
Papaya Seed Extract and Recovery of Some Main Constituents
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Sample Preparation
2.3. Microwave Drying
2.4. Oven Drying
2.5. Calibration Standard Solutions
2.6. UHPLC-PDA-ESI-TOF/MS Instrumentation and Conditions
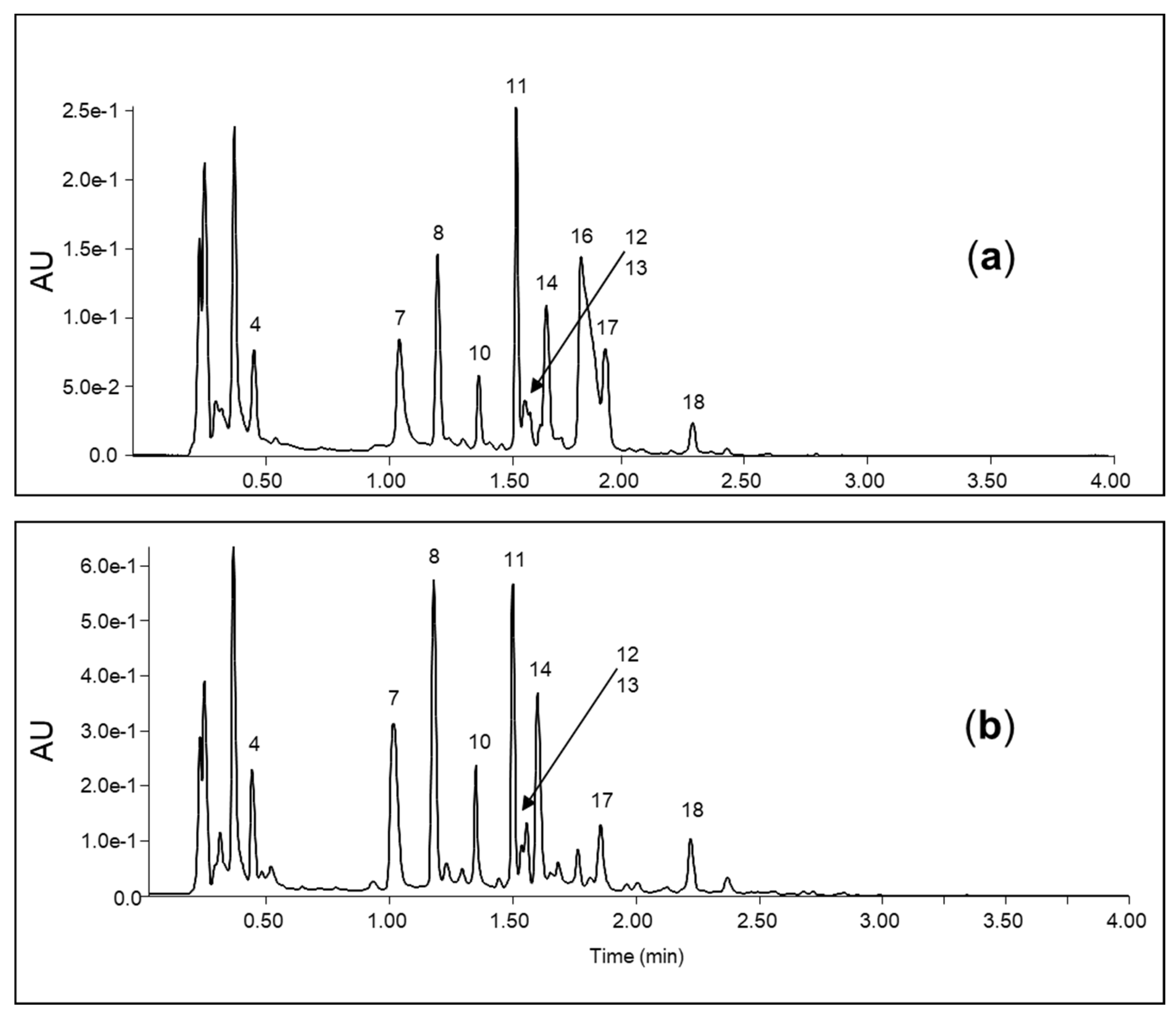
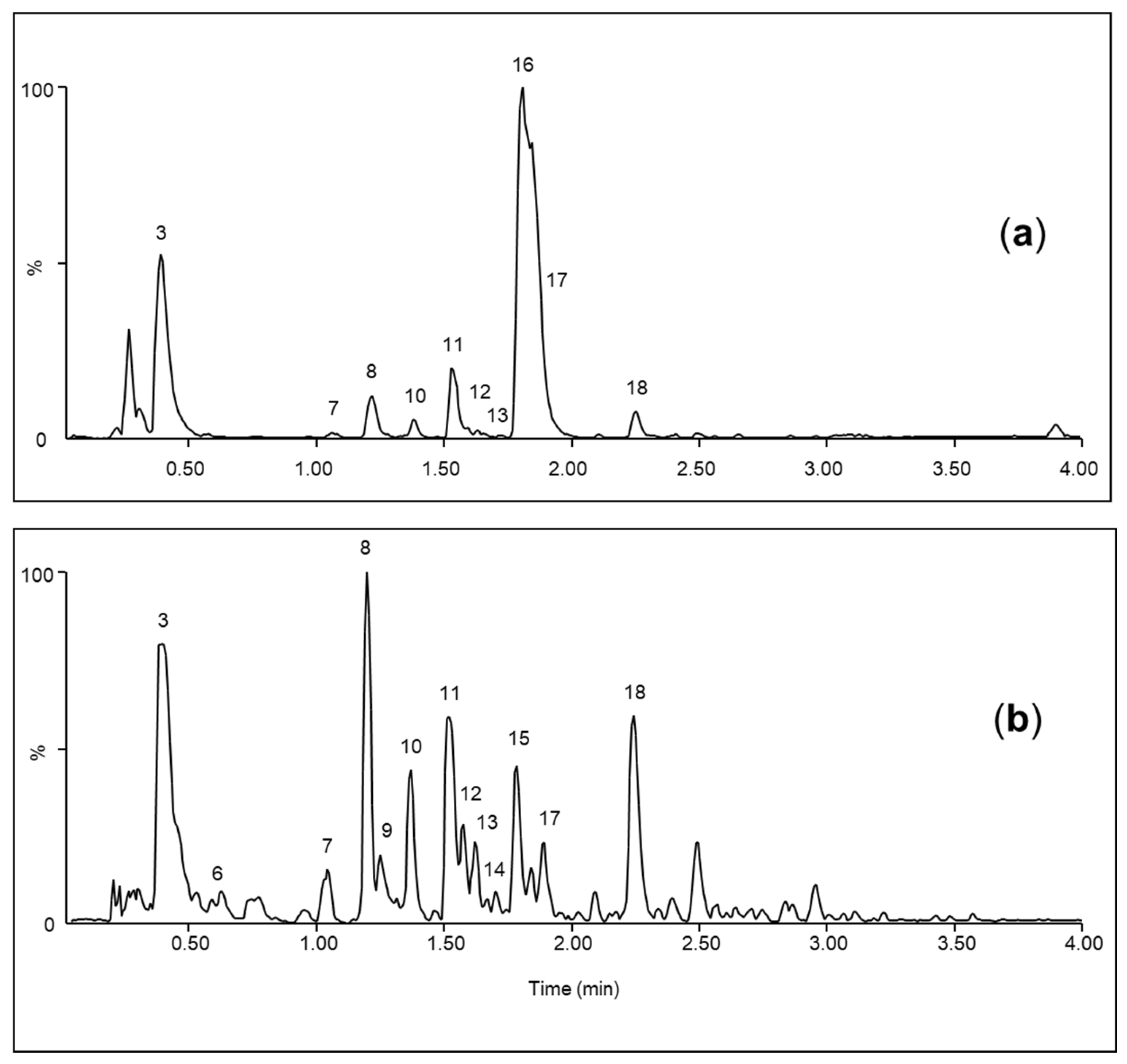
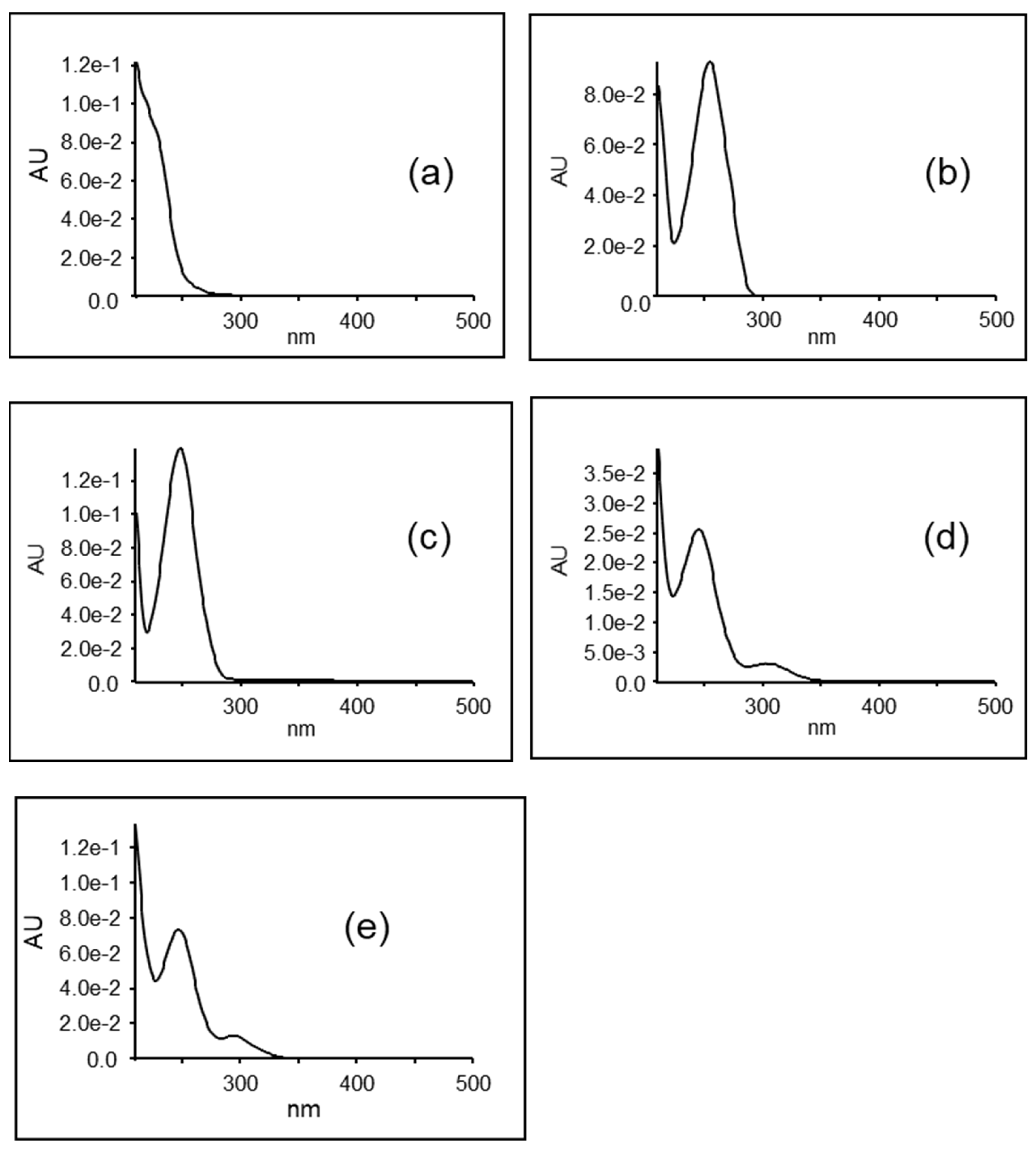
3. Results
3.1. Amino Acids
3.2. Organic Acids
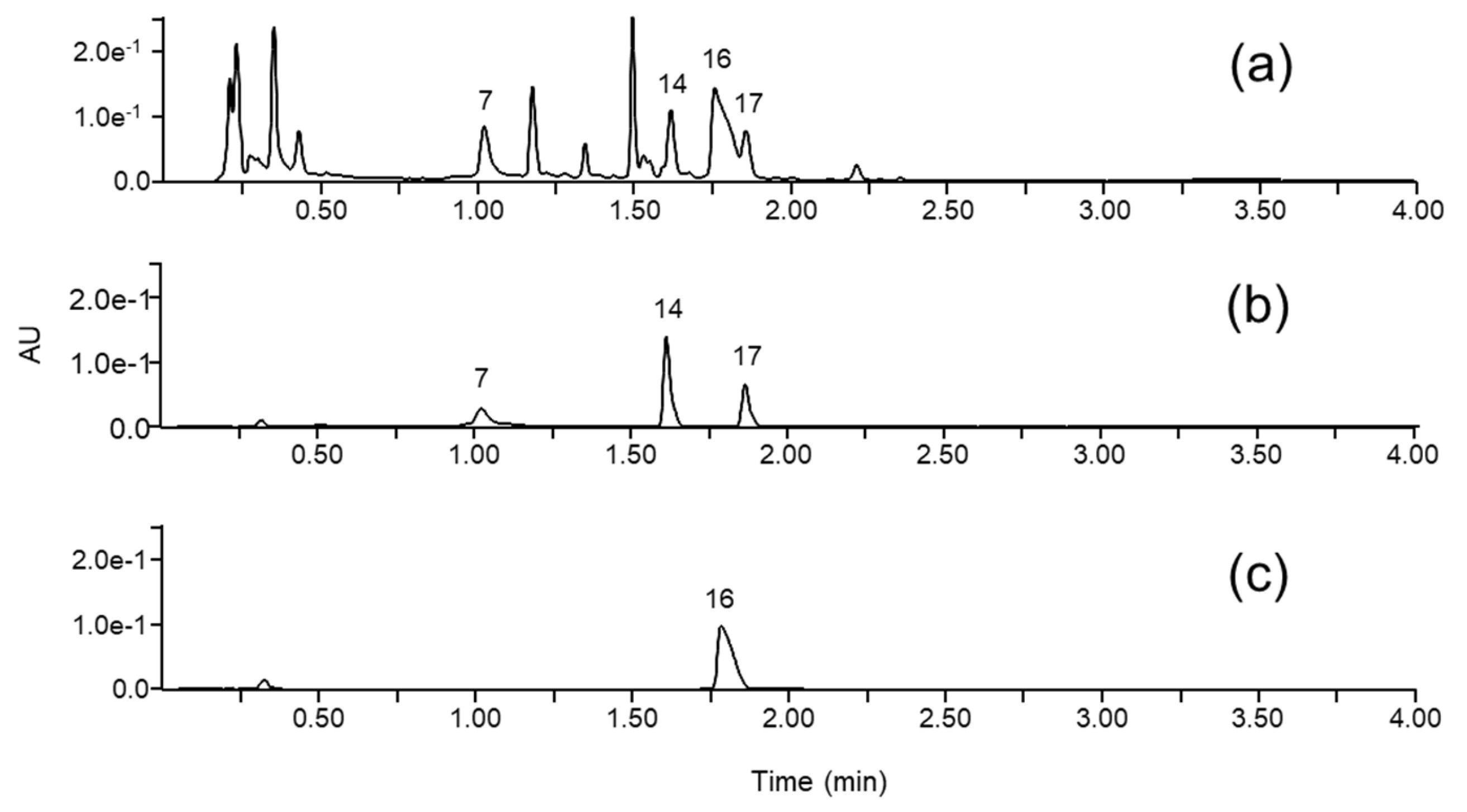
3.3. Glucotropaeolin and Other Glycosides
3.4. Flavonoids
3.5. Unknown Metabolites
3.6. UHPLC-PDA Quantification of Trp, GTL and 4-HBA
3.7. Recovery
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dos Santos, C.; De Abreu, C.; Freire, J.; Queiroz, E.; Mendonca, M. Chemical characterization of the flour of peel and seed from two papaya cultivars. Food Sci. Technol. 2014, 34, 353–357. [Google Scholar] [CrossRef]
- Williams, D.; Pun, S.; Chaliha, M.; Scheelings, P.; O’Hare, T. An unusual combination in papaya (Carica papaya): The good (glucosinolates) and the bad (cyanogenic glycosides). J. Food Comp. Anal. 2013, 29, 82–86. [Google Scholar] [CrossRef]
- Nugroho, A.; Heryani, H.; Choi, J.; Park, H. Identification and quantification of flavonoids in Carica papaya leaf and peroxynitritescavenging activity. Asian Pac. J. Trop. Biomed. 2017, 7, 208–213. [Google Scholar] [CrossRef]
- Rivera-Pastrana, D.; Yahia, E.; Gonzalez-Aguilar, G. Phenolic and carotenoid profiles of papaya fruit (Carica papaya L.) and their contents under low temperature storage. J. Sci. Food Agric. 2010, 90, 2358–2365. [Google Scholar] [CrossRef]
- Ikram, E.; Stanley, R.; Netzel, M.; Fanning, K. Phytochemicals of papaya and its traditional health and culinary uses—A review. J. Food Compos. Anal. 2015, 41, 201–211. [Google Scholar] [CrossRef]
- Barroso, P.T.; de Carvalho, P.P.; Rocha, T.B.; Pessoa, F.L.; Azevedo, D.A.; Mendes, M.F. Evaluation of the composition of Carica papaya L. seed oil extracted with supercritical CO2. Biotechnol. Rep. 2016, 11, 110–116. [Google Scholar] [CrossRef]
- Gogna, N.; Hamid, N.; Dorai, K. Metabolomic profiling of the phytomedicinal constituents of Carica papaya L. leaves and seeds by H-1 NMR spectroscopy and multivariate statistical analysis. J. Pharm. Biomed. Anal. 2015, 115, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D. Glucosinolates, structures and analysis in food. Anal. Methods 2010, 2, 310–325. [Google Scholar] [CrossRef]
- Fahey, J.; Zalcmann, A.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Vaughn, S.; Berhow, M. Glucosinolate hydrolysis products from various plant sources: pH effects isolation, and purification. Ind. Crops Prod. 2005, 21, 193–202. [Google Scholar] [CrossRef]
- Bennett, R.; Kiddle, G.; Wallsgrove, R. Biosynthesis of benzylglucosinolate, cyanogenic glucosides and phenylpropanoids in Carica papaya. Phytochemistry 1997, 45, 59–66. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yoshimoto, M.; Murata, Y.; Shimoishi, Y.; Asai, Y.; Park, E.; Sato, K. Papaya seed represents a rich source of biologically active isothiocyanate. J. Agric. Food Chem. 2007, 55, 4407–4413. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, M.; do Nascimento, J.; Purgatto, E.; Fabi, J.; Lajolo, F.; Cordenunsi, B. Benzylglucosinolate, Benzylisothiocyanate, and Myrosinase Activity in Papaya Fruit during Development and Ripening. J. Agric. Food Chem. 2008, 56, 9592–9599. [Google Scholar] [CrossRef]
- Gründemann, C.; Huber, R. Chemoprevention with isothiocyanates—From bench to bedside. Cancer Lett. 2018, 414, 26–33. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef] [PubMed]
- Conaway, C.; Yang, Y.; Chung, F. Isothiocyanates as cancer chemopreventive agents: Their biological activities and metabolism in rodents and humans. Curr. Drug Metab. 2002, 3, 233–255. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.; Kostov, R. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef]
- Kalkunte, S.; Swamy, N.; Dizon, D.S.; Brard, L. Benzyl isothiocyanate (BITC) induces apoptosis in ovarian cancer cells in vitro. J. Exp. Ther. Oncol. 2006, 5, 287–300. [Google Scholar] [PubMed]
- Li, Z.; Wang, Y.; Shen, W.; Zhou, P. Content determination of benzyl glucosinolate and anti-cancer activity of its hydrolysis product in Carica papaya L. Asian Pac. J. Trop. Med. 2012, 5, 231–233. [Google Scholar] [CrossRef]
- Li, P.; Zhao, Y.M.; Wang, C.; Zhu, H.P. Antibacterial activity and main action pathway of benzyl isothiocyanate extracted from papaya seeds. J. Food Sci. 2021, 86, 169–176. [Google Scholar] [CrossRef]
- Senrayan, J.; Venkatachalam, S. Solvent-assisted extraction of oil from papaya (Carica papaya L.) seeds: Evaluation of its physiochemical properties and fatty-acid composition. Sep. Sci. Technol. 2018, 53, 2852–2859. [Google Scholar] [CrossRef]
- Zhang, Y.; Kensler, T.; Cho, C.; Posner, G.; Talalay, P. Anticarcinogenic activities of sulforaphane and structurally related synthetic norbornyl isothiocyanates. Proc. Natl. Acad. Sci. USA 1994, 91, 3147–3150. [Google Scholar] [CrossRef]
- He, X.; Ma, Y.; Yi, G.; Wu, J.; Zhou, L.; Guo, H. Chemical composition and antifungal activity of Carica papaya Linn. seed essential oil against Candida spp. Lett. Appl. Microbiol. 2017, 64, 350–354. [Google Scholar] [CrossRef]
- Kermanshai, R.; McCarry, B.; Rosenfeld, J.; Summers, P.; Weretilnyk, E.; Sorger, G. Benzyl isothiocyanate is the chief or sole anthelmintic in papaya seed extracts. Phytochemistry 2001, 57, 427–435. [Google Scholar] [CrossRef]
- Pathak, N.; Mishra, P.; Manivannan, B.; Lohiya, N. Sterility due to inhibition of sperm motility by oral administration of benzene chromatographic fraction of the chloroform extract of the seeds of Carica papaya in rats. Phytomedicine 2000, 7, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Singh, O.; Ali, M. Phytochemical and antifungal profiles of the seeds of Carica Papaya L. Indian J. Pharm. Sci. 2011, 73, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, L.; Ortiz, A.; Cooke, R.; Fernandez, R. The dependence of crude papain yields on different collection (tapping) procedures for papaya latex. J. Sci. Food Agric. 1980, 31, 279–285. [Google Scholar] [CrossRef]
- Castro-Vargas, H.; Baumann, W.; Parada-Alfonso, F. Valorization of agroindustrial wastes: Identification by LC-MS and NMR of benzylglucosinolate from papaya (Carica papaya L.) seeds, a protective agent against lipid oxidation in edible oils. Electrophoresis 2016, 37, 1930–1937. [Google Scholar] [CrossRef]
- Van Eylen, D.; Hendrickx, M.; Van Loey, A. Temperature and pressure stability of mustard seed (Sinapis alba L.) myrosinase. Food Chem. 2006, 97, 263–271. [Google Scholar] [CrossRef]
- Panusa, A.; Petrucci, R.; Marrosu, G.; Multari, G.; Gallo, F. UHPLC-PDA-ESI-TOF/MS metabolic profiling of Arctostaphylos pungens and Arctostaphylos uva-ursi A comparative study of phenolic compounds from leaf methanolic extracts. Phytochemistry 2015, 115, 79–88. [Google Scholar] [CrossRef]
- Alonso-Salces, R.; Ndjoko, K.; Queiroz, E.; Ioset, J.; Hostettmann, K.; Berrueta, L.; Gallo, B.; Vicente, F. On-line characterisation of apple polyphenols by liquid chromatography coupled with mass spectrometry and ultraviolet absorbance detection. J. Chromatogr. A 2004, 1046, 89–100. [Google Scholar] [CrossRef]
- Sumner, L.; Amberg, A.; Barrett, D.; Beale, M.; Beger, R.; Daykin, C.; Fan, T.; Fiehn, O.; Goodacre, R.; Griffin, J.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- ISO 9167:2019; Rapeseed and Rapeseed Meals—Determination of Glucosinolates Content—Method Using High-Performance Liquid Chromatography. ISO: Geneva, Switzerland, 2019. Available online: https://www.iso.org/obp/ui/en/#iso:std:iso:9167:ed-1:v1:en (accessed on 12 August 2024).
- Matthaus, B.; Luftmann, H. Glucosinolates in members of the family Brassicaceae: Separation and identification by LC/ESI-MS-MS. J. Agric. Food Chem. 2000, 48, 2234–2239. [Google Scholar] [CrossRef]
- Mellon, F.; Bennett, R.; Holst, B.; Williamson, G. Intact glucosinolate analysis in plant extracts by programmed cone voltage electrospray LC/MS: Performance and comparison with LC/MS/MS methods. Anal. Biochem. 2002, 306, 83–91. [Google Scholar] [CrossRef]
- Ares, A.; Valverde, S.; Nozal, M.; Bernal, J.; Bernal, J. Development and validation of a specific method to quantify intact glucosinolates in honey by LC-MS/MS. J. Food Compos. Anal. 2016, 46, 114–122. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, M.J.; Jeong, M.H.; Kim, J.E. Identification and Quantification of Glucosinolates in Kimchi by Liquid Chromatography-Electrospray Tandem Mass Spectrometry. Int. J. Anal. Chem. 2017, 2017, 6753481. [Google Scholar] [CrossRef]
- Vallejo-Castillo, V.; Muñoz-Mera, J.; Pérez-Bustos, M.; Rodriguez-Stouvenel, A. Recovery of antioxidants from Papaya (Carica papaya L.) peel and pulp by microwave-assisted extraction. Rev. Mex. Ing. Quim 2020, 19, 85–98. [Google Scholar] [CrossRef]
- Sticha, K.; Kenney, P.; Boysen, G.; Liang, H.; Su, X.; Wang, M.; Upadhyaya, P.; Hecht, S. Effects of benzyl isothiocyanate and phenethyl isothiocyanate on DNA adduct formation by a mixture of benzo[a]pyrene and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone in A/J mouse lung. Carcinogenesis 2002, 23, 1433–1439. [Google Scholar] [CrossRef]
- Miyoshi, N.; Takabayashi, S.; Osawa, T.; Nakamura, Y. Benzyl isothiocyanate inhibits excessive superoxide generation in inflammatory leukocytes: Implication for prevention against inflammation-related carcinogenesis. Carcinogenesis 2004, 25, 567–575. [Google Scholar] [CrossRef]
- Dinh, T.; Parat, M.; Ong, Y.; Khaw, K. Anticancer activities of dietary benzyl isothiocyanate: A comprehensive review. Pharm. Res. 2021, 169, 105666. [Google Scholar] [CrossRef]
- Wong, R. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87–101. [Google Scholar] [CrossRef]
- Lui, V.; Wentzel, A.; Xiao, D.; Lew, K.; Singh, S.; Grandis, J. Requirement of a carbon spacer in benzyl isothiocyanate-mediated cytotoxicity and MAPK activation in head and neck squamous cell carcinoma. Carcinogenesis 2003, 24, 1705–1712. [Google Scholar] [CrossRef]
- Lai, K.; Huang, A.; Hsu, S.; Kuo, C.; Yang, J.; Wu, S.; Chung, J. Benzyl Isothiocyanate (BITC) Inhibits Migration and Invasion of Human Colon Cancer HT29 Cells by Inhibiting Matrix Metalloproteinase-2/-9 and Urokinase Plasminogen (uPA) through PKC and MAPK Signaling Pathway. J. Agric. Food Chem. 2010, 58, 2935–2942. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, L.; Meng, Y.; Huang, L. Benzyl isothiocyanate inhibits breast cancer cell tumorigenesis via repression of the FoxH1-Mediated Wnt/β-catenin pathway. Int. J. Clin. Exp. Med. 2015, 8, 17601–17611. [Google Scholar] [PubMed]
- Mi, L.; Gan, N.; Chung, F. Isothiocyanates inhibit proteasome activity and proliferation of multiple myeloma cells. Cancer Res. 2011, 71, 5130–5142. [Google Scholar] [CrossRef]
- Drobnica, L.; Kristian, P.J.; Augustin, J. Cyanates and their Thio Derivatives Part 2. In The Chemistry of Cyanates and Their Thio Derivatives; Patai, S., Ed.; John Wiley & Sons Ltd.: Chichester, UK, 1977; pp. 1003–1221. [Google Scholar] [CrossRef]
- Getahun, S.M.; Chung, F.L. Conversion of glucosinolates to isothiocyanates in humans after ingestion of cooked watercress. Cancer Epidemiol. Biomark. Prev. 1999, 8, 447–451. [Google Scholar] [PubMed]
- Adeoye, R.I.; Olopade, E.T.; Olayemi, I.O.; Okaiyeto, K.; Akiibinu, M.O. Nutritional and therapeutic potentials of Carica papaya Linn. seed: A comprehensive review. Plant Sci. Today 2024, 11, 671–680. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
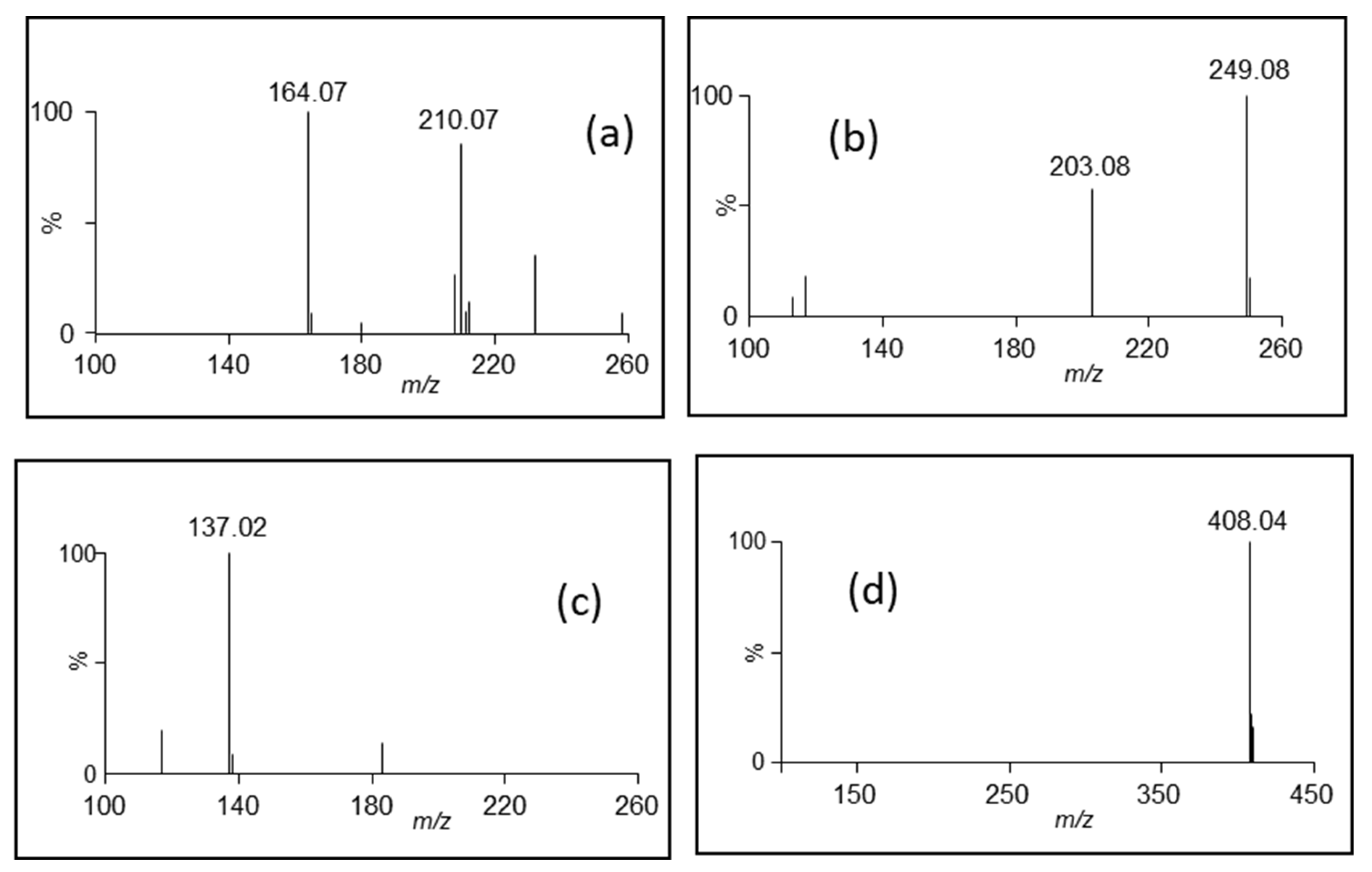
| No. | RT (min) | λ max (nm) | [M-H]− m/z | [M+H]+ m/z | Chemical Formula | Compound |
| 1 | 0.23 | 116.07 | C5H9NO2 | Proline * | ||
| 2 | 0.27 | 118.09 | C5H11NO2 | Valine * | ||
| 3 | 0.35 | 191.02 | C6H8O7 | Citric acid * | ||
| 4 | 0.43 | 223/273 | 182.08 | C9H11NO3 | Tyrosine * | |
| 5 | 0.47 | 130.09 | 132.10 | C6H13NO2 | Leucine * | |
| 6 | 0.59 | 292.14 | 294.15 | Unknown | ||
| 7 | 1.01 | 257 | 164.07 | 166.09 | C9H11NO2 | Phenylalanine * |
| 8 | 1.17 | 248 | 299.08 | 301.09 | C13H16O8 | 4-hydroxybenzoic acid glycoside ** |
| 9 | 1.25 | 326 | Unknown | |||
| 10 | 1.33 | 248 | 461.13 | [M+Na]+ 485.13 | C19H26O13 | 4-hydroxybenzoic acid diglycoside ** |
| 11 | 1.49 | 248 | 623.18 | [M+Na]+ 647.18 | C25H36O18 | 4-hydroxybenzoic acid triglycoside ** |
| 12 | 1.54 | 247/295 | 801.21 | C34H42O22 | Rhamnetin triglycoside 1 ** | |
| 13 | 1.61 | 244/310 | 801.21 | C34H42O22 | Rhamnetin triglycoside 2 ** | |
| 14 | 1.64 | 218/279 | 203.08 | 205.10 | C11H12N2O2 | Tryptophan * |
| 15 | 1.74 | 313.09 | Unknown | |||
| 16 | 1.76 | 240 sh | 408.04 | [M+H-SO3]+ 330.10 | C14H19NO9S2 | Glucotropaeolin * |
| 17 | 1.85 | 254 | 137.02 | C7H6O3 | 4-hydroxybenzoic acid * | |
| 18 | 2.20 | 230 | 236.06 | 238.07 | Unknown |
| Myrosinase Deactivation | Trp | GTL | 4-HBA |
|---|---|---|---|
| Microwave | 69 ± 1 | 245 ± 72 | 56 ± 13 |
| Oven | 85 ± 5 | 308 ± 31 | 74 ± 19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panusa, A.; Mammone, F.R.; Rotundo, P.; Multari, G.; Palazzino, G.; Gallo, F.R. Papaya Seed Extract and Recovery of Some Main Constituents. Separations 2024, 11, 254. https://doi.org/10.3390/separations11090254
Panusa A, Mammone FR, Rotundo P, Multari G, Palazzino G, Gallo FR. Papaya Seed Extract and Recovery of Some Main Constituents. Separations. 2024; 11(9):254. https://doi.org/10.3390/separations11090254
Chicago/Turabian StylePanusa, Alessia, Francesca Romana Mammone, Paola Rotundo, Giuseppina Multari, Giovanna Palazzino, and Francesca Romana Gallo. 2024. "Papaya Seed Extract and Recovery of Some Main Constituents" Separations 11, no. 9: 254. https://doi.org/10.3390/separations11090254