
Antioxidant and Hepatoprotective Activity of an Extract from the Overground Parts of Phlomis russeliana Lag. ex Benth
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
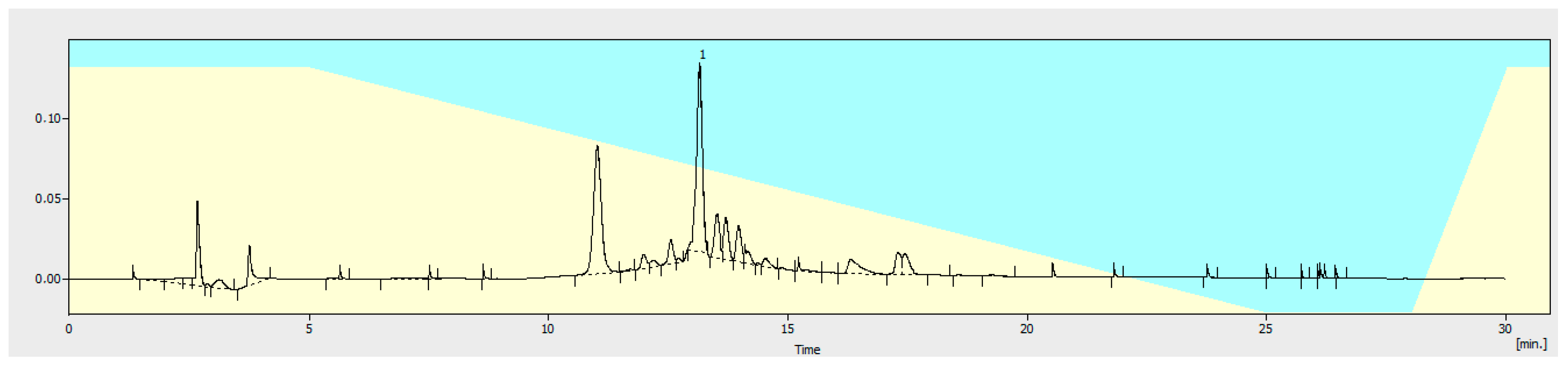
2.2. Extraction and Isolation of Verbascoside
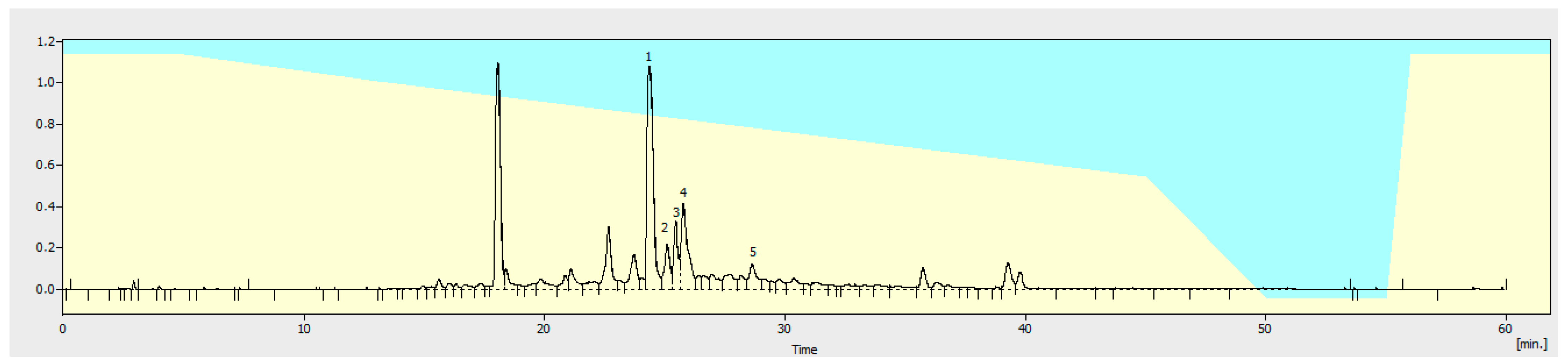
2.3. Phytochemical Analysis
2.4. Animals
2.5. In Vitro Experiments on Isolated Rat Hepatocytes
2.6. In Vivo Experimental Design
2.6.1. GSH and MDA Determination in Liver Tissue
2.6.2. Assay of Aspartate Aminotransferase (ASAT) and Alanine Aminotransferase (ALAT) in Blood Serum
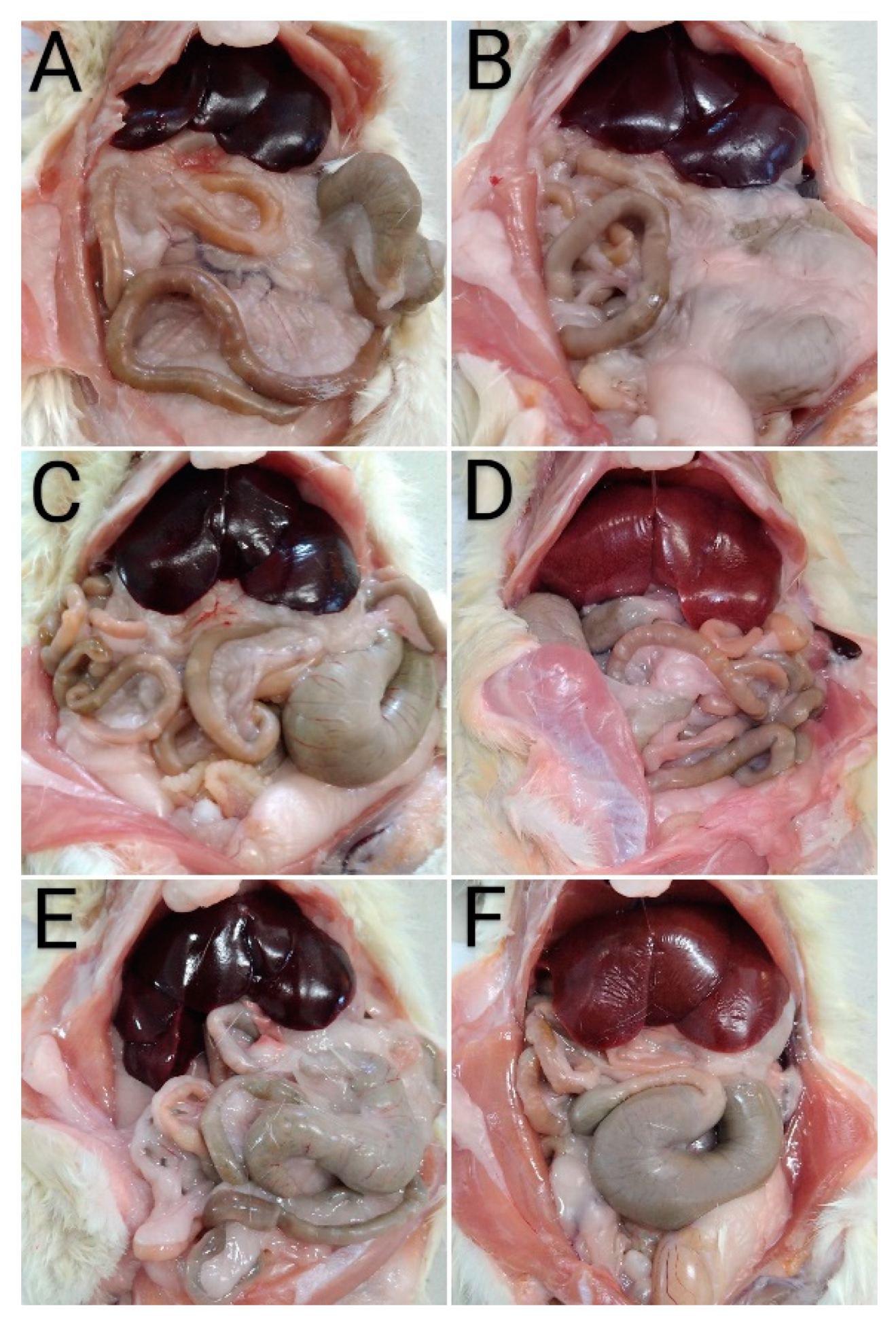
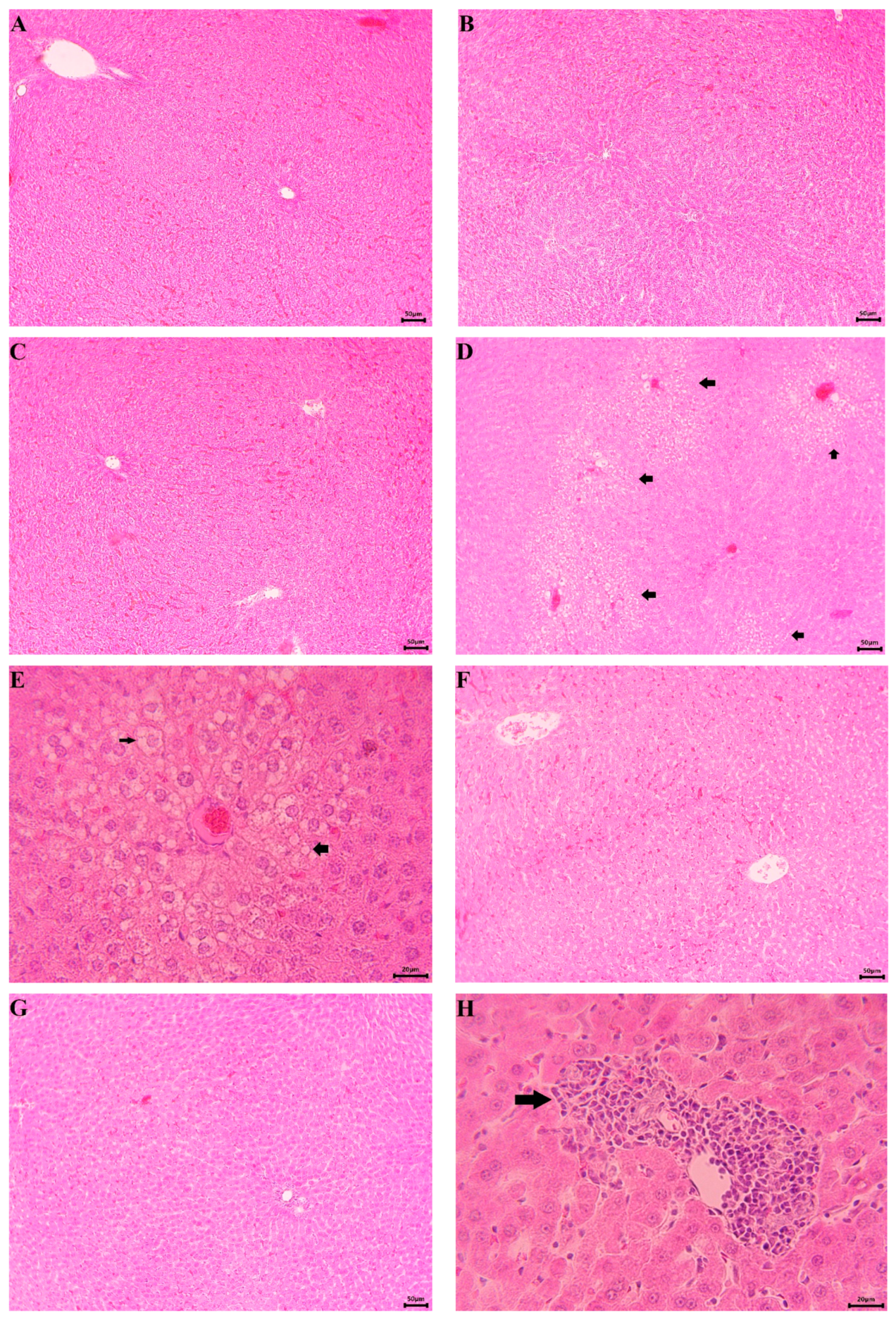
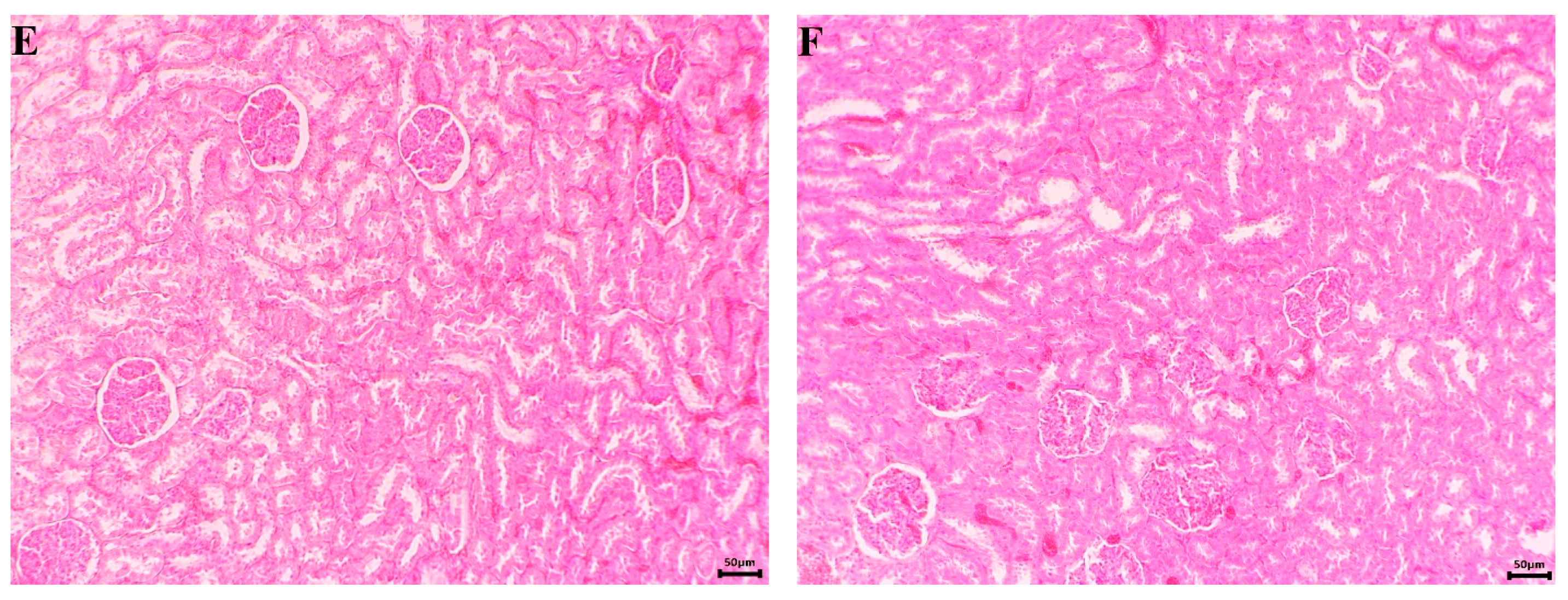
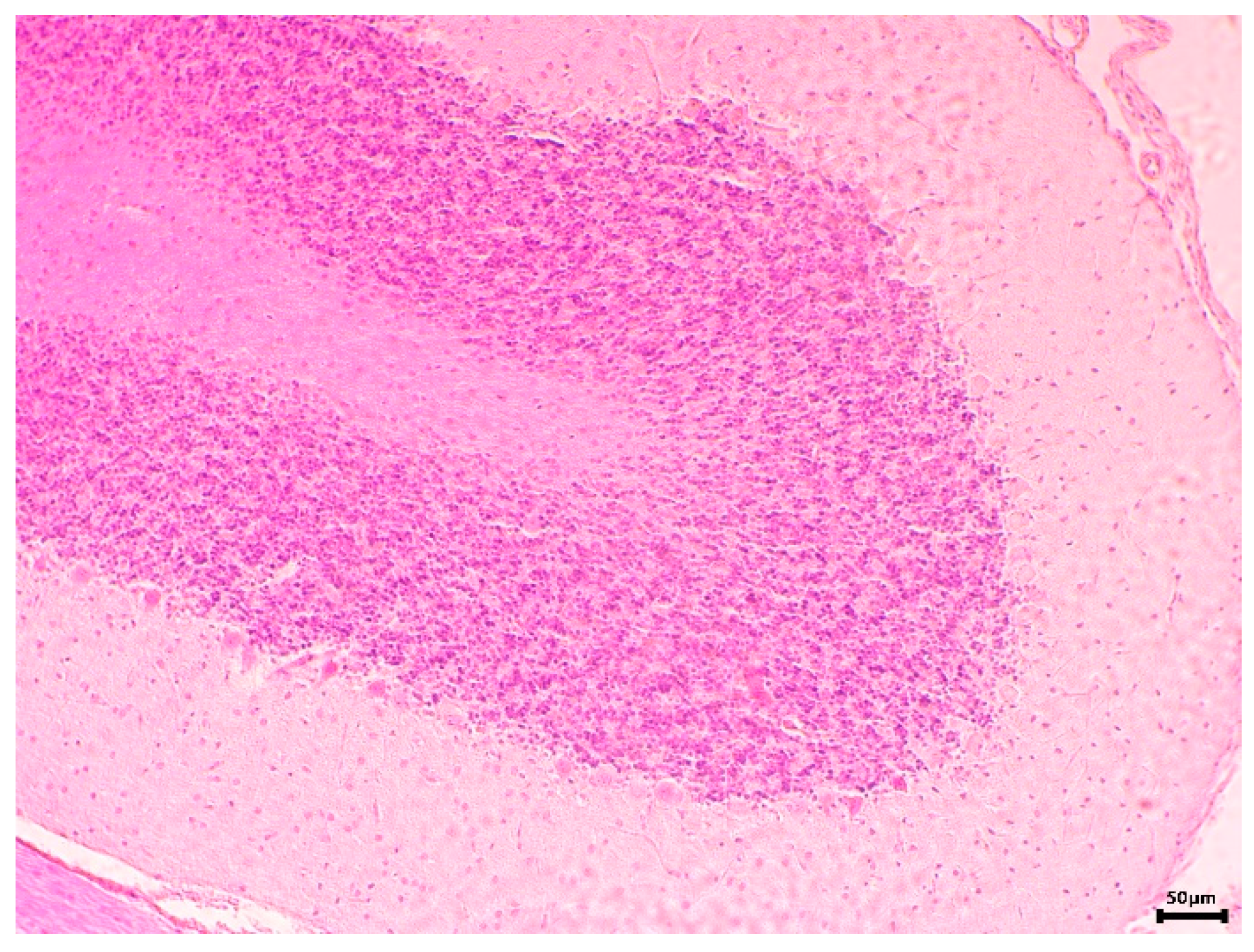
2.6.3. Histopathological Examination
2.7. Statistical Analysis
3. Results
3.1. Obtaining EPhR and Chemical Analysis
3.1.1. Flavonoid Identification in EPhR
3.1.2. Assay of Total Flavonoids
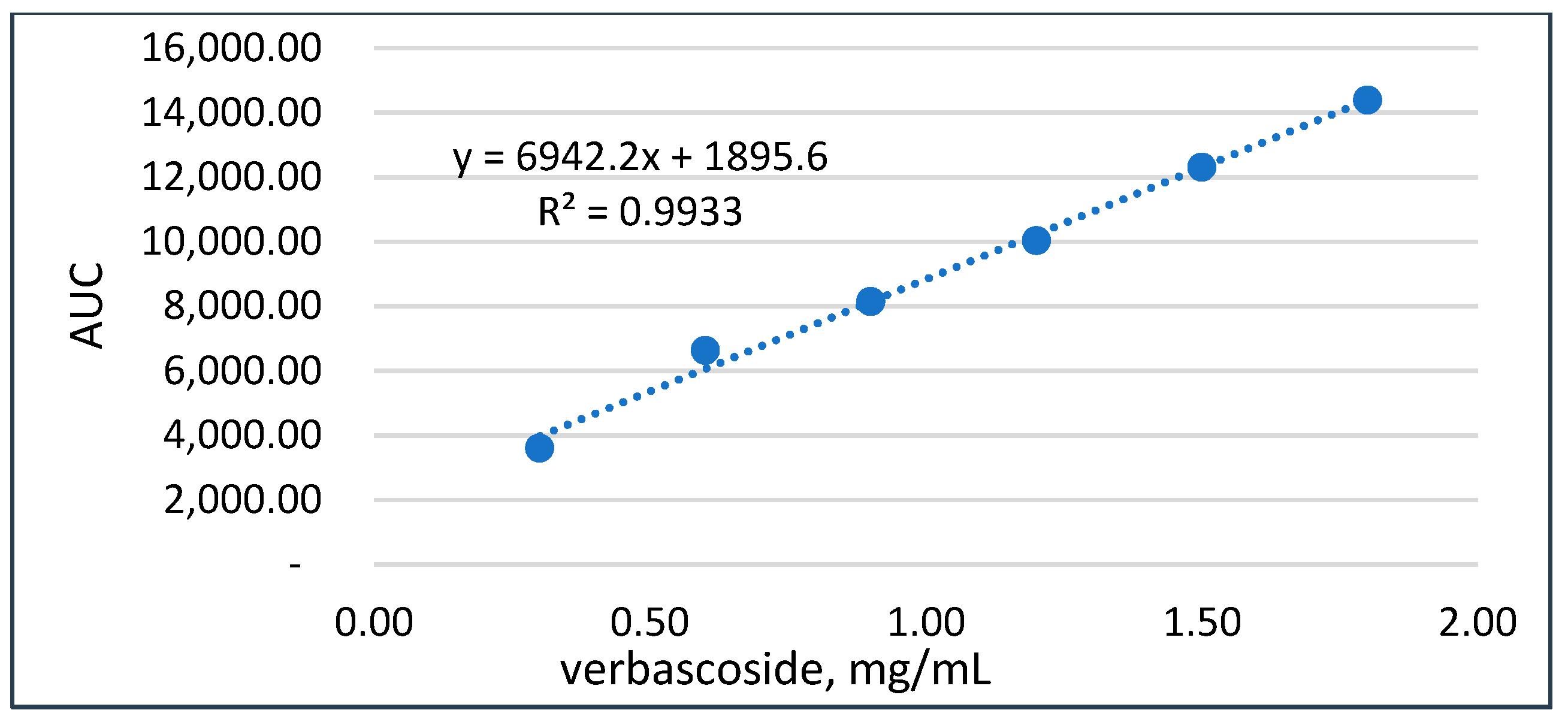
3.1.3. Quantitation of Verbascoside
3.1.4. Total Compounds Content
3.2. Evaluation of the Pharmacological Effects
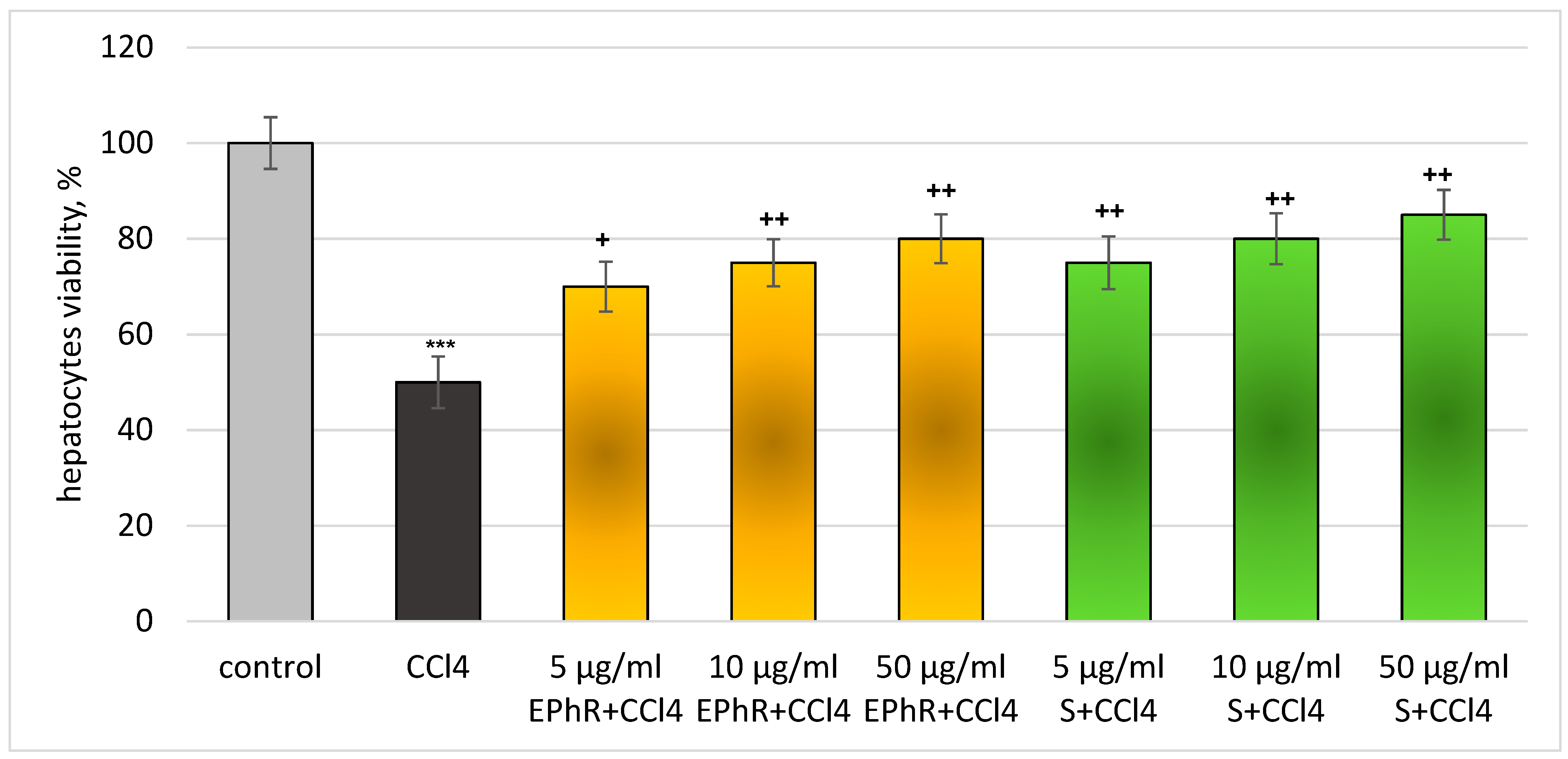
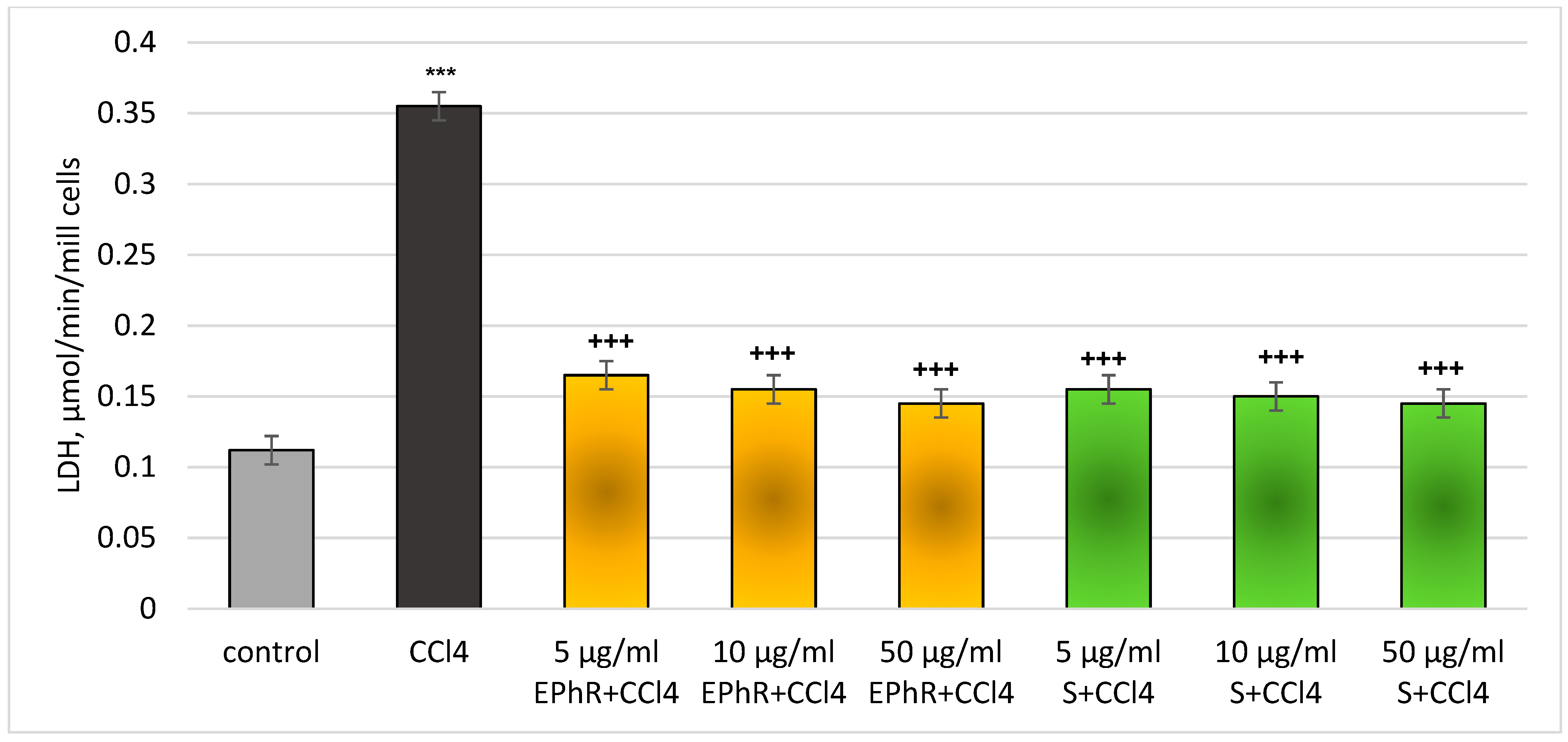
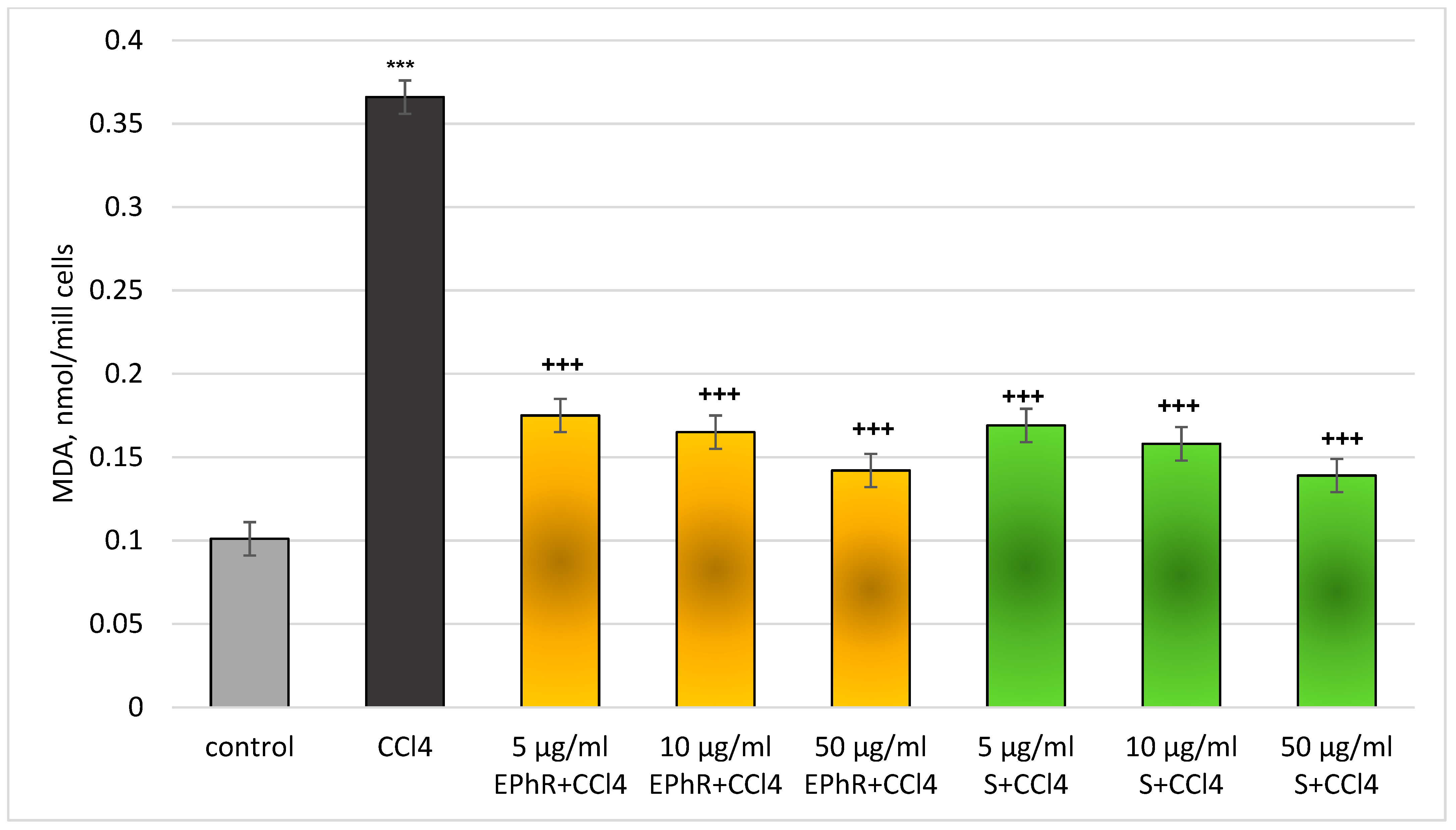
3.2.1. In Vitro Investigations
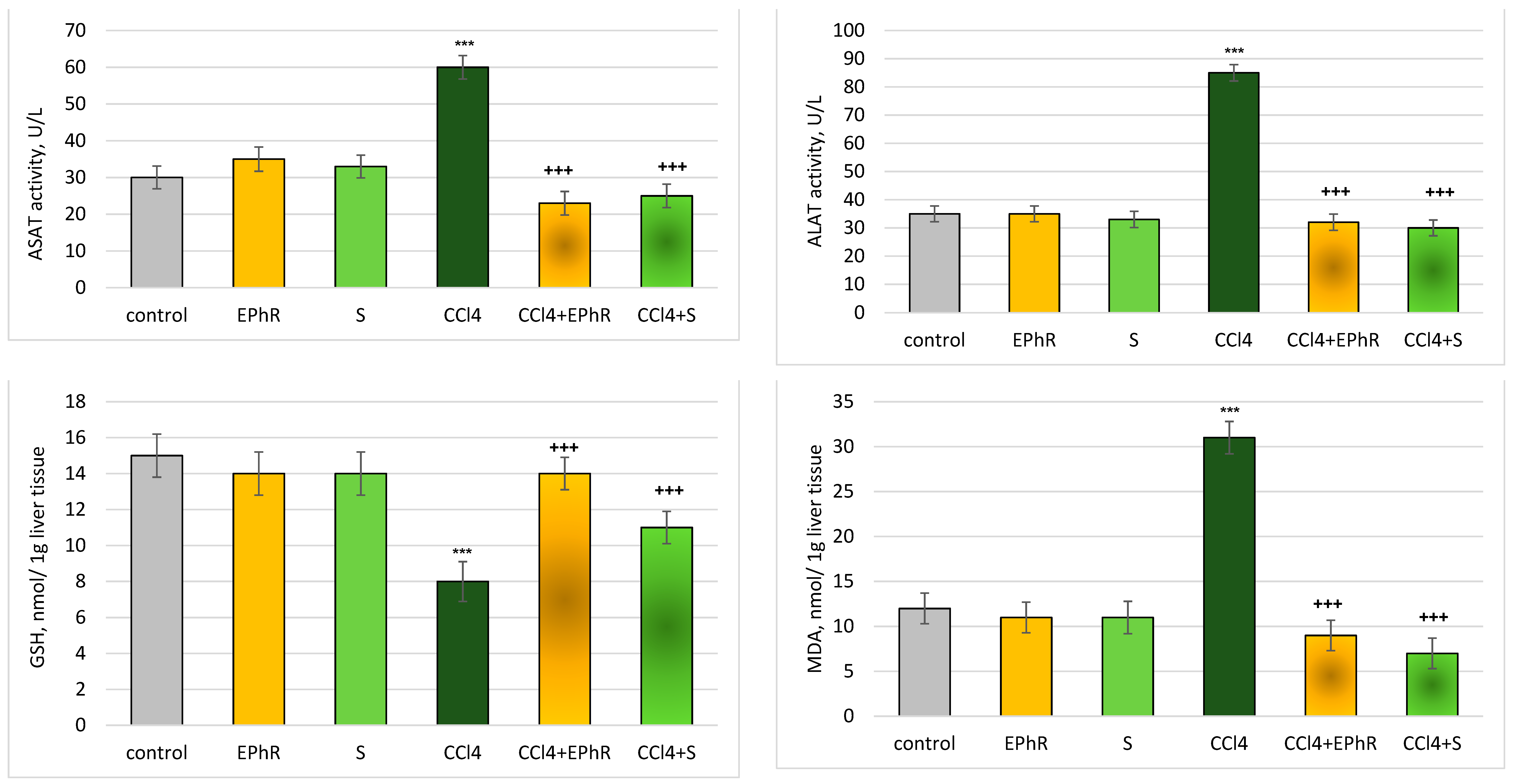
3.2.2. In Vivo Experiments with Old Wistar Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and Resupply of Pharmacologically Active Plant-Derived Natural Products: A Review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed]
- Recknagel, R.O.; Glende, E.A.; Dolak, J.A.; Waller, R.L. Mechanisms of Carbon Tetrachloride Toxicity. Pharmacol. Ther. 1989, 43, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Sarkhail, P.; Amin, G.; Surmaghi, M.H.S.; Shafiee, A. Composition of the Volatile Oils of Phlomis Lanceolata Boiss. & Hohen., Phlomis anisodonta Boiss. and Phlomis bruguieri Desf. from Iran. Flavour Fragr. J. 2005, 20, 327–329. [Google Scholar] [CrossRef]
- Couladis, M.; Tzakou, O.; Verykokidou, E.; Harvala, C. Screening of Some Greek Aromatic Plants for Antioxidant Activity. Phytother. Res. 2003, 17, 194–195. [Google Scholar] [CrossRef] [PubMed]
- Amor, I.L.-B.; Boubaker, J.; Sgaier, M.B.; Skandrani, I.; Bhouri, W.; Neffati, A.; Kilani, S.; Bouhlel, I.; Ghedira, K.; Chekir-Ghedira, L. Phytochemistry and Biological Activities of Phlomis Species. J. Ethnopharmacol. 2009, 125, 183–202. [Google Scholar] [CrossRef] [PubMed]
- Delazar, A.; Sabzevari, A.; Mojarrab, M.; Nazemiyeh, H.; Esnaashari, S.; Nahar, L.; Razavi, S.M.; Sarker, S.D. Free-Radical-Scavenging Principles from Phlomis Caucasica. J. Nat. Med. 2008, 62, 464–466. [Google Scholar] [CrossRef] [PubMed]
- Kondeva-Burdina, M.; Shkondrov, A.; Popov, G.; Manov, V.; Krasteva, I. In Vitro/In Vivo Hepatoprotective and Antioxidant Effects of Defatted Extract and a Phenolic Fraction Obtained from Phlomis Tuberosa. Int. J. Mol. Sci. 2023, 24, 10631. [Google Scholar] [CrossRef] [PubMed]
- Demirci, B.; Toyota, M.; Demirci, F.; Dadandi, M.Y.; Can Baser, K.H. Anticandidal Pimaradiene Diterpene from Phlomis Essential Oils. Comptes Rendus Chim. 2009, 12, 612–621. [Google Scholar] [CrossRef]
- Kırmızıbekmez, H.; Montoro, P.; Piacente, S.; Pizza, C.; Dönmez, A.; Çalış, İ. Identification by HPLC-PAD-MS and Quantification by HPLC-PAD of Phenylethanoid Glycosides of Five Phlomis Species. Phytochem. Anal. 2005, 16, 1–6. [Google Scholar] [CrossRef]
- Okur, M.E.; Karadağ, A.E.; Üstündağ Okur, N.; Özhan, Y.; Sipahi, H.; Ayla, Ş.; Daylan, B.; Demirci, B.; Demirci, F. In Vivo Wound Healing and In Vitro Anti-Inflammatory Activity Evaluation of Phlomis Russeliana Extract Gel Formulations. Molecules 2020, 25, 2695. [Google Scholar] [CrossRef]
- Alpay, M.; Dulger, G.; Sahin, I.E.; Dulger, B. Evaluating Antimicrobial and Antioxidant Capacity of Endemic Phlomis Russeliana from Turkey and Its Antiproliferative Effect on Human Caco-2 Cell Lines. An. Acad. Bras. Cienc. 2019, 91, e20180404. [Google Scholar] [CrossRef] [PubMed]
- Akaydin, G.; Şimşek, I.; Arituluk, Z.C.; Yeşilada, E. An Ethnobotanical Survey in Selected Towns of the Mediterranean Subregion (Turkey). Turkish J. Biol. 2013, 37, 230–247. [Google Scholar] [CrossRef]
- Europe Council. European Pharmacopoeia 9.0; Europe Council: London, UK, 2017; ISBN 978-3-7692-6633-7. [Google Scholar]
- Shkondrov, A.; Krasteva, I.; Pavlova, D.; Zdraveva, P. Determination of Flavonoids in Related Astragalus Species (Sect. Incani) Occurring in Bulgaria. Comptes Rendus L’Academie Bulg. Sci. 2017, 70, 363–366. [Google Scholar]
- López, M.I.; Callao, M.P.; Ruisánchez, I. A Tutorial on the Validation of Qualitative Methods: From the Univariate to the Multivariate Approach. Anal. Chim. Acta 2015, 891, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Markham, K.R. Techniques of Flavonoid Identification; Academic Press: London, UK, 1982; Volume 31. [Google Scholar]
- Swartz, M.E.; Krull, I.S. Analytical Method Development and Validation; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Siddiqui, N.; Rauf, A.; Latif, A.; Mahmood, Z. Spectrophotometric Determination of the Total Phenolic Content, Spectral and Fluorescence Study of the Herbal Unani Drug Gul-e-Zoofa (Nepeta Bracteata Benth). J. Taibah Univ. Med. Sci. 2017, 12, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Fau, D.; Berson, A.; Eugene, D.; Fromenty, B.; Fisch, C.; Pessayre, D. Mechanism for the Hepatotoxicity of the Antiandrogen, Nilutamide. Evidence Suggesting That Redox Cycling of This Nitroaromatic Drug Leads to Oxidative Stress in Isolated Hepatocytes. J. Pharmacol. Exp. Ther. 1992, 263, 69–77. [Google Scholar]
- Mitcheva, M.; Kondeva, M.; Vitcheva, V.; Nedialkov, P.; Kitanov, G. Effect of Benzophenones from Hypericum Annulatum on Carbon Tetrachloride-Induced Toxicity in Freshly Isolated Rat Hepatocytes. Redox Rep. 2006, 11, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Abdelghffar, E.A.R.; El-Nashar, H.A.S.; Fayez, S.; Obaid, W.A.; Eldahshan, O.A. Ameliorative Effect of Oregano (Origanum Vulgare) versus Silymarin in Experimentally Induced Hepatic Encephalopathy. Sci. Rep. 2022, 12, 17854. [Google Scholar] [CrossRef]
- Kim, S.-H.; Choo, G.-S.; Yoo, E.-S.; Woo, J.-S.; Han, S.-H.; Lee, J.-H.; Jung, J.-Y. Silymarin Induces Inhibition of Growth and Apoptosis through Modulation of the MAPK Signaling Pathway in AGS Human Gastric Cancer Cells. Oncol. Rep. 2019, 42, 1904–1914. [Google Scholar] [CrossRef] [PubMed]
- Ahn, T.-H.; Yang, Y.-S.; Lee, J.-C.; Moon, C.-J.; Kim, S.-H.; Jun, W.; Park, S.-C.; Kim, J.-C. Ameliorative Effects of Pycnogenol® on Carbon Tetrachloride-Induced Hepatic Oxidative Damage in Rats. Phyther. Res. 2007, 21, 1015–1019. [Google Scholar] [CrossRef]
- Bai, X.; Qiu, A.; Guan, J.; Shi, Z. Antioxidant and Protective Effect of an Oleanolic Acid-Enriched Extract of A. Deliciosa Root on Carbon Tetrachloride Induced Rat Liver Injury. Asia Pac. J. Clin. Nutr. 2007, 16 (Suppl. 1), 169–173. [Google Scholar] [PubMed]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2018. [Google Scholar]
- International Council for Harmonisation of Technical Requirements for Pharmaceuticals for Human Use. Validation of Analytical Procedures: Text and Methodology Q2 (R2); International Council for Harmonisation of Technical Requirements for Pharmaceuticals for Human Use: Geneva, Switzerland, 2023. [Google Scholar]
- Wu, L.; Georgiev, M.I.; Cao, H.; Nahar, L.; El-Seedi, H.R.; Sarker, S.D.; Xiao, J.; Lu, B. Therapeutic Potential of Phenylethanoid Glycosides: A Systematic Review. Med. Res. Rev. 2020, 40, 2605–2649. [Google Scholar] [CrossRef] [PubMed]
- Timbrell, J. Factors Affecting Metabolism and Disposition. Toxic Responses to Foreign Compounds - Direct Toxic Action: Tissue Lessions. In Principles of Biochemical Toxicology, 3rd ed.; Timbrell, J., Ed.; Taylor and Francis: Oxfordshire, UK, 2000; pp. 175–180. [Google Scholar]
- Weber, L.W.D.; Boll, M.; Stampfl, A. Hepatotoxicity and Mechanism of Action of Haloalkanes: Carbon Tetrachloride as a Toxicological Model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef] [PubMed]
- Sridar, C.; Goosen, T.C.; Kent, U.M.; Williams, J.A.; Hollenberg, P.F. Silybin Inactivates Cytochromes P450 3A4 and 2C9 and Inhibits Major Hepatic Glucuronosyltransferases. Drug Metab. Dispos. 2004, 32, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Muriel, P.; Mourelle, M. Prevention by Silymarin of Membrane Alterations in Acute CCl4 Liver Damage. J. Appl. Toxicol. 1990, 10, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-N.; Hsieh, Y.-S.; Chiou, H.-L.; Chu, S.-C. Silibinin Inhibits Cell Invasion through Inactivation of Both PI3K-Akt and MAPK Signaling Pathways. Chem. Biol. Interact. 2005, 156, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Faroon, O. Toxicological Profile for Carbon Tetrachloride; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2005. [Google Scholar]
- Gu, H.; Gu, X.; Xu, Q.; Kang, W. Antioxidant Activity in Vitro and Hepatoprotective Effect of Phlomis Maximowiczii In Vivo. Afr. J. Tradit. Complement. Altern. Med. AJTCAM 2014, 11, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Usmanov, D.; Yusupova, U.; Syrov, V.; Ramazonov, N.; Rasulev, B. Iridoid Glucosides and Triterpene Acids from Phlomis Linearifolia, Growing in Uzbekistan and Its Hepatoprotective Activity. Nat. Prod. Res. 2021, 35, 2449–2453. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.L.; Van Wettere, A.J.; Cullen, J.M. Hepatobiliary System and Exocrine Pancreas1. In Pathologic Basis of Veterinary Disease; Zachary, J., Ed.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2017; Chapter 8; pp. 412–470. [Google Scholar]
- Kwon, A.J.; Morales, L.; Chatagnier, L.; Quigley, J.; Pascua, J.; Pinkowski, N.; Brasser, S.M.; Hong, M.Y. Effects of Moderate Ethanol Exposure on Risk Factors for Cardiovascular Disease and Colorectal Cancer in Adult Wistar Rats. Alcohol 2024. [Google Scholar] [CrossRef]
- Chen, X.; Shi, M.; Yang, L.; Guo, F.; Liang, Y.; Ma, L.; Fu, P.P. Phenylethanoid Glycoside Verbascoside Ameliorates Podocyte Injury of Diabetic Kidney Disease by Regulating NR4A1-LKB1-AMPK Signaling. Acta Mater. Medica 2023, 2, 72–83. [Google Scholar] [CrossRef]
- Slama, K.; Rouag, M.; Tichati, L.; Taibi, F.; Boumendjel, M.; Boumendjel, A.; Messarah, M. Nephroprotective Role and Antioxidant Capacity of Atriplex Halimus on Carbon Tetrachloride-Induced Kidney Damage in Rats. Comp. Clin. Path. 2021, 30, 75–87. [Google Scholar] [CrossRef]
- Gomaa, A.A.-R.; Samy, M.N.; Attia, E.Z.; Attya, M.E.; Fawzy, M.A.; Desoukey, S.Y.; Kamel, M.S. Antioxidant, Hepatoprotective and Nephroprotective Activities of Gazania Rigens against Carbon Tetrachloride-Induced Hepatotoxicity and Nephrotoxicity in Rats. Tradit. Med. Res. 2022, 7, 44. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | tR, min ± SD | EPhR |
|---|---|---|
| Flavonoids | ||
| Rhamnetin | 43.260 ± 0.18 | - |
| Apigenin | 37.830 ± 0.15 | - |
| Luteolin | 33.750 ± 0.12 | - |
| Naringin | 25.823 ± 0.10 | + |
| Saponarin | 21.227 ± 0.11 | - |
| Apigenin-7-glucuronide | 28.683 ± 0.13 | - |
| Apigenin-7-glucoside | 27.683 ± 0.12 | - |
| Luteolin-7-glucuronide | 25.740 ± 0.11 | + |
| Luteolin-3′-glucuronide | 28.810 ± 0.10 | + |
| Spiraeoside | 28.013 ± 0.13 | - |
| Kaempferol-3-glucoside | 27.210 ± 0.14 | - |
| Quercetin-3-glucoside | 25.157 ± 0.13 | + |
| Vitexin | 23.080 ± 0.12 | - |
| Phenylethanoids | ||
| Verbascoside | 24.470 ± 0.12 | + |
| Group | Hepatocytes’ Viability, % | LDH, µmol/min/106 Cells | GSH, nmol/106 Cells | MDA, nmol/106 Cells |
|---|---|---|---|---|
| Control | 86 ± 3.2 | 0.111 ± 0.01 | 20 ± 4.1 | 0.110 ± 0.01 |
| 5 µg/mL EPhR | 86 ± 3.1 | 0.115 ± 0.01 | 18 ± 4.4 | 0.115 ± 0.01 |
| 10 µg/mL EPhR | 85 ± 3.5 | 0.117 ± 0.01 | 17 ± 4.5 | 0.116 ± 0.01 |
| 50 µg/mL EPhR | 83 ± 3.8 | 0.118 ± 0.01 | 17 ± 4.8 | 0.118 ± 0.01 |
| 5 µg/mL S | 85 ± 3.7 | 0.114 ± 0.01 | 19 ± 4.1 | 0.113 ± 0.01 |
| 10 µg/mL S | 84 ± 3.5 | 0.115 ± 0.01 | 18 ± 3.9 | 0.114 ± 0.01 |
| 50 µg/mL S | 83 ± 3.7 | 0.115 ± 0.01 | 18 ± 3.8 | 0.115 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondeva-Burdina, M.; Shkondrov, A.; Popov, G.; Manov, V.; Krasteva, I. Antioxidant and Hepatoprotective Activity of an Extract from the Overground Parts of Phlomis russeliana Lag. ex Benth. Separations 2024, 11, 117. https://doi.org/10.3390/separations11040117
Kondeva-Burdina M, Shkondrov A, Popov G, Manov V, Krasteva I. Antioxidant and Hepatoprotective Activity of an Extract from the Overground Parts of Phlomis russeliana Lag. ex Benth. Separations. 2024; 11(4):117. https://doi.org/10.3390/separations11040117
Chicago/Turabian StyleKondeva-Burdina, Magdalena, Aleksandar Shkondrov, Georgi Popov, Vasil Manov, and Ilina Krasteva. 2024. "Antioxidant and Hepatoprotective Activity of an Extract from the Overground Parts of Phlomis russeliana Lag. ex Benth" Separations 11, no. 4: 117. https://doi.org/10.3390/separations11040117