Combined TPEF and SHG Imaging for the Microstructural Characterization of Different Wood Species Used in Artworks

 ,
,  ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
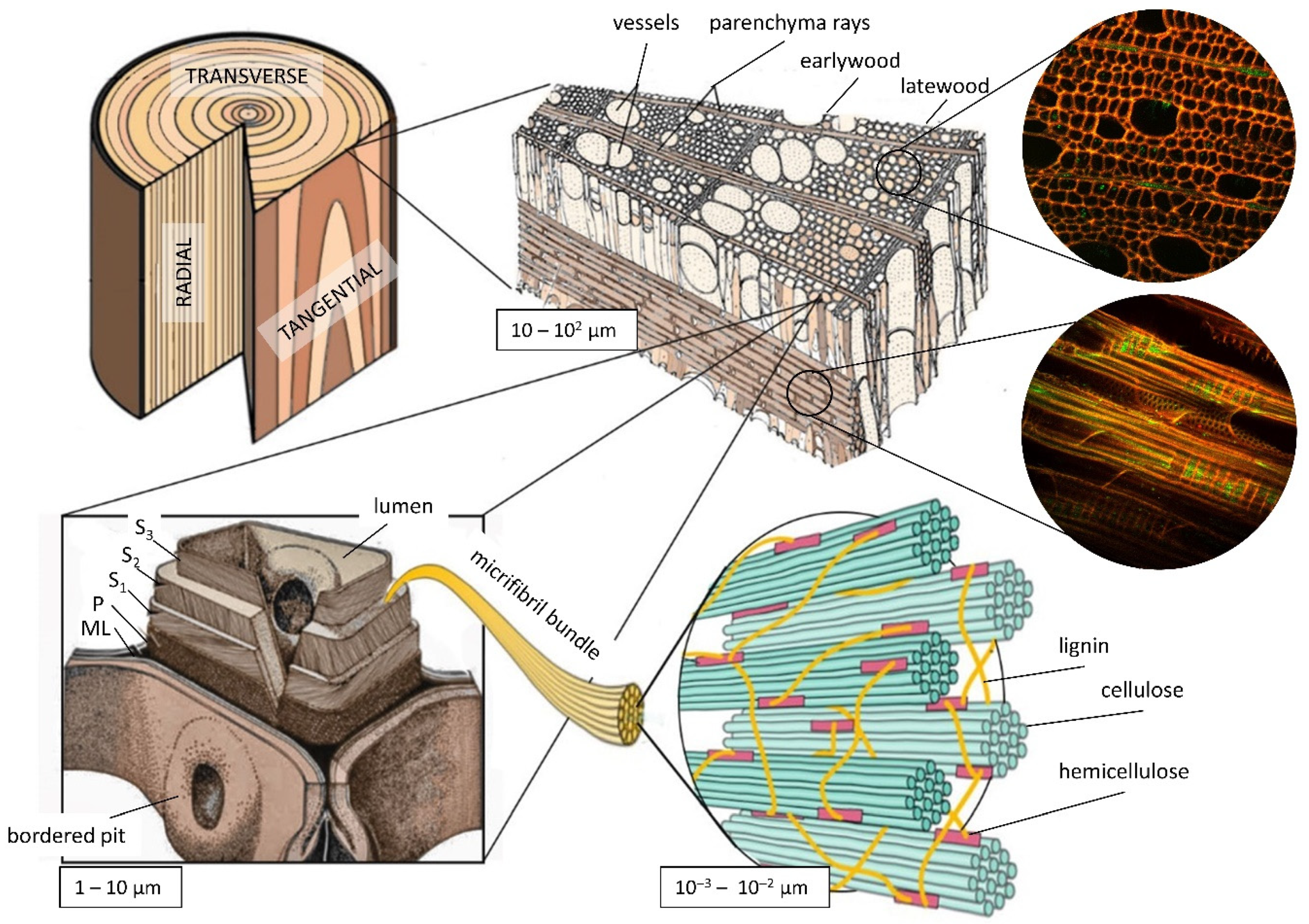
2.1. Samples
2.2. Microstructural Features
2.3. Nonlinear Setup
3. Results
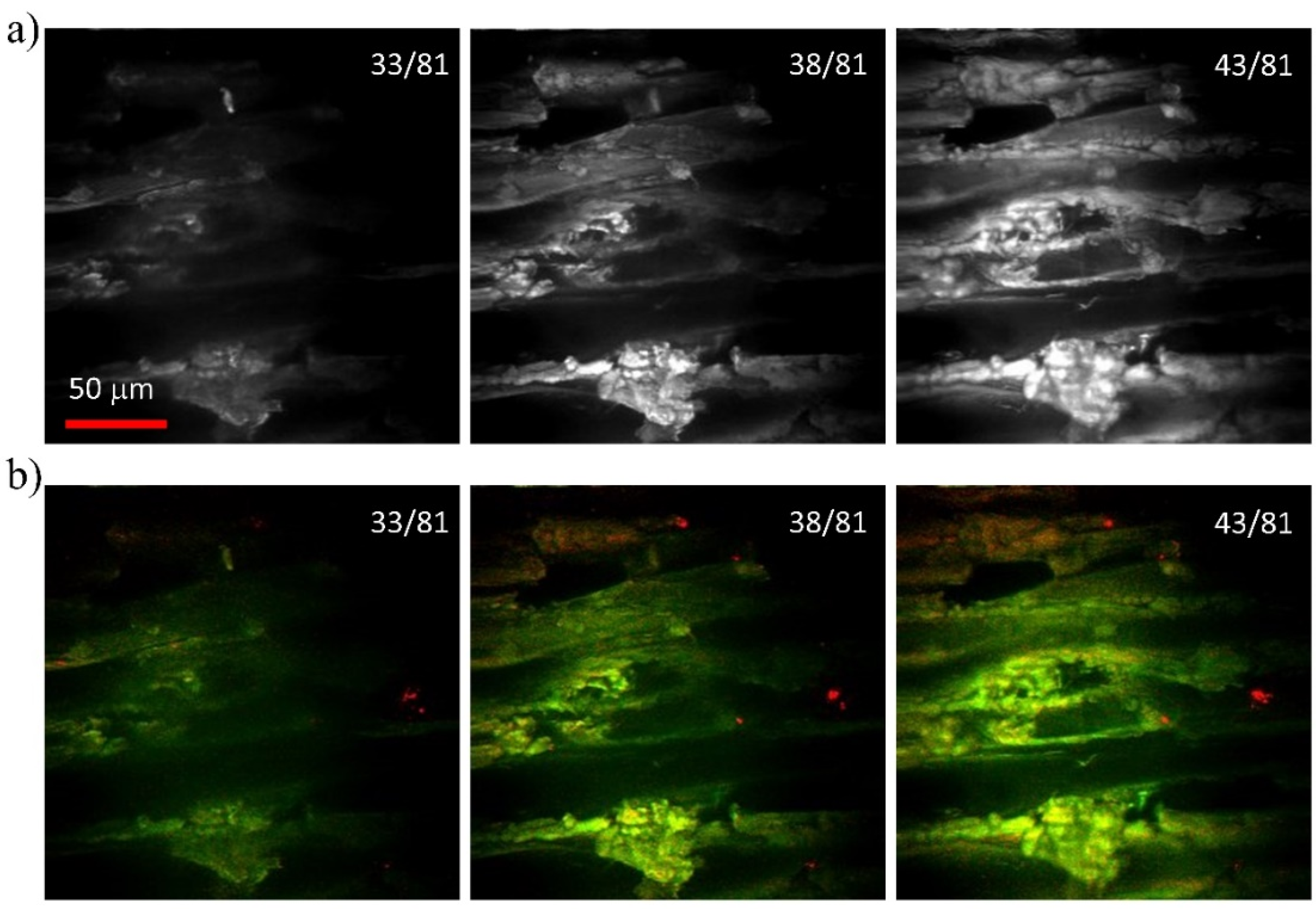
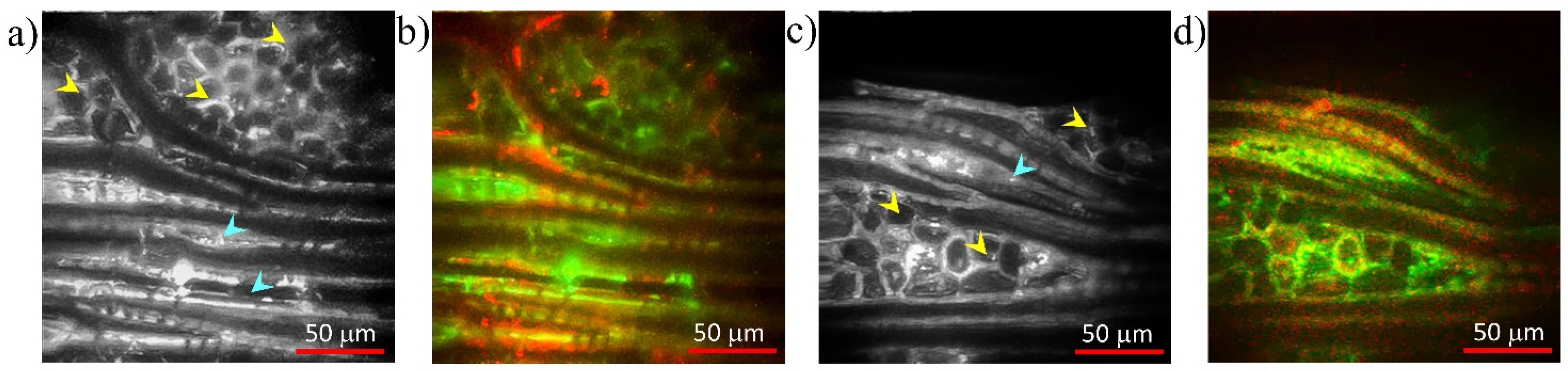
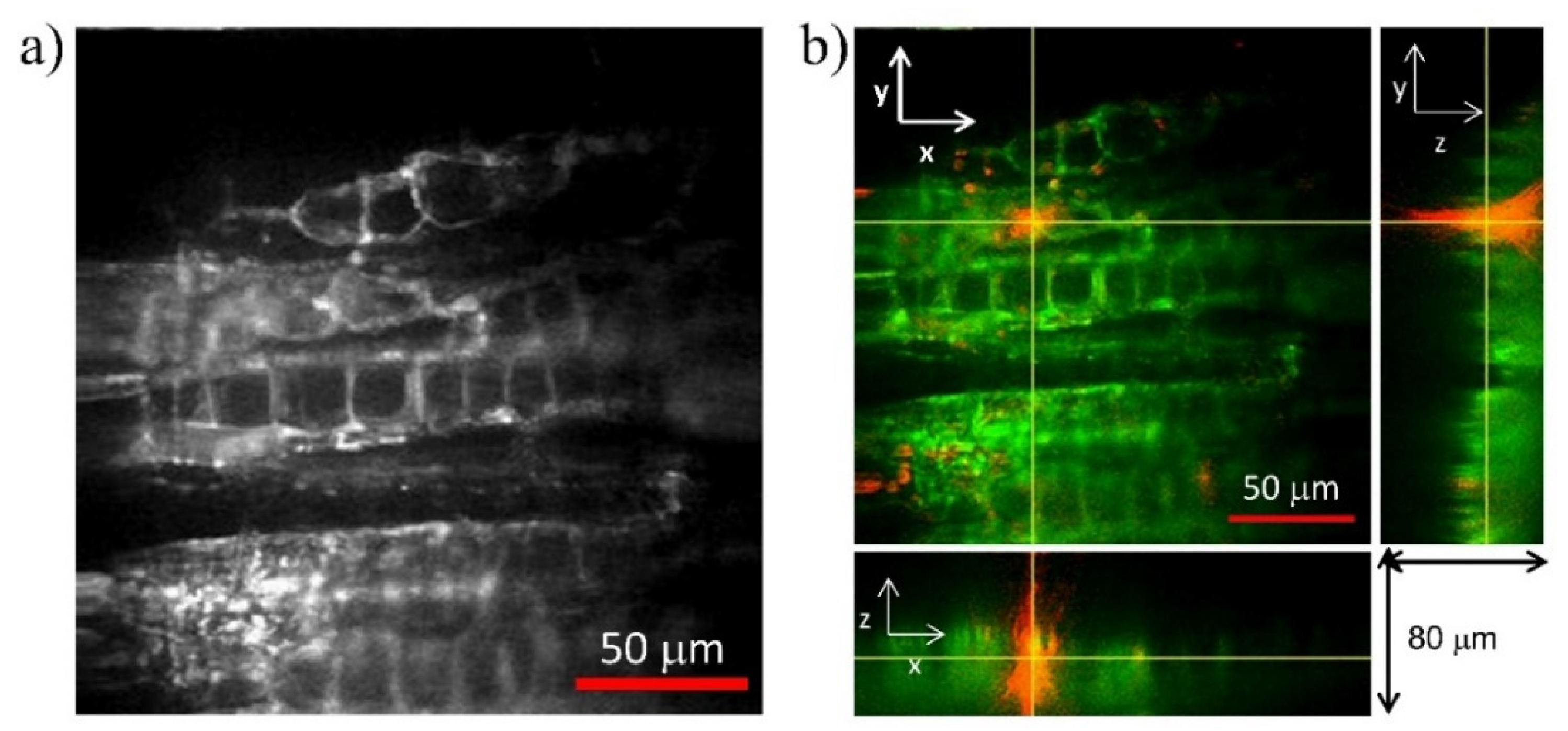
3.1. Poplar (Populus tremula L.)
3.2. Beech (Fagus sylvatica L.)
3.3. Chestnut (Castanea sativa M.)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Stagno, V.; Egizi, F.; Corticelli, F.; Morandi, V.; Valle, F.; Costantini, G.; Longo, S.; Capuani, S. Microstructural features assessment of different waterlogged wood species by NMR diffusion validated with complementary techniques. Magn. Reson. Imaging 2021, 83, 139–151. [Google Scholar] [CrossRef]
- Plötze, M.; Niemz, P. Porosity and pore size distribution of different wood types as determined by mercury intrusion porosimetry. Eur. J. Wood Wood Prod. 2011, 69, 649–657. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Man, Y.; Wen, J.; Guo, Y.; Lin, J. Advances in Imaging Plant Cell Walls. Trends Plant Sci. 2019, 24, 867–878. [Google Scholar] [CrossRef]
- Jeffries, T.W. Physical, chemical and biochemical considerations in the biological degradation of wood. US Forest Products Laboratory. In Wood and Cellulosics: Industrial Utilisation, Biotechnology, Structure and Properties; Kennedy, J.F., Phillips, G.O., William, P.A., Eds.; Ellis Horwood Ltd.: Chichester, UK, 1988; Chapter 24; pp. 213–230. [Google Scholar]
- Oltean, L.; Teischinger, A.; Hansmann, C. Influence of temperature on cracking and mechanical properties of wood during wood drying–A review. BioResources 2007, 2, 789–811. [Google Scholar]
- Mizutani, G.; Sonoda, Y.; Sano, H.; Sakamoto, M.; Takahashi, T.; Ushioda, S. Detection of starch granules in a living plant by optical second harmonic microscopy. J. Lumin. 2000, 87–89, 824–826. [Google Scholar] [CrossRef]
- Chu, S.-W.; Chen, I.-H.; Liu, T.-M.; Chen, P.C.; Sun, C.-K.; Lin, B.-L. Multimodal nonlinear spectral microscopy based on a femtosecond Cr:forsterite laser. Opt. Lett. 2001, 26, 1909–1911. [Google Scholar] [CrossRef]
- Cox, G.; Moreno, N.; Feijó, J. Second-harmonic imaging of plant polysaccharides. J. Biomed. Opt. 2005, 10, 024013. [Google Scholar] [CrossRef]
- Parodi, V.; Jacchetti, E.; Osellame, R.; Cerullo, G.; Polli, D.; Raimondi, M.T. Nonlinear Optical Microscopy: From Fundamentals to Applications in Live Bioimaging. Front. Bioeng. Biotechnol. 2020, 8, 585363. [Google Scholar] [CrossRef]
- Gratton, E.; Barry, N.P.; Beretta, S.; Celli, A. Multiphoton fluorescence microscopy. Methods 2001, 25, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Denk, W.; Strickler, J.H.; Webb, W.W. Two-photon laser scanning fluorescence microscope. Science 1990, 248, 73–76. [Google Scholar] [CrossRef] [Green Version]
- Hell, S.W.; Bahlmann, K.; Schrader, M.; Soini, A.; Malak, H.M.; Gryczynski, I.; Lakowicz, J.R. Three-photon excitation in fluorescence microscopy. J. Biomed. Opt. 1996, 1, 71–74. [Google Scholar] [CrossRef]
- Gauderon, R.; Lukins, P.; Sheppard, C. Optimization of second-harmonic generation microscopy. Micron 2001, 32, 691–700. [Google Scholar] [CrossRef]
- Oron, D.; Yelin, D.; Tal, E.; Raz, S.; Fachima, R.; Silberberg, Y. Depth-resolved structural imaging by third-harmonic generation microscopy. J. Struct. Biol. 2004, 147, 3–11. [Google Scholar] [CrossRef]
- Fovo, A.D.; Castillejo, M.; Fontana, R. Nonlinear optical microscopy for artworks physics. Riv. Nuovo Cim. 2021, 44, 453–498. [Google Scholar] [CrossRef]
- Mertz, J. Nonlinear microscopy: New techniques and applications. Curr. Opin. Neurobiol. 2004, 14, 610–616. [Google Scholar] [CrossRef]
- Fovo, A.D.; Sanz, M.; Mattana, S.; Oujja, M.; Marchetti, M.; Pavone, F.; Cicchi, R.; Fontana, R.; Castillejo, M. Safe limits for the application of nonlinear optical microscopies to cultural heritage: A new method for in-situ assessment. Microchem. J. 2020, 154, 104568. [Google Scholar] [CrossRef]
- Latour, G.; Echard, J.-P.; Soulier, B.; Emond, I.; Vaiedelich, S.; Elias, M. Structural and optical properties of wood and wood finishes studied using optical coherence tomography: Application to an 18th century Italian violin. Appl. Opt. 2009, 48, 6485–6491. [Google Scholar] [CrossRef]
- Popescu, M.-C.; Popescu, C.-M.; Lisa, G.; Sakata, Y. Evaluation of morphological and chemical aspects of different wood species by spectroscopy and thermal methods. J. Mol. Struct. 2011, 988, 65–72. [Google Scholar] [CrossRef]
- Brown, R.M., Jr.; Millard, A.C.; Campagnola, P.J. Macromolecular structure of cellulose studied by second-harmonic generation imaging microscopy. Opt. Lett. 2003, 28, 2207–2209. [Google Scholar] [CrossRef]
- Psilodimitrakopoulos, S.; Amat-Roldan, I.; Loza-Alvarez, P.; Artigas, D. Estimating the helical pitch angle of amylopectin in starch using polarization second harmonic generation microscopy. J. Opt. 2010, 12, 084007. [Google Scholar] [CrossRef]
- Psilodimitrakopoulos, S.; Gavgiotaki, E.; Melessanaki, K.; Tsafas, V.; Filippidis, G. Polarization Second Harmonic Generation Discriminates Between Fresh and Aged Starch-Based Adhesives Used in Cultural Heritage. Microsc. Microanal. 2016, 22, 1072–1083. [Google Scholar] [CrossRef]
- Xue, Y.; Qiu, X.; Ouyang, X. Insights into the effect of aggregation on lignin fluorescence and its application for microstructure analysis. Int. J. Biol. Macromol. 2020, 154, 981–988. [Google Scholar] [CrossRef]
- Djikanović, D.; Kalauzi, A.; Radotić, K.; Lapierre, C.; Jeremić, M. Deconvolution of lignin fluorescence spectra: A contribution to the comparative structural studies of lignins. Russ. J. Phys. Chem. A 2007, 81, 1425–1428. [Google Scholar] [CrossRef]
- Donaldson, L. Softwood and Hardwood Lignin Fluorescence Spectra of Wood Cell Walls in Different Mounting Media. IAWA J. 2013, 34, 3–19. [Google Scholar] [CrossRef]
- Donaldson, L. Autofluorescence in Plants. Molecules 2020, 25, 2393. [Google Scholar] [CrossRef]
- Latour, G.; Echard, J.-P.; Didier, M.; Schanne-Klein, M.-C. In situ 3D characterization of historical coatings and wood using multimodal nonlinear optical microscopy. Opt. Express 2012, 20, 24623–24635. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, L.; Radotic, K. Fluorescence lifetime imaging of lignin autofluorescence in normal and compression wood. J. Microsc. 2013, 251, 178–187. [Google Scholar] [CrossRef]
- Débarre, D.; Olivier, N.; Beaurepaire, E. Signal epidetection in third-harmonic generation microscopy of turbid media. Opt. Express 2007, 15, 8913. [Google Scholar] [CrossRef] [Green Version]
- Dammström, S.; Salmén, L.; Gatenholm, P. On the interactions between cellulose and xylan, a biomimetic simulation of the hardwood cell wall. BioResources 2009, 4, 3–14. [Google Scholar]
- Fujita, M.; Harada, H. Ultrastructure and formation of wood cell wall. In Wood and Cellulosic Chemistry; Hon, D.N.S., Shiraishi, N., Eds.; Marcel Dekker: New York, NY, USA, 2001. [Google Scholar]
- Choat, B.; Cobb, A.R.; Jansen, S. Structure and function of bordered pits: New discoveries and impacts on whole-plant hydraulic function. New Phytol. 2008, 177, 608–626. [Google Scholar] [CrossRef]
- Simmons, T.; Mortimer, J.; Bernardinelli, O.; Pöppler, A.-C.; Brown, S.; DeAzevedo, E.; Dupree, R.; Dupree, P. Folding of xylan onto cellulose fibrils in plant cell walls revealed by solid-state NMR. Nat. Commun. 2016, 7, 13902. [Google Scholar] [CrossRef]
- Rencurosi, A.; Röhrling, J.; Pauli, J.; Potthast, A.; Jäger, C.; Pérez, S.; Kosma, P.; Imberty, A. Polymorphism in the Crystal Structure of the Cellulose Fragment Analogue Methyl 4-O-Methyl-β-D-Glucopyranosyl-(1-4)-β-D-Glucopyranoside. Angew. Chem. Int. Ed. 2002, 41, 4277–4281. [Google Scholar] [CrossRef]
- Berti, S.; Fioravanti, M.; Macchioni, N. La Struttura Anatomica del Legno ed il Riconoscimento dei Legnami Italiani di Più Ricorrente Impiego; Berti, N., Ed.; CNE-INVALSA: Sesto Fiorentino, Italy, 2006. [Google Scholar]
- Marchetti, M.; Baria, E.; Cicchi, R.; Pavone, F.S. Custom Multiphoton/Raman Microscopy Setup for Imaging and Characterization of Biological Samples. Methods Protoc. 2019, 2, 51. [Google Scholar] [CrossRef] [Green Version]
- Fromm, J.; Rockel, B.; Lautner, S.; Windeisen, E.; Wanner, G. Lignin distribution in wood cell walls determined by TEM and backscattered SEM techniques. J. Struct. Biol. 2003, 143, 77–84. [Google Scholar] [CrossRef]
- Nadiarnykh, O.; LaComb, R.B.; Campagnola, P.J.; Mohler, W.A. Coherent and incoherent SHG in fibrillar cellulose matrices. Opt. Express 2007, 15, 3348–3360. [Google Scholar] [CrossRef]
- Chen, B.-C.; Sung, J.; Lim, S.-H. Chemical Imaging with Frequency Modulation Coherent Anti-Stokes Raman Scattering Microscopy at the Vibrational Fingerprint Region. J. Phys. Chem. B 2010, 114, 16871–16880. [Google Scholar] [CrossRef] [Green Version]
- Peciulyte, A.; Kiskis, J.; Larsson, P.T.; Olsson, L.; Enejder, A. Visualization of structural changes in cellulosic substrates during enzymatic hydrolysis using multimodal nonlinear microscopy. Cellulose 2016, 23, 1521–1536. [Google Scholar] [CrossRef] [Green Version]
- Tai, H.-C.; Chen, P.-L.; Xu, J.-W.; Chen, S.-Y. Two-photon fluorescence and second harmonic generation hyperspectral imaging of old and modern spruce woods. Opt. Express 2020, 28, 38831–38841. [Google Scholar] [CrossRef]
- Lionetto, F.; Del Sole, R.; Cannoletta, D.; Vasapollo, G.; Maffezzoli, A. Monitoring Wood Degradation during Weathering by Cellulose Crystallinity. Materials 2012, 5, 1910–1922. [Google Scholar] [CrossRef] [Green Version]
- Howell, C.; Hastrup, A.C.S.; Goodell, B.; Jellison, J. Temporal changes in wood crystalline cellulose during degradation by brown rot fungi. Int. Biodeterior. Biodegrad. 2009, 63, 414–419. [Google Scholar] [CrossRef]
- Budrugeac, P.; Emandi, A. The use of thermal analysis methods for conservation state determination of historical and/or cultural objects manufactured from lime tree wood. J. Therm. Anal. 2010, 101, 881–886. [Google Scholar] [CrossRef]
- Cicchi, R.; Kapsokalyvas, D.; Stampouli, D.; De Giorgi, V.; Massi, D.; Lotti, T.; Pavone, F.S. In-vivo tissue imaging using a compact mobile nonlinear microscope. In Proceedings of the Multiphoton Microscopy in the Biomedical Sciences X, San Francisco, CA, USA, 23–28 January 2011; Volume 7569, p. 756930. [Google Scholar] [CrossRef]
- König, K. High-resolution multimodal clinical multiphoton tomography of skin. In Proceedings of the Photonic Therapeutics and Diagnostics VII, San Francisco, CA, USA, 22–27 January 2011; Volume 7883, p. 78830D. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dal Fovo, A.; Mattana, S.; Marchetti, M.; Anichini, M.; Giovannelli, A.; Baria, E.; Fontana, R.; Cicchi, R. Combined TPEF and SHG Imaging for the Microstructural Characterization of Different Wood Species Used in Artworks. Photonics 2022, 9, 170. https://doi.org/10.3390/photonics9030170
Dal Fovo A, Mattana S, Marchetti M, Anichini M, Giovannelli A, Baria E, Fontana R, Cicchi R. Combined TPEF and SHG Imaging for the Microstructural Characterization of Different Wood Species Used in Artworks. Photonics. 2022; 9(3):170. https://doi.org/10.3390/photonics9030170
Chicago/Turabian StyleDal Fovo, Alice, Sara Mattana, Marco Marchetti, Monica Anichini, Alessio Giovannelli, Enrico Baria, Raffaella Fontana, and Riccardo Cicchi. 2022. "Combined TPEF and SHG Imaging for the Microstructural Characterization of Different Wood Species Used in Artworks" Photonics 9, no. 3: 170. https://doi.org/10.3390/photonics9030170
APA StyleDal Fovo, A., Mattana, S., Marchetti, M., Anichini, M., Giovannelli, A., Baria, E., Fontana, R., & Cicchi, R. (2022). Combined TPEF and SHG Imaging for the Microstructural Characterization of Different Wood Species Used in Artworks. Photonics, 9(3), 170. https://doi.org/10.3390/photonics9030170