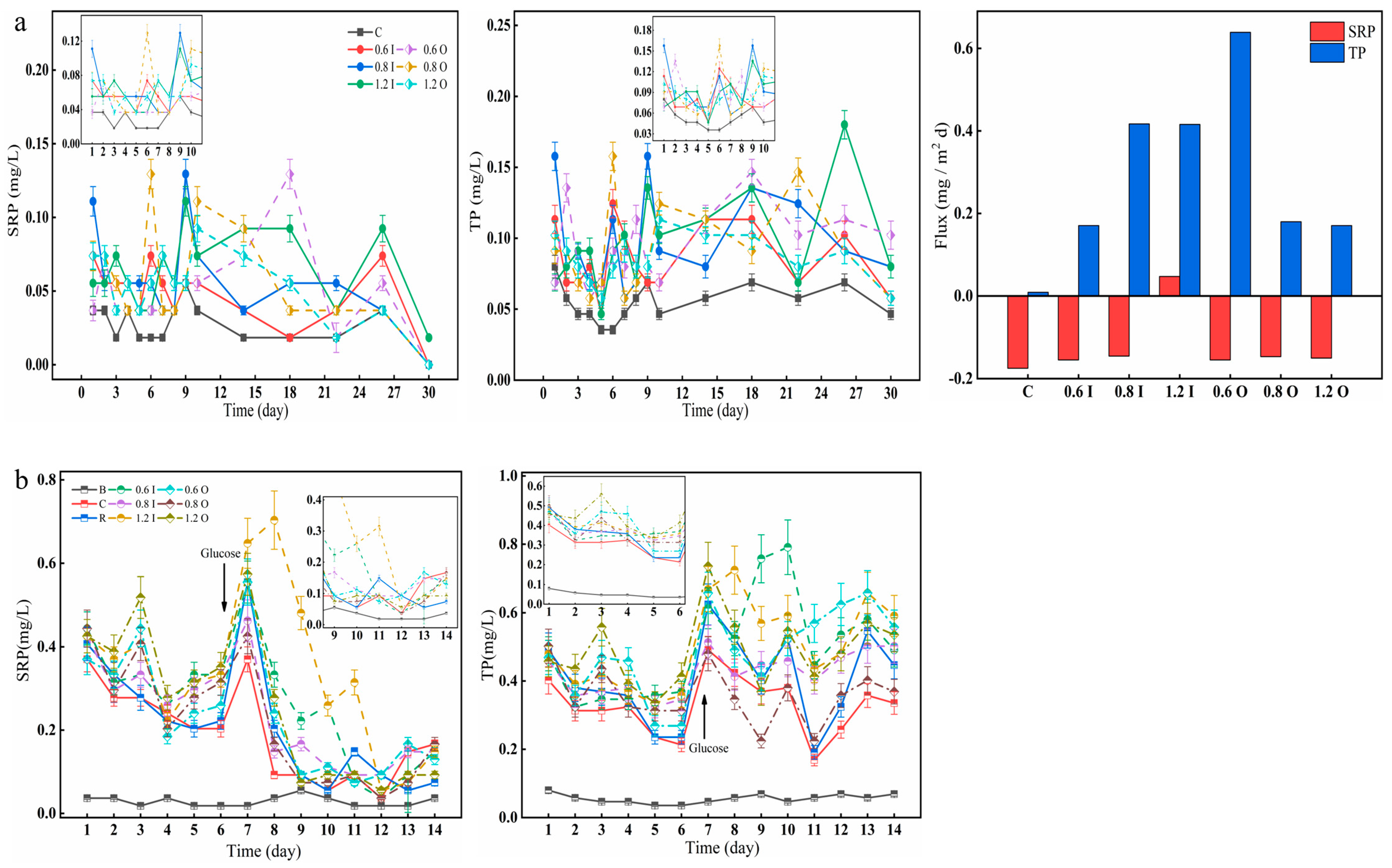
3.2. Phosphorus Performance in the Overlying Water
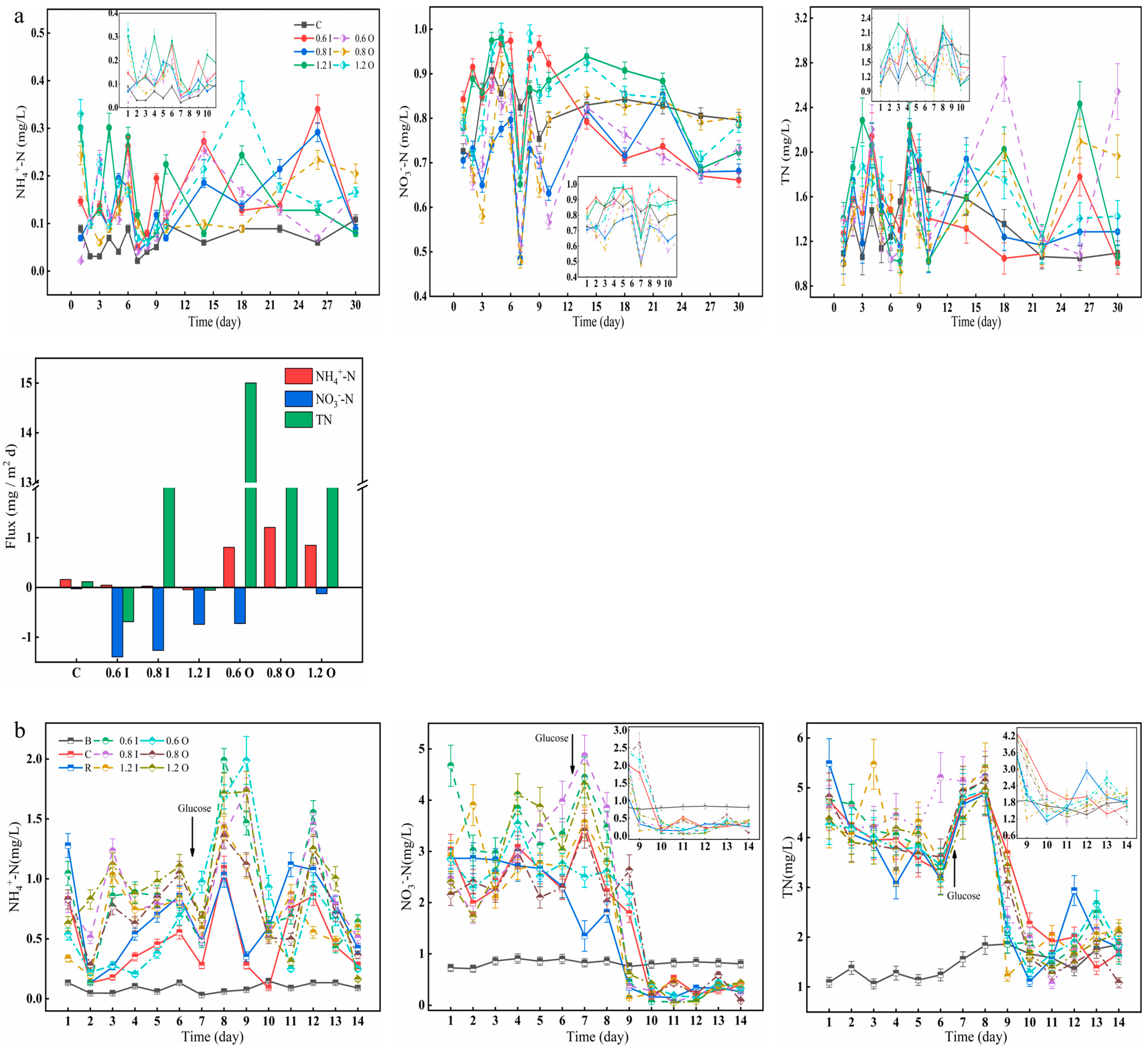
3.3. Nitrogen Concentration in the Overlying Water
3.4. Iron Content of the Overlying Water
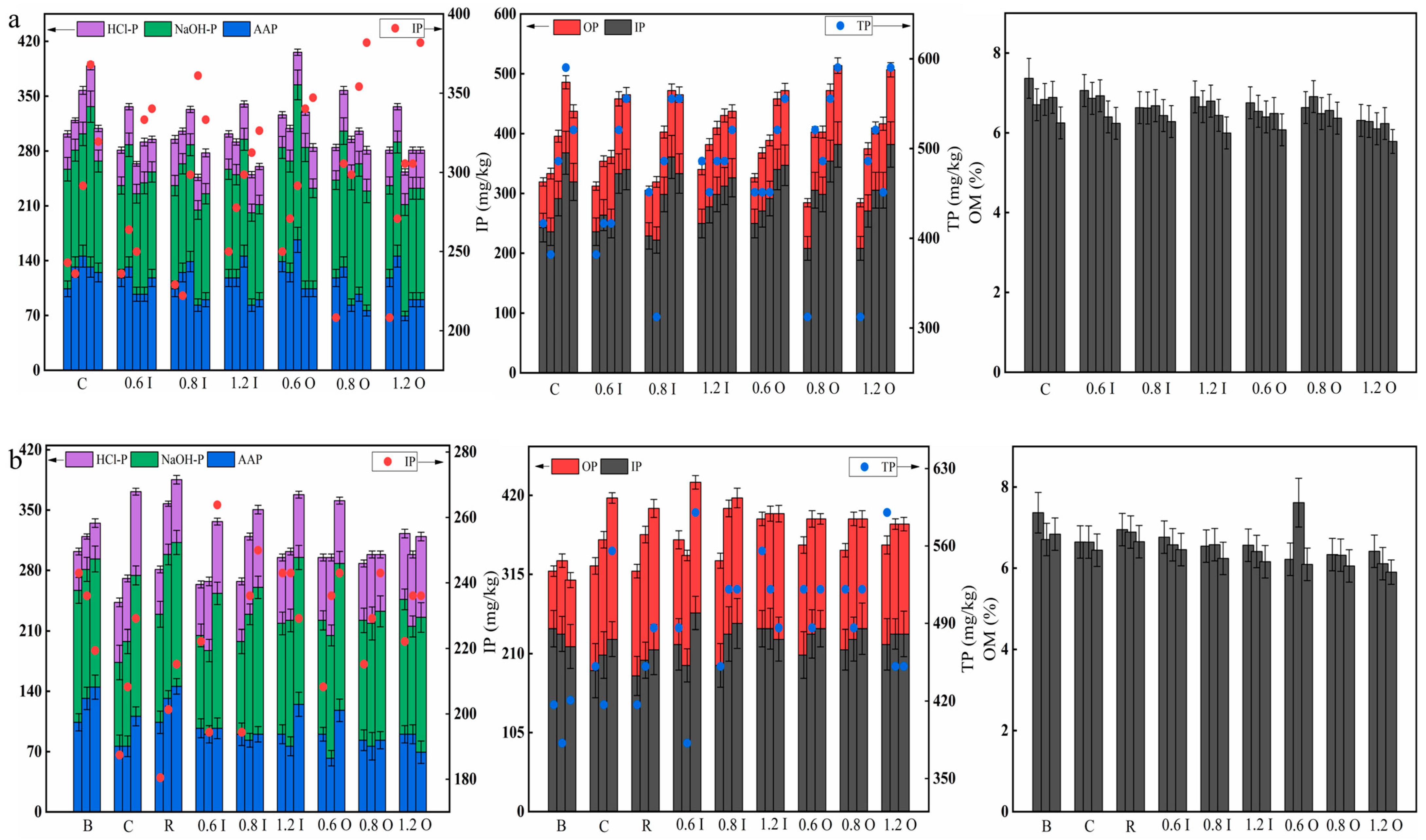
3.5. Variations in the Phosphorus Fractions and OM in the Sediments
3.6. Analysis of the Linkage between the Relevant Parameters of the Overlying Water and Phosphorus in the Sediment
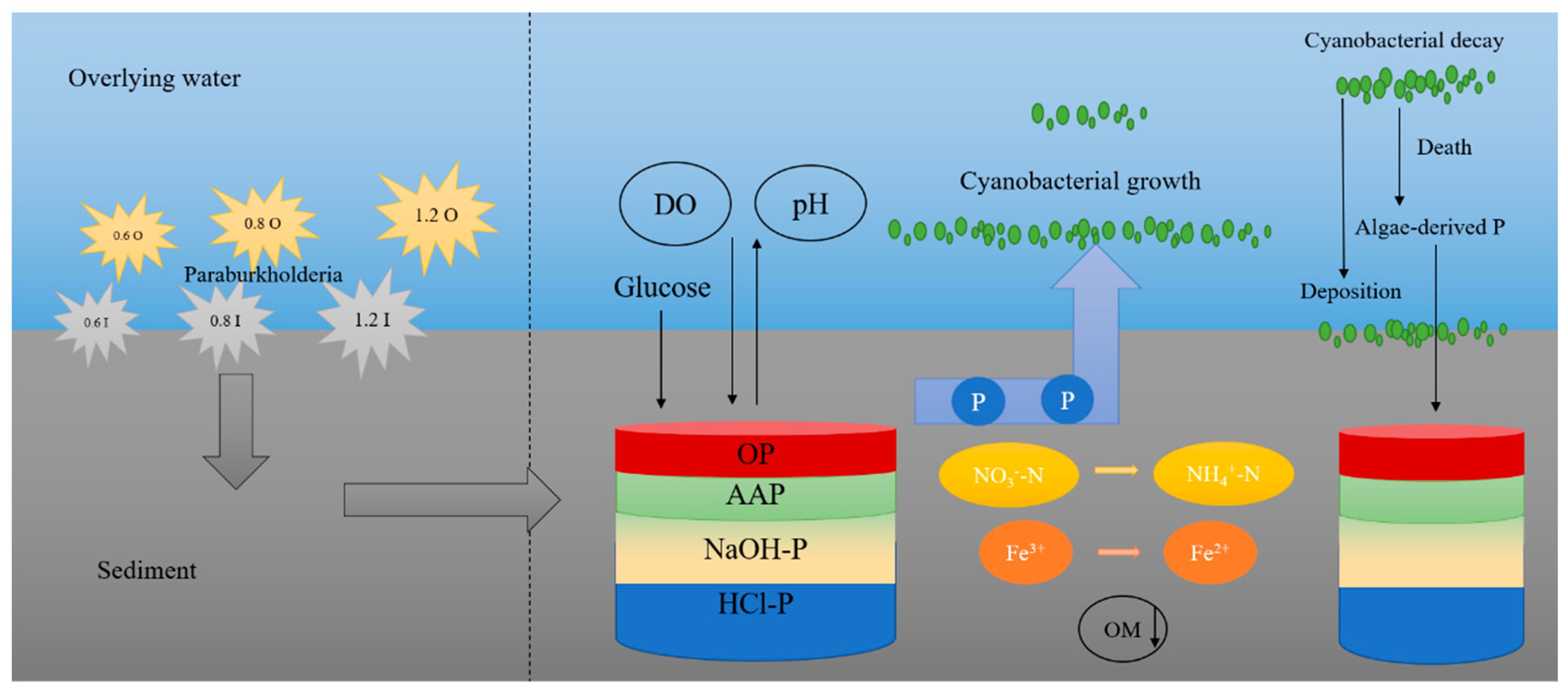
3.7. Analyzing the Potential Function of Paraburkholderia
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tanvir, R.U.; Hu, Z.; Zhang, Y.; Lu, J. Cyanobacterial community succession and associated cyanotoxin production in hypereutrophic and eutrophic freshwaters. Environ. Pollut. 2021, 290, 118056. [Google Scholar] [CrossRef]
- Berry, M.A.; Davis, T.W.; Cory, R.M.; Duhaime, M.B.; Johengen, T.H.; Kling, G.W.; Marino, J.A.; Den Uyl, P.A.; Gossiaux, D.; Dick, G.J.; et al. Cyanobacterial harmful algal blooms are a biological disturbance to Western Lake Erie bacterial communities. Environ. Microbiol. 2017, 19, 1149–1162. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, L.; Hou, J.; Wei, Q.; Fu, F.; Shao, H. Decomposition of macroalgal blooms influences phosphorus release from the sediments and implications for coastal restoration in Swan Lake, Shandong, China. Ecol. Eng. 2013, 60, 19–28. [Google Scholar] [CrossRef]
- Dai, J.; Chen, D.; Wu, S.; Wu, X.; Gao, G.; Tang, X.; Shao, K.; Lv, X.; Xue, W.; Yang, Q. Dynamics of phosphorus and bacterial phoX genes during the decomposition of Microcystis blooms in a mesocosm. PLoS ONE 2018, 13, e0195205. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, J.; Chen, X.; Jiang, Q.; Liu, Y.; Xie, S. Cyanobacterial bloom induces structural and functional succession of microbial communities in eutrophic lake sediments. Environ. Pollut. 2021, 284, 117157. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.Q.; Jin, Z.H.; Che, F.F.; Cao, X.; Song, X.S.; Lu, C.Y.; Huang, W. Characterization of phosphorus sorption and microbial community in lake sediments during overwinter and recruitment periods of cyanobacteria. Chemosphere 2022, 307, 135777. [Google Scholar] [CrossRef]
- Schindler, D.W.; Hecky, R.; Findlay, D.; Stainton, M.; Parker, B.; Paterson, M.; Beaty, K.; Lyng, M.; Kasian, S. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [PubMed]
- Xie, E.; Su, Y.; Deng, S.; Kontopyrgou, M.; Zhang, D. Significant influence of phosphorus resources on the growth and alkaline phosphatase activities of Microcystis aeruginosa. Environ. Pollut. 2021, 268, 115807. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Shen, Q.; Liu, Y.; Liu, J. Mobility of different phosphorus pools in the sediment of Lake Dianchi during cyanobacterial blooms. Environ. Monit. Assess. 2007, 132, 141–153. [Google Scholar] [CrossRef]
- Yao, L.; Zhao, X.; Zhou, G.-J.; Liang, R.; Gou, T.; Xia, B.; Li, S.; Liu, C. Seasonal succession of phytoplankton functional groups and driving factors of cyanobacterial blooms in a subtropical reservoir in South China. Water 2020, 12, 1167. [Google Scholar] [CrossRef]
- Zhu, M.; Zhu, G.; Li, W.; Zhang, Y.; Zhao, L.; Gu, Z. Estimation of the algal-available phosphorus pool in sediments of a large, shallow eutrophic lake (Taihu, China) using profiled SMT fractional analysis. Environ. Pollut. 2013, 173, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Dolman, A.M.; Rücker, J.; Pick, F.R.; Fastner, J.; Rohrlack, T.; Mischke, U.; Wiedner, C. Cyanobacteria and cyanotoxins: The influence of nitrogen versus phosphorus. PLoS ONE 2012, 7, e38757. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Gibson, G.; Carlson, R.; Simpson, J.; Smeltzer, E.; Gerritson, J.; Chapra, S.; Heiskary, S.; Jones, J.; Kennedy, R. Nutrient Criteria Technical Guidance Manual: Lakes and Reservoirs (EPA-822-B-00-001); United States Environment Protection Agency: Washington, DC, USA, 2000. [Google Scholar]
- Grant, W.D.; Long, P.E. Environmental Microbiology; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Xie, L.; Xie, P.; Tang, H. Enhancement of dissolved phosphorus release from sediment to lake water by Microcystis blooms—An enclosure experiment in a hyper-eutrophic, subtropical Chinese lake. Environ. Pollut. 2003, 122, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Li, Y.; He, C.; Acharya, K. Dynamic behavior of sediment resuspension and nutrients release in the shallow and wind-exposed Meiliang Bay of Lake Taihu. Sci. Total Environ. 2020, 708, 135131. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wang, Y.Q.; He, J.; Luo, X.Z.; Zheng, Z. Phosphorus mobility among sediments, water and cyanobacteria enhanced by cyanobacteria blooms in eutrophic Lake Dianchi. Environ. Pollut. 2016, 219, 580–587. [Google Scholar] [CrossRef]
- Jalali, M.; Peikam, E.N. Phosphorus sorption-desorption behaviour of river bed sediments in the Abshineh river, Hamedan, Iran, related to their composition. Environ. Monit. Assess. 2013, 185, 537–552. [Google Scholar] [CrossRef]
- Burkholder, W.H. Three bacterial plant pathogens: Phytomonas earyophylli sp. n., Phytomonas alliicola sp. n., and Phytomonas manihotis (Arthaud-Berthet et Sondar) Viégas. Phytopathology 1942, 32, 141–149. [Google Scholar]
- Yabuuchi, E.; Kosako, Y.; Oyaizu, H.; Yano, I.; Hotta, H.; Hashimoto, Y.; Ezaki, T.; Arakawa, M. Proposal of Burkholderia gen. nov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes 1981) comb. nov. Microbiol. Immunol. 1992, 36, 1251–1275. [Google Scholar] [CrossRef]
- Coenye, T.; Vandamme, P. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ. Microbiol. 2003, 5, 719–729. [Google Scholar] [CrossRef]
- Estrada-De Los Santos, P.; Bustillos-Cristales, R.O.; Caballero-Mellado, J. Burkholderia, a genus rich in plant-associated nitrogen fixers with wide environmental and geographic distribution. Appl. Environ. Microbiol. 2001, 67, 2790–2798. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Aguilar, L.; Díaz, R.; Pena-Cabriales, J.J.; Estrada-de Los Santos, P.; Dunn, M.F.; Caballero-Mellado, J. Multichromosomal genome structure and confirmation of diazotrophy in novel plant-associated Burkholderia species. Appl. Environ. Microbiol. 2008, 74, 4574–4579. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Mellado, J.; Onofre-Lemus, J.; Estrada-De Los Santos, P.; Martínez-Aguilar, L. The tomato rhizosphere, an environment rich in nitrogen-fixing Burkholderia species with capabilities of interest for agriculture and bioremediation. Appl. Environ. Microbiol. 2007, 73, 5308–5319. [Google Scholar] [CrossRef] [PubMed]
- Peix, A.; Mateos, P.; Rodriguez-Barrueco, C.; Martinez-Molina, E.; Velazquez, E. Growth promotion of common bean (Phaseolus vulgaris L.) by a strain of Burkholderia cepacia under growth chamber conditions. Soil Biol. Biochem. 2001, 33, 1927–1935. [Google Scholar] [CrossRef]
- Ruban, V.; López-Sánchez, J.; Pardo, P.; Rauret, G.; Muntau, H.; Quevauviller, P. Harmonized protocol and certified reference material for the determination of extractable contents of phosphorus in freshwater sediments—A synthesis of recent works. Fresenius’ J. Anal. Chem. 2001, 370, 224–228. [Google Scholar] [CrossRef]
- Younis, A.M.; Soliman, N.F.; Elkady, E.M.; Mohamedein, L.I. Distribution and ecological risk evaluation of bioavailable phosphorus in sediments of El Temsah Lake, Suez Canal. Oceanologia 2022, 64, 287–298. [Google Scholar] [CrossRef]
- Huang, W.; Lu, Y.; Li, J.H.; Zheng, Z.; Zhang, J.B.; Jiang, X. Effect of ionic strength on phosphorus sorption in different sediments from a eutrophic plateau lake. Rsc Adv. 2015, 5, 79607–79615. [Google Scholar] [CrossRef]
- Hu, M.; Sardans, J.; Le, Y.; Yan, R.; Zhong, Y.; Huang, J.; Peñuelas, J.; Tong, C. Biogeochemical behavior of P in the soil and porewater of a low-salinity estuarine wetland: Availability, diffusion kinetics, and mobilization mechanism. Water Res. 2022, 219, 118617. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Cao, X.Y.; Li, H.; Zhou, Z.J.; Wang, S.Y.; Wang, Z.C.; Song, C.L.; Zhou, Y.Y. Distribution of phosphorus-solubilizing bacteria in relation to fractionation and sorption behaviors of phosphorus in sediment of the Three Gorges Reservoir. Environ. Sci. Pollut. Res. 2017, 24, 17679–17687. [Google Scholar] [CrossRef]
- Teng, Z.; Shao, W.; Zhang, K.; Huo, Y.; Li, M. Characterization of phosphate solubilizing bacteria isolated from heavy metal contaminated soils and their potential for lead immobilization. J. Environ. Manag. 2019, 231, 189–197. [Google Scholar] [CrossRef]
- Chelsky, A.; Pitt, K.A.; Ferguson, A.J.; Bennett, W.W.; Teasdale, P.R.; Welsh, D.T. Decomposition of jellyfish carrion in situ: Short-term impacts on infauna, benthic nutrient fluxes and sediment redox conditions. Sci. Total Environ. 2016, 566, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Lin, H.; Bartlett, S.L.; Houghton, E.M.; Robertson, D.M.; Guo, L. Partitioning and transformation of organic and inorganic phosphorus among dissolved, colloidal and particulate phases in a hypereutrophic freshwater estuary. Water Res. 2021, 196, 117025. [Google Scholar] [CrossRef]
- Olguin, E.; Hernández, B.; Araus, A.; Camacho, R.; González, R.; Ramírez, M.; Galicia, S.; Mercado, G. Simultaneous high-biomass protein production and nutrient removal using Spirulina maxima in sea water supplemented with anaerobic effluents. World J. Microbiol. Biotechnol. 1994, 10, 576–578. [Google Scholar] [CrossRef] [PubMed]
- Van Baalen, C.; Hoare, D.S.; Brandt, E. Heterotrophic growth of blue-green algae in dim light. J. Bacteriol. 1971, 105, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Lewis Jr, W.M.; Wurtsbaugh, W.A.; Paerl, H.W. Rationale for control of anthropogenic nitrogen and phosphorus to reduce eutrophication of inland waters. Environ. Sci. Technol. 2011, 45, 10300–10305. [Google Scholar] [CrossRef] [PubMed]
- Sarneel, J.; Geurts, J.; Beltman, B.; Lamers, L.; Nijzink, M.; Soons, M.; Verhoeven, J. The effect of nutrient enrichment of either the bank or the surface water on shoreline vegetation and decomposition. Ecosystems 2010, 13, 1275–1286. [Google Scholar] [CrossRef]
- Zhu, M.; Zhu, G.; Zhao, L.; Yao, X.; Zhang, Y.; Gao, G.; Qin, B. Influence of algal bloom degradation on nutrient release at the sediment–water interface in Lake Taihu, China. Environ. Sci. Pollut. Res. 2013, 20, 1803–1811. [Google Scholar] [CrossRef]
- Capone, D.G.; Carpenter, E.J. Nitrogen fixation in the marine environment. Science 1982, 217, 1140–1142. [Google Scholar] [CrossRef]
- Gao, Y.; Cornwell, J.C.; Stoecker, D.K.; Owens, M.S. Influence of cyanobacteria blooms on sediment biogeochemistry and nutrient fluxes. Limnol. Oceanogr. 2014, 59, 959–971. [Google Scholar] [CrossRef]
- Brunberg, A.-K. Contribution of bacteria in the mucilage of Microcystis spp.(Cyanobacteria) to benthic and pelagic bacterial production in a hypereutrophic lake. FEMS Microbiol. Ecol. 1999, 29, 13–22. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, S.; Yu, M.; Zhang, Z.; Wang, X.; Zhang, S.; Wang, G. Seasonal iron-sulfur interactions and the stimulated phosphorus mobilization in freshwater lake sediments. Sci. Total Environ. 2021, 768, 144336. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, Y.; Bai, X.; Li, W. Effect of algal blooms outbreak and decline on phosphorus migration in Lake Taihu, China. Environ. Pollut. 2022, 296, 118761. [Google Scholar] [CrossRef] [PubMed]
- Kalscheur, K.N.; Rojas, M.; Peterson, C.G.; Kelly, J.J.; Gray, K.A. Algal exudates and stream organic matter influence the structure and function of denitrifying bacterial communities. Microb. Ecol. 2012, 64, 881–892. [Google Scholar] [CrossRef]
- Han, C.; Ding, S.; Yao, L.; Shen, Q.; Zhu, C.; Wang, Y.; Xu, D. Dynamics of phosphorus–iron–sulfur at the sediment–water interface influenced by algae blooms decomposition. J. Hazard. Mater. 2015, 300, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.D.A.; España, M.; Aguirre, C.; Kojima, K.; Ohkama-Ohtsu, N.; Sekimoto, H.; Yokoyama, T. Burkholderia and Paraburkholderia are predominant soybean rhizobial genera in Venezuelan soils in different climatic and topographical regions. Microbes Environ. 2019, 34, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Nürnberg, G.K.; Molot, L.A.; O’Connor, E.; Jarjanazi, H.; Winter, J.; Young, J. Evidence for internal phosphorus loading, hypoxia and effects on phytoplankton in partially polymictic Lake Simcoe, Ontario. J. Great Lakes Res. 2013, 39, 259–270. [Google Scholar] [CrossRef]
- Markovic, S.; Liang, A.; Watson, S.B.; Guo, J.; Mugalingam, S.; Arhonditsis, G.; Morley, A.; Dittrich, M. Biogeochemical mechanisms controlling phosphorus diagenesis and internal loading in a remediated hard water eutrophic embayment. Chem. Geol. 2019, 514, 122–137. [Google Scholar] [CrossRef]
- Norton, S.A.; Coolidge, K.; Amirbahman, A.; Bouchard, R.; Kopacek, J.; Reinhardt, R. Speciation of Al, Fe, and P in recent sediment from three lakes in Maine, USA. Sci. Total Environ. 2008, 404, 276–283. [Google Scholar] [CrossRef]
- Jan, J.; Borovec, J.; Kopacek, J.; Hejzlar, J. What do results of common sequential fractionation and single-step extractions tell us about P binding with Fe and Al compounds in non-calcareous sediments? Water Res. 2013, 47, 547–557. [Google Scholar] [CrossRef]
- Worsfold, P.J.; Monbet, P.; Tappin, A.D.; Fitzsimons, M.F.; Stiles, D.A.; McKelvie, I.D. Characterisation and quantification of organic phosphorus and organic nitrogen components in aquatic systems: A review. Anal Chim Acta 2008, 624, 37–58. [Google Scholar] [CrossRef]
- Venkatesan, A.K.; Gan, W.; Ashani, H.; Herckes, P.; Westerhoff, P. Size exclusion chromatography with online ICP-MS enables molecular weight fractionation of dissolved phosphorus species in water samples. Water Res. 2018, 133, 264–271. [Google Scholar] [CrossRef]
- Ding, S.; Sun, Q.; Xu, D.; Jia, F.; He, X.; Zhang, C. High-resolution simultaneous measurements of dissolved reactive phosphorus and dissolved sulfide: The first observation of their simultaneous release in sediments. Environ. Sci. Technol. 2012, 46, 8297–8304. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, H.M.; Ferreira, T.O.; Barcellos, D.; Nóbrega, G.N.; Antelo, J.; Otero, X.L.; Bernardino, A.F. From sinks to sources: The role of Fe oxyhydroxide transformations on phosphorus dynamics in estuarine soils. J. Environ. Manage. 2021, 278, 111575. [Google Scholar] [CrossRef] [PubMed]
- Jordan, T.E.; Cornwell, J.C.; Boynton, W.R.; Anderson, J.T. Changes in phosphorus biogeochemistry along an estuarine salinity gradient: The iron conveyer belt. Limnol. Oceanogr. 2008, 53, 172–184. [Google Scholar] [CrossRef]
- Tripura, C.; Sashidhar, B.; Podile, A.R. Ethyl methanesulfonate mutagenesis–enhanced mineral phosphate solubilization by groundnut-associated Serratia marcescens GPS-5. Curr. Microbiol. 2007, 54, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Stephen, J.; Jisha, M. Gluconic acid production as the principal mechanism of mineral phosphate solubilization by Burkholderia sp.(MTCC 8369). J. Trop. Agric. 2011, 49, 99–103. [Google Scholar]
- Lin, T.-F.; Huang, H.-I.; Shen, F.-T.; Young, C.-C. The protons of gluconic acid are the major factor responsible for the dissolution of tricalcium phosphate by Burkholderia cepacia CC-Al74. Bioresour. Technol. 2006, 97, 957–960. [Google Scholar] [CrossRef] [PubMed]
- Song, O.-R.; Lee, S.-J.; Lee, Y.-S.; Lee, S.-C.; Kim, K.-K.; Choi, Y.-L. Solubilization of insoluble inorganic phosphate by Burkholderia cepacia DA23 isolated from cultivated soil. Braz. J. Microbiol. 2008, 39, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Maar, M.; Timmermann, K.; Petersen, J.K.; Gustafsson, K.E.; Storm, L.M. A model study of the regulation of blue mussels by nutrient loadings and water column stability in a shallow estuary, the Limfjorden. J. Sea Res. 2010, 64, 322–333. [Google Scholar] [CrossRef]
- Nędzarek, A.; Tórz, A.; Rakusa-Suszczewski, S.; Bonisławska, M. Nitrogen and phosphorus release during fish decomposition and implications for the ecosystem of maritime Antarctica. Polar Biol. 2015, 38, 733–740. [Google Scholar] [CrossRef]
- Li, X.; Song, J.; Ma, Q.; Li, N.; Yuan, H.; Duan, L.; Qu, B. Experiments and evidences: Jellyfish (Nemopilema nomurai) decomposing and nutrients (nitrogen and phosphorus) released. Acta Oceanol. Sin. 2015, 34, 1–12. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, P.; Wang, C.; Hou, J.; Miao, L.; Yuan, Y.; Wang, T.; Liu, C. Assessment of mobilization of labile phosphorus and iron across sediment-water interface in a shallow lake (Hongze) based on in situ high-resolution measurement. Environ. Pollut. 2016, 219, 873–882. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phase I (bacteria only) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| TPs | NaOH-P | HCl-P | AAP | SRP | TPw | TN | Fe | ||
| TPs | 1 | 0.24 | 0.32 | −0.40 * | −0.30 | 0.21 | 0.08 | 0.41 * | |
| NaOH-P | 0.24 | 1 | −0.01 | 0.36 | 0.32 | 0.45 * | 0.04 | 0.17 | |
| HCl-P | 0.32 | −0.01 | 1 | −0.27 | −0.17 | −0.24 | 0.22 | 0.04 | |
| AAP | −0.40 * | 0.36 | −0.27 | 1 | 0.42 * | 0.05 | 0.06 | −0.35 | |
| SRP | −0.30 | 0.32 | −0.17 | 0.42 * | 1 | 0.52 ** | −0.11 | −0.39 * | |
| TPw | 0.21 | 0.45 * | −0.24 | 0.05 | 0.52 ** | 1 | −0.06 | 0.31 | |
| TN | 0.08 | 0.04 | 0.22 | 0.06 | −0.11 | −0.06 | 1 | −0.20 | |
| Fe | 0.41 * | 0.17 | 0.04 | −0.35 | −0.39 * | 0.31 | −0.20 | 1 | |
| Phase II (bacteria–algae) | |||||||||
| TPs | NaOH-P | HCl-P | AAP | Chl a | SRP | TPw | TN | Fe | |
| TPs | 1 | 0.55 * | −0.07 | 0.12 | −0.15 | 0.15 | −0.51 * | 0.20 | 0.02 |
| NaOH-P | 0.55 * | 1 | 0.34 | 0.21 | −0.58 * | −0.51 * | 0.06 | −0.39 | 0.58 * |
| HCl-P | −0.07 | 0.34 | 1 | −0.43 | −0.36 | −0.63 ** | 0.45 | −0.50 * | 0.34 |
| AAP | 0.12 | 0.21 | −0.43 | 1 | −0.31 | 0.12 | 0.13 | 0.15 | 0.16 |
| Chl a | −0.15 | −0.58 * | −0.36 | −0.31 | 1 | 0.43 | −0.09 | 0.11 | −0.33 |
| SRP | 0.15 | −0.51 * | −0.63 ** | 0.12 | 0.43 | 1 | −0.36 | 0.80 ** | −0.60 ** |
| TPw | −0.51 * | 0.06 | 0.45 | 0.13 | −0.09 | −0.36 | 1 | −0.64 ** | 0.46 |
| TN | 0.20 | −0.39 | −0.50 * | 0.15 | 0.11 | 0.80 ** | −0.64 ** | 1 | −0.73 ** |
| Fe | 0.02 | 0.58 * | 0.34 | 0.16 | −0.33 | −0.60 ** | 0.46 | −0.73 ** | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, C.; Chang, Y.; Dong, X.; Huang, W. The Effect of Paraburkholderia on the Nutrients in Eutrophic Lakes. Separations 2023, 10, 595. https://doi.org/10.3390/separations10120595
Tu C, Chang Y, Dong X, Huang W. The Effect of Paraburkholderia on the Nutrients in Eutrophic Lakes. Separations. 2023; 10(12):595. https://doi.org/10.3390/separations10120595
Chicago/Turabian StyleTu, Chengqi, Yongsheng Chang, Xiaoshuang Dong, and Wei Huang. 2023. "The Effect of Paraburkholderia on the Nutrients in Eutrophic Lakes" Separations 10, no. 12: 595. https://doi.org/10.3390/separations10120595