The Maturation Pathway of Nickel Urease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
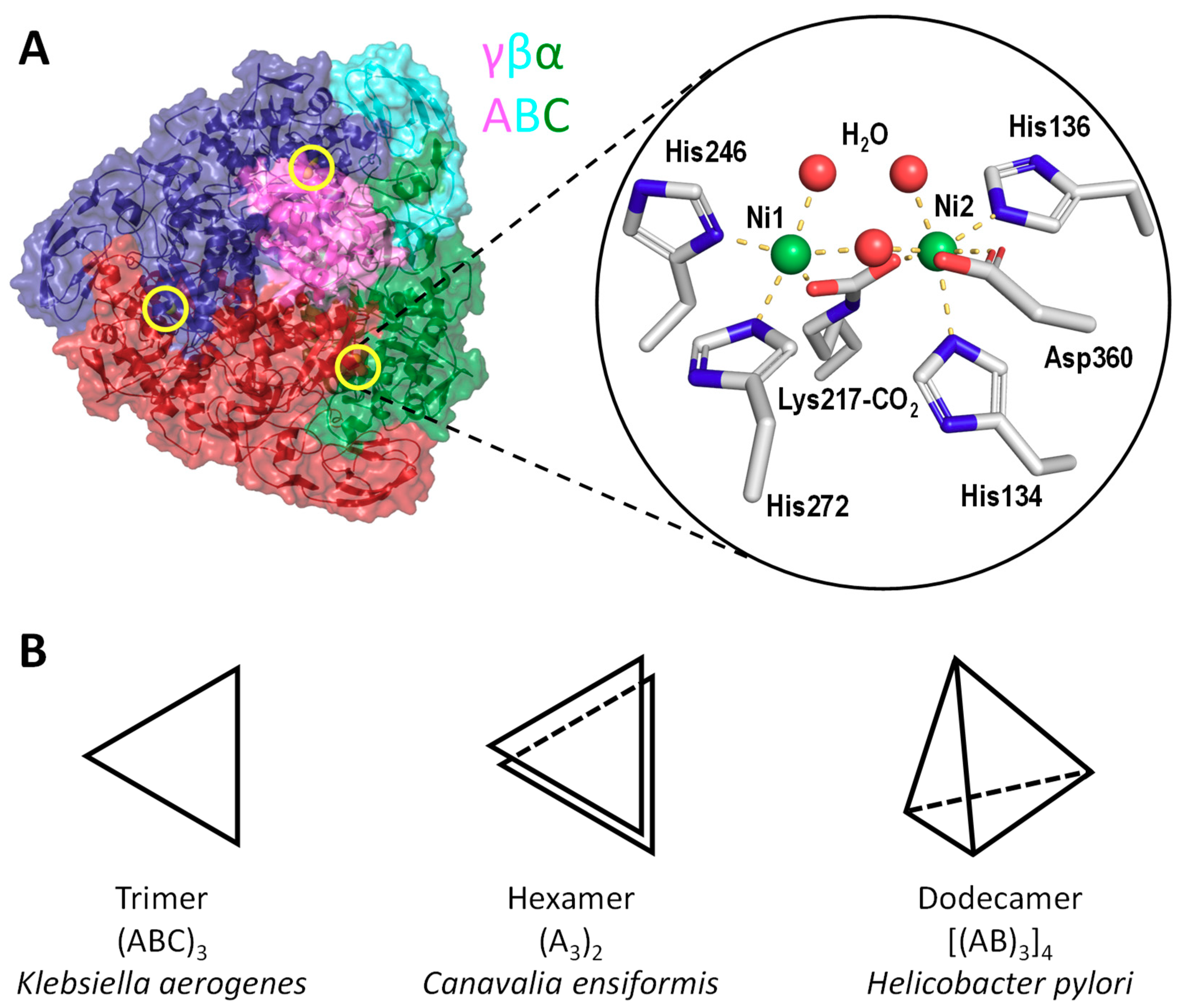
2. Structures of Urease
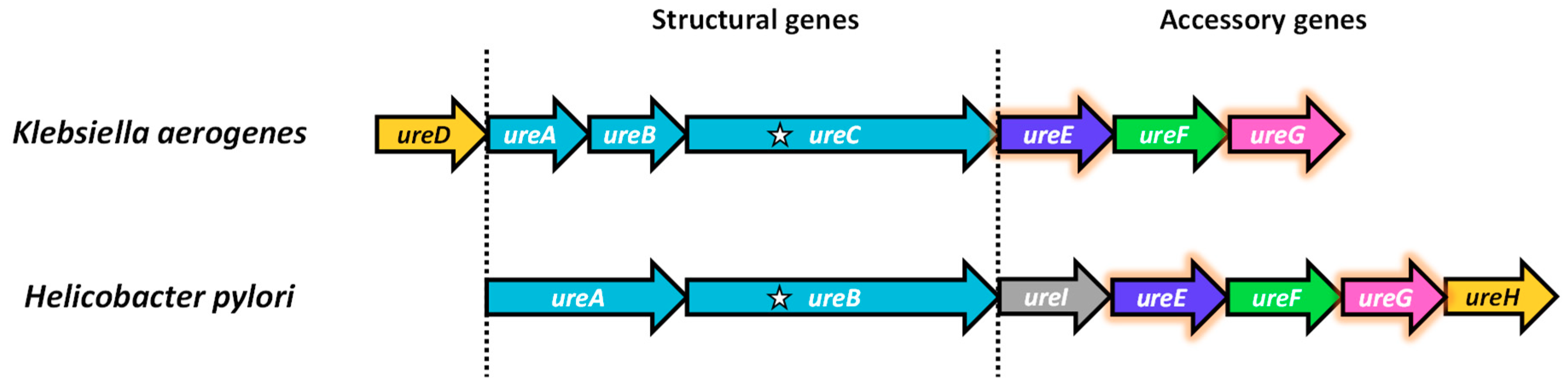
3. Genetic Studies Showed the Importance of Urease Accessory Proteins
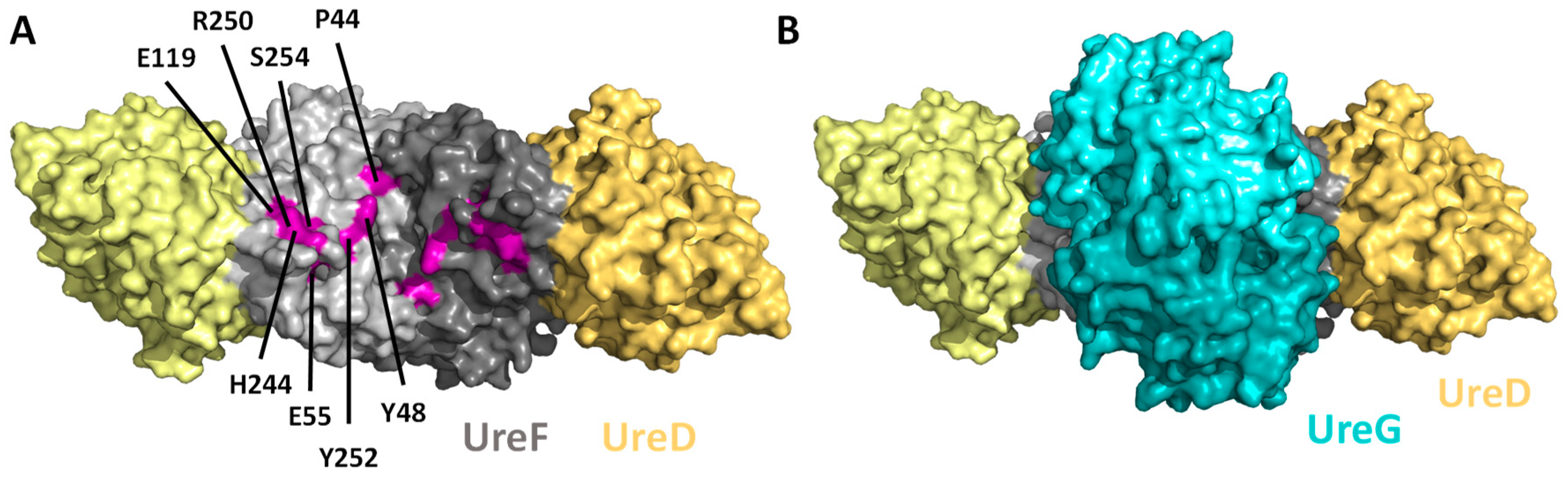
4. The Formation of UreGFD Complex
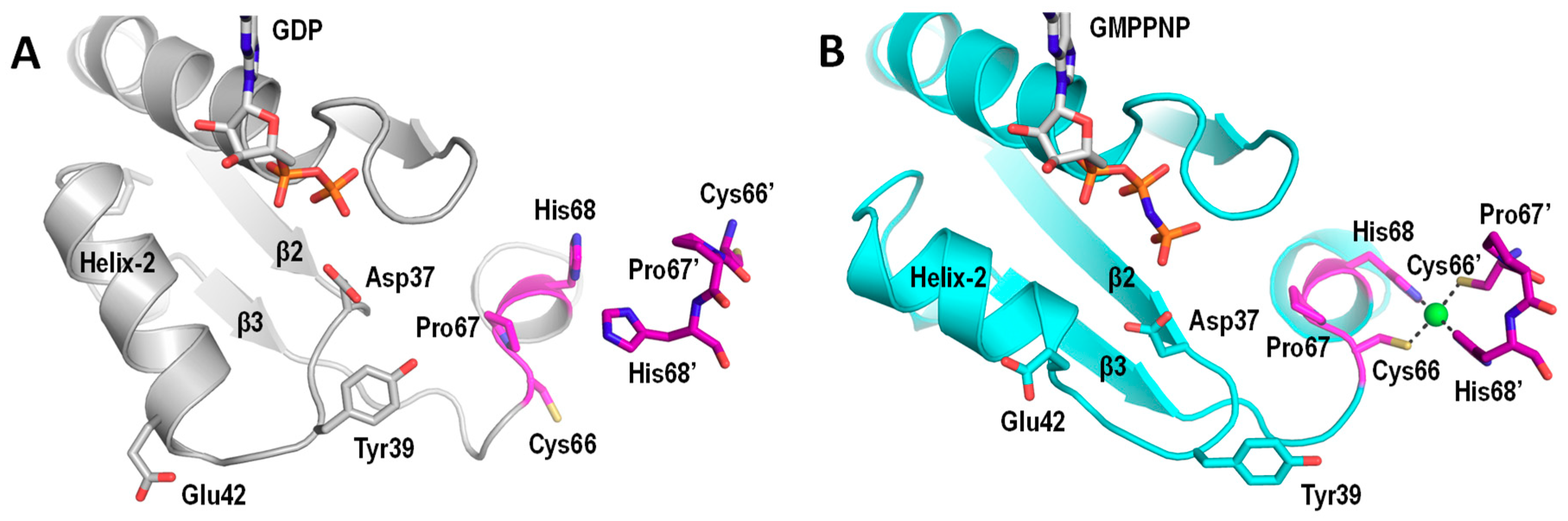
5. UreG Dissociates from the UreGFD Complex and Forms a Dimer in the Presence of Ni/GTP
6. UreG–UreE Interaction Is GTP-Dependent
7. How Urease Accessory Proteins Facilitate Urease Maturation
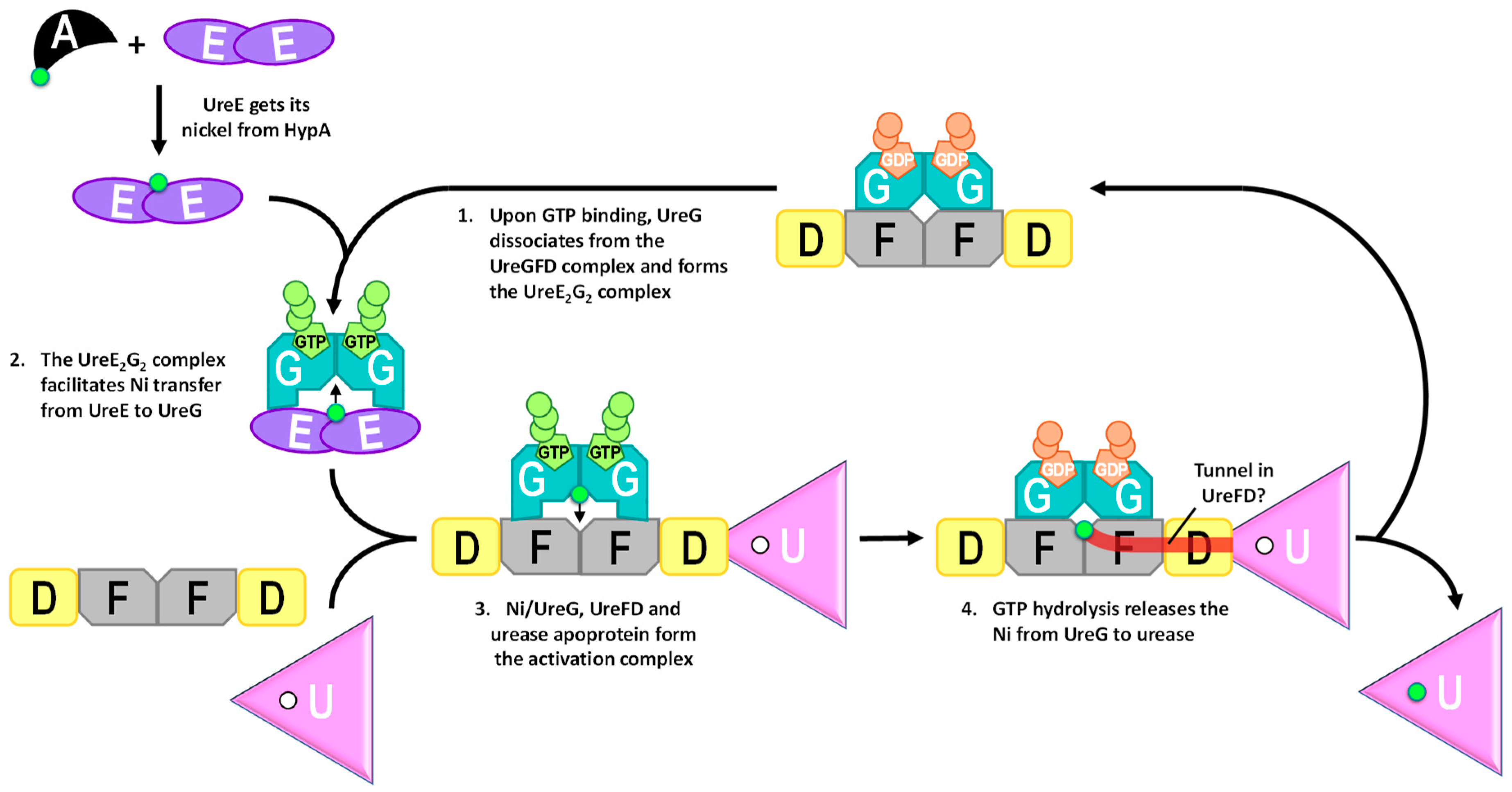
8. UreE Gets Its Nickel from Cross-Talking to the Hydrogenase Maturation Pathway
9. Urease Maturation Pathway in Plants
10. Urease Maturation Pathway Is Druggable
11. Future Perspectives
12. Conclusions
Funding
Conflicts of Interest
References
- Krajewska, B. Ureases I. Functional, Catalytic and Kinetic Properties: A Review. J. Mol. Catal. B Enzym. 2009, 59, 9–21. [Google Scholar] [CrossRef]
- Hanlon, D.P. The Distribution of Arginase and Urease in Marine Invertebrates. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1975, 52, 261–264. [Google Scholar] [CrossRef]
- Alonso-Saez, L.; Waller, A.S.; Mende, D.R.; Bakker, K.; Farnelid, H.; Yager, P.L.; Lovejoy, C.; Tremblay, J.-E.; Potvin, M.; Heinrich, F.; et al. Role for Urea in Nitrification by Polar Marine Archaea. Proc. Natl. Acad. Sci. USA 2012, 109, 17989–17994. [Google Scholar] [CrossRef] [PubMed]
- Dixon, N.E.; Gazzola, C.; Watters, J.J.; Blakeley, R.L.; Zerner, B. Jack Bean Urease (EC 3.5.1.5). A Metalloenzyme. A Simple Biological Role for Nickel? J. Am. Chem. Soc. 1975, 97, 4131–4133. [Google Scholar] [CrossRef] [PubMed]
- Sumner, J.B. The Isolation and Crystallization of the Enzyme Urease. J. Biol. Chem. 1926, 69, 435–442. [Google Scholar]
- Kao, C.-Y.; Sheu, B.-S.; Wu, J.-J. Helicobacter pylori infection: An overview of bacterial virulence facztors and pathogenesis. Biomed. J. 2016, 39, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Ishaq, S.; Nunn, L. Helicobacter pylori and Gastric Cancer: A State of the Art Review. Gastroenterol. Hepatol. Bed Bench 2015, 8, S6–S14. [Google Scholar] [PubMed]
- Narayanan, M.; Reddy, K.M.; Marsicano, E. Peptic Ulcer Disease and Helicobacter pylori Infection. Mo. Med. 2018, 115, 219–224. [Google Scholar] [PubMed]
- Carter, E.L.; Tronrud, D.E.; Taber, S.R.; Karplus, P.A.; Hausinger, R.P. Iron-Containing Urease in a Pathogenic Bacterium. Proc. Natl. Acad. Sci. USA 2011, 108, 13095–13099. [Google Scholar] [CrossRef]
- Capdevila, D.A.; Edmonds, K.A.; Giedroc, D.P. Metallochaperones and Metalloregulation in Bacteria. Essays Biochem. 2017, 61, 177–200. [Google Scholar] [CrossRef]
- Waldron, K.J.; Robinson, N.J. How Do Bacterial Cells Ensure That Metalloproteins Get the Correct Metal? Nat. Rev. Microbiol. 2009, 7, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Kappaun, K.; Piovesan, A.R.; Carlini, C.R.; Ligabue-Braun, R. Ureases: Historical Aspects, Catalytic, and Non-Catalytic Properties—A Review. J. Adv. Res. 2018, 13, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Todd, M.J.; Hausinger, R.P. Purification and Characterization of the Nickel-Containing Multicomponent Urease from Klebsiella aerogenes. J. Biol. Chem. 1987, 262, 5963–5967. [Google Scholar] [PubMed]
- Hu, L.T.; Mobley, H.L. Purification and N-Terminal Analysis of Urease from Helicobacter pylori. Infect. Immun. 1990, 58, 992–998. [Google Scholar] [PubMed]
- Blakeley, R.L.; Zerner, B. Jack Bean Urease: The First Nickel Enzyme. J. Mol. Catal. 1984, 23, 263–292. [Google Scholar] [CrossRef]
- Pearson, M.A.; Michel, L.O.; Hausinger, R.P.; Karplus, P.A. Structures of Cys319 Variants and Acetohydroxamate-Inhibited Klebsiella aerogenes Urease. Biochemistry 1997, 36, 8164–8172. [Google Scholar] [CrossRef] [PubMed]
- Jabri, E.; Carr, M.B.; Hausinger, R.P.; Karplus, P.A. The Crystal Structure of Urease from Klebsiella aerogenes. Science 1995, 268, 998–1004. [Google Scholar] [CrossRef]
- Benini, S.; Rypniewski, W.R.; Wilson, K.S.; Miletti, S.; Ciurli, S.; Mangani, S. A New Proposal for Urease Mechanism Based on the Crystal Structures of the Native and Inhibited Enzyme from Bacillus pasteurii: Why Urea Hydrolysis Costs Two Nickels. Structure 1999, 7, 205–216. [Google Scholar] [CrossRef]
- Balasubramanian, A.; Ponnuraj, K. Crystal Structure of the First Plant Urease from Jack Bean: 83 Years of Journey from Its First Crystal to Molecular Structure. J. Mol. Biol. 2010, 400, 274–283. [Google Scholar] [CrossRef]
- Ha, N.; Oh, S.; Sung, J.Y.; Cha, K.A.; Lee, M.H.; Oh, B. Supramolecular Assembly and Acid Resistance of Helicobacter pylori Urease. Nat. Struct. Mol. Biol. 2001, 8, 505–509. [Google Scholar] [CrossRef]
- Ciurli, S.; Benini, S.; Rypniewski, W.R.; Wilson, K.S.; Miletti, S.; Mangani, S. Structural Properties of the Nickel Ions in Urease: Novel Insights into the Catalytic and Inhibition Mechanisms. Coord. Chem. Rev. 1999, 190–192, 331–355. [Google Scholar] [CrossRef]
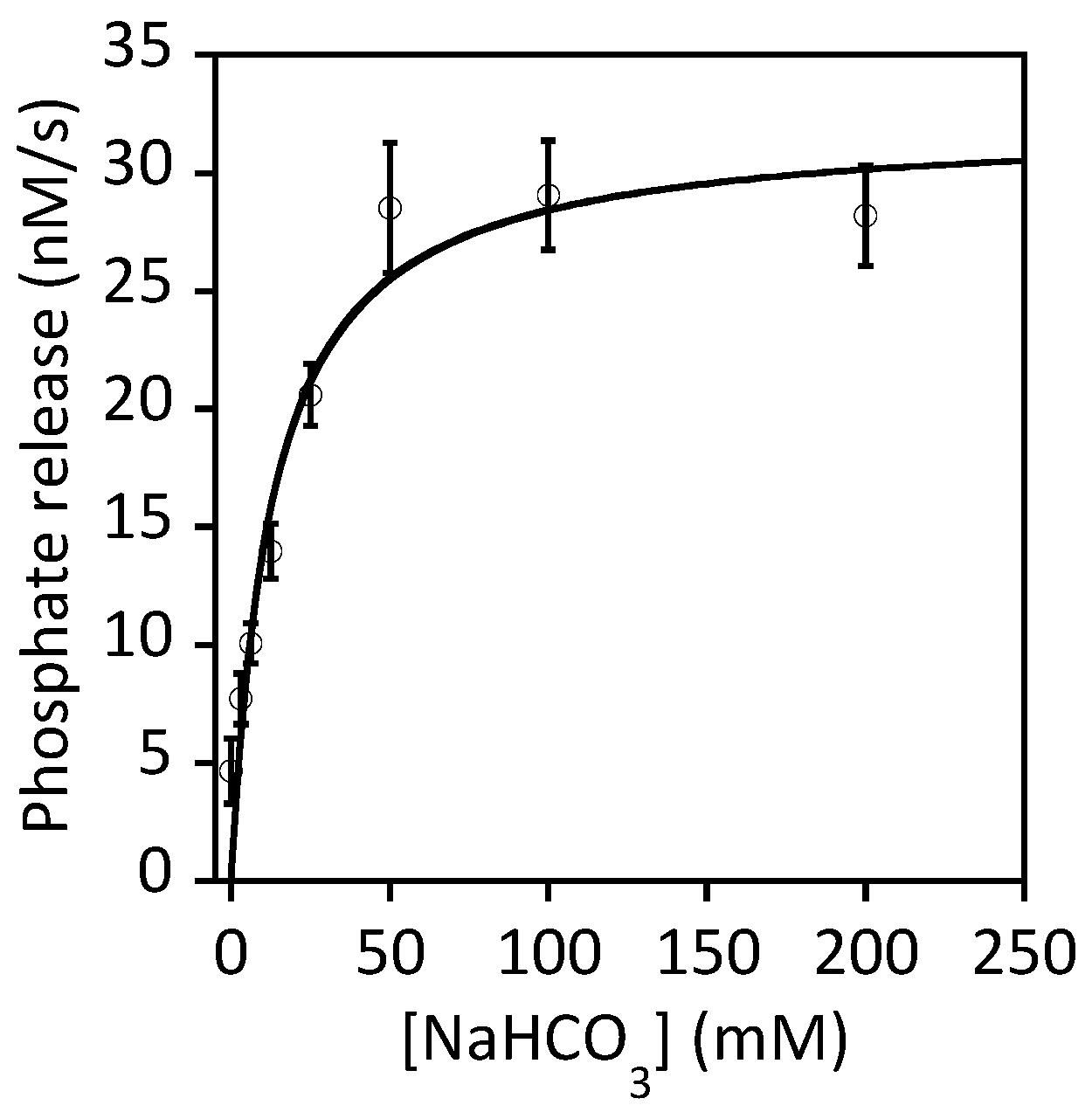
- Park, I.S.; Hausinger, R.P. Requirement of Carbon Dioxide for in vitro Assembly of the Urease Nickel Metallocenter. Science 1995, 267, 1156–1158. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.A.; Schaller, R.A.; Michel, L.O.; Karplus, P.A.; Hausinger, R.P. Chemical Rescue of Klebsiella aerogenes Urease Variants Lacking the Carbamylated-Lysine Nickel Ligand. Biochemistry 1998, 37, 6214–6220. [Google Scholar] [CrossRef] [PubMed]
- Park, I.S.; Hausinger, R.P. Metal Ion Interactions with Urease and UreD-Urease Apoproteins. Biochemistry 1996, 35, 5345–5352. [Google Scholar] [CrossRef] [PubMed]
- Mulrooney, S.B.; Hausinger, R.P. Sequence of the Klebsiella aerogenes Urease Genes and Evidence for Accessory Proteins Facilitating Nickel Incorporation. J. Bacteriol. 1990, 172, 5837–5843. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Mulrooney, S.B.; Renner, M.J.; Markowicz, Y.; Hausinger, R.P. Klebsiella aerogenes Urease Gene Cluster: Sequence of UreD and Demonstration That Four Accessory Genes (UreD, UreE, UreF, and UreG) Are Involved in Nickel Metallocenter Biosynthesis. J. Bacteriol. 1992, 174, 4324–4330. [Google Scholar] [CrossRef] [PubMed]
- Akada, J.K.; Shirai, M.; Takeuchi, H.; Tsuda, M.; Nakazawa, T. Identification of the Urease Operon in Helicobacter pylori and Its Control by mRNA Decay in Response to pH. Mol. Microbiol. 2000, 36, 1071–1084. [Google Scholar] [CrossRef]
- Cussac, V.; Ferrero, R.L.; Labigne, A. Expression of Helicobacter pylori Urease Genes in Escherichia coli Grown under Nitrogen-Limiting Conditions. J. Bacteriol. 1992, 174, 2466–2473. [Google Scholar] [CrossRef]
- Weeks, D.L.; Eskandari, S.; Scott, D.R.; Sachs, G. A H+-Gated Urea Channel: The Link between Helicobacter pylori Urease and Gastric Colonization. Science 2000, 287, 482–485. [Google Scholar] [CrossRef]
- Rektorschek, M.; Buhmann, A.; Weeks, D.; Schwan, D.; Bensch, K.W.; Eskandari, S.; Scott, D.; Sachs, G.; Melchers, K. Acid Resistance of Helicobacter pylori Depends on the UreI Membrane Protein and an Inner Membrane Proton Barrier. Mol. Microbiol. 2000, 36, 141–152. [Google Scholar] [CrossRef]
- Benoit, S.L.; Zbell, A.L.; Maier, R.J. Nickel Enzyme Maturation in Helicobacter hepaticus: Roles of Accessory Proteins in Hydrogenase and Urease Activities. Microbiology 2007, 153, 3748–3756. [Google Scholar] [CrossRef] [PubMed]
- Sriwanthana, B.; Island, M.D.; Maneval, D.; Mobley, H.L.T. Single-Step Purification of Proteus mirabilis Urease Accessory Protein UreE, a Protein with a Naturally Occurring Histidine Tail, by Nickel Chelate Affinity Chromatography. J. Bacteriol. 1994, 176, 6836–6841. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Foxall, P.A.; Russell, R.; Mobley, H.L.T. Purification of Recombinant Helicobacter pylori Urease Apoenzyme Encoded by UreA and UreB. Infect. Immun. 1992, 60, 2657–2667. [Google Scholar] [PubMed]
- Lee, M.H.; Mulrooney, S.B.; Hausinger, R.P. Purification, Characterization, and in vivo Reconstitution of Klebsiella aerogenes Urease Apoenzyme. J. Bacteriol. 1990, 172, 4427–4431. [Google Scholar] [CrossRef] [PubMed]
- Voland, P.; Weeks, D.L.; Marcus, E.A.; Prinz, C.; Sachs, G.; Scott, D. Interactions among the Seven Helicobacter pylori Proteins Encoded by the Urease Gene Cluster. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G96–G106. [Google Scholar] [CrossRef] [PubMed]
- Benoit, S.L.; Mehta, N.; Weinberg, M.V.; Maier, C.; Maier, R.J. Interaction between the Helicobacter pylori Accessory Proteins HypA and UreE Is Needed for Urease Maturation. Microbiology 2007, 153, 1474–1482. [Google Scholar] [CrossRef] [PubMed]
- Park, I.S.; Carr, M.B.; Hausinger, R.P. In vitro Activation of Urease Apoprotein and Role of UreD as a Chaperone Required for Nickel Metallocenter Assembly. Proc. Natl. Acad. Sci. USA 1994, 91, 3233–3237. [Google Scholar] [CrossRef] [PubMed]
- Carter, E.L.; Hausinger, R.P. Characterization of the Klebsiella aerogenes Urease Accessory Protein UreD in Fusion with the Maltose Binding Protein. J. Bacteriol. 2010, 192, 2294–2304. [Google Scholar] [CrossRef]
- Moncrief, M.B.C.; Hausinger, R.P. Purification and Activation Properties of UreD-UreF-Urease Apoprotein Complexes. J. Bacteriol. 1996, 178, 5417–5421. [Google Scholar] [CrossRef][Green Version]
- Kim, J.K.; Mulrooney, S.B.; Hausinger, R.P. The UreEF Fusion Protein Provides a Soluble and Functional Form of the UreF Urease Accessory Protein. J. Bacteriol. 2006, 188, 8413–8420. [Google Scholar] [CrossRef]
- Soriano, A.; Hausinger, R.P. GTP-Dependent Activation of Urease Apoprotein in Complex with the UreD, UreF, and UreG Accessory Proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 11140–11144. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Hausinger, R.P. Evidence for the Presence of Urease Apoprotein Complexes Containing UreD, UreF, and UreG in Cells That Are Competent for in vivo Enzyme Activation. J. Bacteriol. 1995, 177, 1947–1951. [Google Scholar] [CrossRef] [PubMed]
- Fong, Y.H.; Wong, H.C.; Yuen, M.H.; Lau, P.H.; Chen, Y.W.; Wong, K.-B. Structure of UreG/UreF/UreH Complex Reveals How Urease Accessory Proteins Facilitate Maturation of Helicobacter pylori Urease. PLoS Biol. 2013, 11, e1001678. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, M.A.; Han, L.; Zhong, Y.; Boer, J.L.; Ruotolo, B.T.; Hausinger, R.P. Analysis of a Soluble (UreD:UreF:UreG)2 Accessory Protein Complex and Its Interactions with Klebsiella aerogenes Urease by Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2013, 24, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Moncrief, M.B.C.; Hausinger, R.P. Characterization of UreG, Identification of a UreD-UreF-UreG Complex, and Evidence Suggesting That a Nucleotide-Binding Site in UreG Is Required for in vivo Metallocenter Assembly of Klebsiella aerogenes Urease. J. Bacteriol. 1997, 179, 4081–4086. [Google Scholar] [CrossRef][Green Version]
- Boer, J.L.; Quiroz-Valenzuela, S.; Anderson, K.L.; Hausinger, R.P. Mutagenesis of Klebsiella aerogenes UreG to Probe Nickel Binding and Interactions with Other Urease-Related Proteins. Biochemistry 2010, 49, 5859–5869. [Google Scholar] [CrossRef]
- Rain, J.C.; Selig, L.; De Reuse, H.; Battaglia, V.; Reverdy, C.; Simon, S.; Lenzen, G.; Petel, F.; Wojcik, J.; Schächter, V.; et al. The Protein-Protein Interaction Map of Helicobacter pylori. Nature 2001, 409, 211–215. [Google Scholar] [CrossRef]
- Fong, Y.H.; Wong, H.C.; Chuck, C.P.; Chen, Y.W.; Sun, H.; Wong, K.-B. Assembly of Preactivation Complex for Urease Maturation in Helicobacter pylori: Crystal Structure of UreF-UreH Protein Complex. J. Biol. Chem. 2011, 286, 43241–43249. [Google Scholar] [CrossRef]
- Lam, R.; Romanov, V.; Johns, K.; Battaile, K.P.; Wu-Brown, J.; Guthrie, J.L.; Hausinger, R.P.; Pai, E.F.; Chirgadze, N.Y. Crystal Structure of a Truncated Urease Accessory Protein UreF from Helicobacter pylori. Proteins 2010, 78, 2839–2848. [Google Scholar] [CrossRef]
- Boer, J.L.; Hausinger, R.P. Klebsiella aerogenes UreF: Identification of the UreG Binding Site and Role in Enhancing the Fidelity of Urease Activation. Biochemistry 2012, 51, 2298–2308. [Google Scholar] [CrossRef]
- Yuen, M.H.; Fong, Y.H.; Nim, Y.S.; Lau, P.H.; Wong, K.-B. Structural Insights into how GTP-Dependent Conformational Changes in a Metallochaperone UreG Facilitate Urease Maturation. Proc. Natl. Acad. Sci. USA 2017, 114, E10890–E10898. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, H.; Lai, T.-P.; Sun, H. UreE-UreG Complex Facilitates Nickel Transfer and Preactivates GTPase of UreG in Helicobacter pylori. J. Biol. Chem. 2015, 290, 12474–12485. [Google Scholar] [CrossRef] [PubMed]
- Leipe, D.D.; Wolf, Y.I.; Koonin, E.V.; Aravind, L. Classification and Evolution of P-Loop GTPases and Related ATPases. J. Mol. Biol. 2002, 317, 41–72. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, B.; Turano, P.; Musiani, F.; Neyroz, P.; Ciurli, S. Zn2+-Linked Dimerization of UreG from Helicobacter pylori, a Chaperone Involved in Nickel Trafficking and Urease Activation. Proteins Struct. Funct. Bioinform. 2009, 74, 222–239. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, B.; Stola, M.; Musiani, F.; De Vriendt, K.; Samyn, B.; Devreese, B.; Van Beeumen, J.; Turano, P.; Dikiy, A.; Bryant, D.A.; et al. UreG, a Chaperone in the Urease Process, Is an Intrinsically Unstructured Protein Binding a Single Zn2+ Ion. J. Biol. Chem. 2005, 280, 4684–4695. [Google Scholar] [CrossRef] [PubMed]
- Real-Guerra, R.; Staniscuaski, F.; Zambelli, B.; Musiani, F.; Ciurli, S.; Carlini, C.R. Biochemical and Structural Studies on Native and Recombinant Glycine max UreG: A Detailed Characterization of a Plant Urease Accessory Protein. Plant Mol. Biol. 2012, 78, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Miraula, M.; Ciurli, S.; Zambelli, B. Intrinsic Disorder and Metal Binding in UreG Proteins from Archae Hyperthermophiles: GTPase Enzymes Involved in the Activation of Ni(II) Dependent Urease. J. Biol. Inorg. Chem. 2015, 20, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Martin-Diaconescu, V.; Bellucci, M.; Musiani, F.; Ciurli, S.; Maroney, M.J. Unraveling the Helicobacter pylori UreG Zinc Binding Site Using X-Ray Absorption Spectroscopy (XAS) and Structural Modeling. J. Biol. Inorg. Chem. 2012, 17, 353–361. [Google Scholar] [CrossRef]
- Xia, W.; Li, H.; Yang, X.; Wong, K.B.; Sun, H. Metallo-GTPase HypB from Helicobacter pylori and Its Interaction with Nickel Chaperone Protein HypA. J. Biol. Chem. 2012, 287, 6753–6763. [Google Scholar] [CrossRef] [PubMed]
- Lacasse, M.J.; Douglas, C.D.; Zamble, D.B. Mechanism of Selective Nickel Transfer from HypB to HypA, Escherichia coli [NiFe]-Hydrogenase Accessory Proteins. Biochemistry 2016, 55, 6821–6831. [Google Scholar] [CrossRef] [PubMed]
- Gasper, R.; Scrima, A.; Wittinghofer, A. Structural Insights into HypB, a GTP-Binding Protein That Regulates Metal Binding. J. Biol. Chem. 2006, 281, 27492–27502. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Ngu, T.T.; Kaluarachchi, H.; Zamble, D.B. Relationship between the GTPase, Metal-Binding, and Dimerization Activities of E. Coli HypB. J. Inorg. Biochem. 2011, 16, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.H.; Lee, K.M.; Wong, K.B. Interaction between Hydrogenase Maturation Factors HypA and HypB Is Required for [NiFe]-Hydrogenase Maturation. PLoS ONE 2012, 7, e32592. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.H.; Li, T.; Wong, C.O.; Wong, K.B. Structural Basis for GTP-Dependent Dimerization of Hydrogenase Maturation Factor HypB. PLoS ONE 2012, 7, e30547. [Google Scholar] [CrossRef] [PubMed]
- Remaut, H.; Safarov, N.; Ciurli, S.; Van Beeumen, J. Structural Basis for Ni2+ Transport and Assembly of the Urease Active Site by the Metallochaperone UreE from Bacillus pasteurii. J. Biol. Chem. 2001, 276, 49365–49370. [Google Scholar] [CrossRef]
- Song, H.K.; Mulrooney, S.B.; Huber, R.; Hausinger, R.P. Crystal Structure of Klebsiella aerogenes UreE, a Nickel-Binding Metallochaperone for Urease Activation. J. Biol. Chem. 2001, 276, 49359–49364. [Google Scholar] [CrossRef]
- Banaszak, K.; Martin-Diaconescu, V.; Bellucci, M.; Zambelli, B.; Rypniewski, W.; Maroney, M.J.; Ciurli, S. Crystallographic and X-Ray Absorption Spectroscopic Characterization of Helicobacter pylori UreE Bound to Ni2+ and Zn2+ Reveals a Role for the Disordered C-Terminal Arm in Metal Trafficking. Biochem. J. 2012, 441, 1017–1026. [Google Scholar] [CrossRef]
- Shi, R.; Munger, C.; Asinas, A.; Benoit, S.L.; Miller, E.; Matte, A.; Maier, R.J.; Cygler, M. Crystal Structures of apo and Metal-Bound Forms of the UreE Protein from Helicobacter pylori: Role of Multiple Metal Binding Sites. Biochemistry 2010, 49, 7080–7088. [Google Scholar] [CrossRef]
- Stola, M.; Musiani, F.; Mangani, S.; Turano, P.; Safarov, N.; Zambelli, B.; Ciurli, S. The Nickel Site of Bacillus pasteurii UreE, a Urease Metallo-Chaperone, as Revealed by Metal-Binding Studies and X-Ray Absorption Spectroscopy. Biochemistry 2006, 45, 6495–6509. [Google Scholar] [CrossRef]
- Lee, M.H.; Pankratz, H.S.; Wang, S.; Scott, R.A.; Finnegan, M.G.; Johnson, M.K.; Ippolito, J.A.; Christianson, D.W.; Hausinger, R.P. Purification and Characterization of Klebsiella aerogenes UreE Protein: A Nickel-binding Protein That Functions in Urease Metallocenter Assembly. Protein Sci. 1993, 2, 1042–1052. [Google Scholar] [CrossRef]
- Bellucci, M.; Zambelli, B.; Musiani, F.; Turano, P.; Ciurli, S. Helicobacter pylori UreE, a Urease Accessory Protein: Specific Ni2+- and Zn2+-Binding Properties and Interaction with Its Cognate UreG. Biochem. J. 2009, 422, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Grossoehme, N.E.; Mulrooney, S.B.; Hausinger, R.P.; Wilcox, D.E. Thermodynamics of Ni2+, Cu2+, and Zn2+ Binding to the Urease Metallochaperone. Biochemistry 2007, 46, 10506–10516. [Google Scholar] [CrossRef] [PubMed]
- Won, H.; Lee, Y.; Kim, J.; Shin, I.S.; Lee, M.H.; Lee, B. Structural Characterization of the Nickel-Binding Properties of Bacillus pasteurii Urease Accessory Protein (Ure)E in Solution. J. Biol. Chem. 2004, 279, 17466–17472. [Google Scholar] [CrossRef] [PubMed]
- Ciurli, S.; Safarov, N.; Miletti, S.; Dikiy, A.; Christensen, S.K.; Kornetzky, K.; Bryant, D.A.; Vandenberghe, I.; Devreese, B.; Samyn, B.; et al. Molecular Characterization of Bacillus pasteurii UreE, a Metal-Binding Chaperone for the Assembly of the Urease Active Site. J. Biol. Inorg. Chem. 2002, 7, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Colpas, G.J.; Hausinger, R.P. In vivo and in citro Kinetics of Metal Transfer by the Klebsiella aerogenes Urease Nickel Metallochaperone, UreE. J. Biol. Chem. 2000, 275, 10731–10737. [Google Scholar] [CrossRef] [PubMed]
- Colpas, G.J.; Brayman, T.G.; Ming, L.J.; Hausinger, R.P. Identification of Metal-Binding Residues in the Klebsiella aerogenes Urease Nickel Metallochaperone, UreE. Biochemistry 1999, 38, 4078–4088. [Google Scholar] [CrossRef] [PubMed]
- Brayman, T.G.; Hausinger, R.P. Purification, Characterization, and Functional Analysis of a Truncated Klebsiella aerogenes UreE Urease Accessory Protein Lacking the Histidine-Rich Carboxyl Terminus. J. Bacteriol. 1996, 178, 5410–5416. [Google Scholar] [CrossRef] [PubMed]
- Mulrooney, S.B.; Ward, S.K.; Hausinger, R.P. Purification and Properties of the Klebsiella aerogenes UreE Metal-Binding Domain, a Functional Metallochaperone of Urease. J. Bacteriol. 2005, 187, 3581–3585. [Google Scholar] [CrossRef]
- Musiani, F.; Zambelli, B.; Stola, M.; Ciurli, S. Nickel Trafficking: Insights into the Fold and Function of UreE, a Urease Metallochaperone. J. Inorg. Biochem. 2004, 98, 803–813. [Google Scholar] [CrossRef]
- Merloni, A.; Dobrovolska, O.; Zambelli, B.; Agostini, F.; Bazzani, M.; Musiani, F.; Ciurli, S. Molecular Landscape of the Interaction between the Urease Accessory Proteins UreE and UreG. Biochim. Biophys. Acta Proteins Proteom. 2014, 1844, 1662–1674. [Google Scholar] [CrossRef]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS Biomolecular Solvation Software Suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, M.A.; Wang, B.; Feig, M.; Hausinger, R.P. Mutational and Computational Evidence That a Nickel-Transfer Tunnel in UreD Is Used for Activation of Klebsiella aerogenes Urease. Biochemistry 2015, 54, 6392–6401. [Google Scholar] [CrossRef] [PubMed]
- Soriano, A.; Colpas, G.J.; Hausinger, R.P. UreE Stimulation of GTP-Dependent Urease Activation in the UreD-UreF-UreG-Urease Apoprotein Complex. Biochemistry 2000, 39, 12435–12440. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Kuchar, J.; Hausinger, R.P. Chemical Cross-Linking and Mass Spectrometric Identification of Sites of Interaction for UreD, UreF, and Urease. J. Biol. Chem. 2004, 279, 15305–15313. [Google Scholar] [CrossRef]
- Musiani, F.; Gioia, D.; Masetti, M.; Falchi, F.; Cavalli, A.; Recanatini, M.; Ciurli, S. Protein Tunnels: The Case of Urease Accessory Proteins. J. Chem. Theory Comput. 2017, 13, 2322–2331. [Google Scholar] [CrossRef]
- Zambelli, B.; Berardi, A.; Martin-Diaconescu, V.; Mazzei, L.; Musiani, F.; Maroney, M.J.; Ciurli, S. Nickel Binding Properties of Helicobacter pylori UreF, an Accessory Protein in the Nickel-Based Activation of Urease. J. Biol. Inorg. Chem. 2014, 19, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Baykov, A.A.; Evtushenko, O.A.; Avaeva, S.M. A Malachite Green Procedure for Orthophosphate Determination and Its Use in Alkaline Phosphatase-Based Enzyme Immunoassay. Anal. Biochem. 1988, 171, 266–270. [Google Scholar] [CrossRef]
- Maier, T.; Lottspeich, F.; Böck, A. GTP Hydrolysis by HypB Is Essential for Nickel Insertion into Hydrogenases of Escherichia coli. Eur. J. Biochem. 1995, 230, 133–138. [Google Scholar] [CrossRef]
- Olson, J.W.; Fu, C.; Maier, R.J. The HypB Protein from Bradyrhizobium japonicum can Store Nickel and Is Required for the Nickel-dependent Transcriptional Regulation of Hydrogenase. Mol. Microbiol. 1997, 24, 119–128. [Google Scholar] [CrossRef]
- Olson, J.W.; Mehta, N.S.; Maier, R.J. Requirement of Nickel Metabolism Proteins HypA and HypB for Full Activity of Both Hydrogenase and Urease in Helicobacter pylori. Mol. Microbiol. 2001, 39, 176–182. [Google Scholar] [CrossRef]
- Hube, M.; Blokesch, M.; Böck, A. Network of Hydrogenase Maturation in Escherichia coli: Role of Accessory Proteins HypA and HybF. J. Bacteriol. 2002, 184, 3879–3885. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, D.; Gutekunst, K.; Klissenbauer, M.; Schulz-Friedrich, R.; Appel, J. Mutagenesis of Hydrogenase Accessory Genes of Synechocystis sp. PCC 6803: Additional Homologues of hypA and hypB Are Not Active in Hydrogenase Maturation. FEBS J. 2006, 273, 4516–4527. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.; Jacobi, A.; Sauter, M.; Böck, A. The Product of the HypB Gene, Which Is Required for Nickel Incorporation into Hydrogenases, Is a Novel Guanine Nucleotide-Binding Protein. J. Bacteriol. 1993, 175, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, A.; Rossmann, R.; Böck, A. The hyp Operon Gene Products Are Required for the Maturation of Catalytically Active Hydrogenase Isoenzymes in Escherichia coli. Arch. Microbiol. 1992, 158, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Lutz, S.; Jacobi, A.; Schlensog, V.; Böhm, R.; Sawers, G.; Böck, A. Molecular Characterization of an Operon (hyp) Necessary for the Activity of the Three Hydrogenase Isoenzymes in Escherichia coli. Mol. Microbiol. 1991, 5, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Tomb, J.F.; White, O.; Kerlavage, A.R.; Clayton, R.A.; Sutton, G.G.; Fleischmann, R.D.; Ketchum, K.A.; Klenk, H.P.; Gill, S.; Brian, A.D. The Complete Genome Sequence of the Gastric Pathogen Helicobacter pylori. Nature 1997, 388, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Spronk, C.A.E.M.; Żerko, S.; Górka, M.; Koźmiński, W.; Bardiaux, B.; Zambelli, B.; Musiani, F.; Piccioli, M.; Basak, P.; Blum, F.C.; et al. Structure and Dynamics of Helicobacter pylori Nickel-chaperone HypA: An Integrated Approach Using NMR Spectroscopy, Functional Assays and Computational Tools. JBIC J. Biol. Inorg. Chem. 2018, 23, 1309–1330. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Li, H.; Sze, K.H.; Sun, H. Structure of a Nickel Chaperone, HypA, from Helicobacter pylori Reveals Two Distinct Metal Binding Sites. J. Am. Chem. Soc. 2009, 131, 10031–10040. [Google Scholar] [CrossRef]
- Mehta, N.; Olson, J.W.; Maier, R.J. Characterization of Helicobacter pylori Nickel Metabolism Accessory Proteins Needed for Maturation of Both Urease and Hydrogenase. J. Bacteriol. 2003, 185, 726–734. [Google Scholar] [CrossRef]
- Herbst, R.W.; Perovic, I.; Martin-diaconescu, V.; Brien, K.O.; Chivers, P.T.; Pochapsky, S.S.; Pochapsky, T.C.; Maroney, M.J. Communication between the Zinc and Nickel Sites in Dimeric HypA: Metal Recognition and pH Sensing. J. Am. Chem. Soc. 2010, 132, 10338–10351. [Google Scholar] [CrossRef]
- Hu, H.Q.; Johnson, R.C.; Merrell, D.S.; Maroney, M.J. Nickel Ligation of the N-Terminal Amine of HypA Is Required for Urease Maturation in Helicobacter pylori. Biochemistry 2017, 56, 1105–1116. [Google Scholar] [CrossRef] [PubMed]
- Atanassova, A.; Zamble, D.B. Escherichia coli HypA Is a Zinc Metalloprotein with a Weak Affinity for Nickel. J. Bacteriol. 2005, 187, 4689–4697. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Arai, T.; Matsumi, R.; Atomi, H.; Imanaka, T.; Miki, K. Crystal Structure of HypA, a Nickel-Binding Metallochaperone for [NiFe] Hydrogenase Maturation. J. Mol. Biol. 2009, 394, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.C.; Herbst, R.W.; Iwig, J.S.; Chivers, P.T.; Maroney, M.J. A Dynamic Zn Site in Helicobacter pylori HypA: A Potential Mechanism for Metal-Specific Protein Activity. J. Am. Chem. Soc. 2007, 129, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.C.; Hu, H.Q.; Merrell, D.S.; Maroney, M.J. Dynamic HypA Zinc Site Is Essential for Acid Viability and Proper Urease Maturation in Helicobacter pylori. Metallomics 2015, 7, 674–682. [Google Scholar] [CrossRef]
- Stingl, K.; Schauer, K.; Ecobichon, C.; Labigne, A.; Lenormand, P.; Rousselle, J.-C.; Namane, A.; de Reuse, H. In vivo Interactome of Helicobacter pylori Urease Revealed by Tandem Affinity Purification. Mol. Cell. Proteom. 2008, 7, 2429–2441. [Google Scholar] [CrossRef]
- Hu, H.Q.; Huang, H.T.; Maroney, M.J. The Helicobacter pylori HypA·UreE2 Complex Contains a Novel High-Affinity Ni(II)-Binding Site. Biochemistry 2018, 57, 2932–2942. [Google Scholar] [CrossRef]
- Yang, X.; Li, H.; Cheng, T.; Xia, W.; Lai, Y.-T.; Sun, H. Nickel Translocation between Metallochaperones HypA and UreE in Helicobacter pylori. Metallomics 2014, 6, 40–42. [Google Scholar] [CrossRef]
- Benoit, S.L.; McMurry, J.L.; Hill, S.A.; Maier, R.J. Helicobacter pylori Hydrogenase Accessory Protein HypA and Urease Accessory Protein UreG Compete with Each Other for UreE Recognition. Biochim. Biophys. Acta-Gen. Subj. 2012, 1820, 1519–1525. [Google Scholar] [CrossRef]
- Douglas, C.D.; Ngu, T.T.; Kaluarachchi, H.; Zamble, D.B. Metal Transfer within the Escherichia coli HypB−HypA Complex of Hydrogenase Accessory Proteins. Biochemistry 2013, 52, 6030–6039. [Google Scholar] [CrossRef]
- Kaluarachchi, H.; Zhang, J.W.; Zamble, D.B. Escherichia coli SlyD, More Than a Ni(II) Reservoir. Biochemistry 2011, 50, 10761–10763. [Google Scholar] [CrossRef]
- Cheng, T.; Li, H.; Yang, X.; Xia, W.; Sun, H. Interaction of SlyD with HypB of Helicobacter pylori Facilitates Nickel Trafficking. Metallomics 2013, 5, 804–807. [Google Scholar] [CrossRef]
- Zhang, J.W.; Butland, G.; Greenblatt, J.F.; Emili, A.; Zamble, D.B. A Role for SlyD in the Escherichia coli Hydrogenase Biosynthetic Pathway. J. Biol. Chem. 2005, 280, 4360–4366. [Google Scholar] [CrossRef]
- Leach, M.R.; Jie, W.Z.; Zamble, D.B. The Role of Complex Formation between the Escherichia coli Hydrogenase Accessory Factors HypB and SlyD. J. Biol. Chem. 2007, 282, 16177–16186. [Google Scholar] [CrossRef]
- Lacasse, M.J.; Zamble, D.B. [NiFe]-Hydrogenase Maturation. Biochemistry 2016, 55, 1689–1701. [Google Scholar] [CrossRef]
- Zeer-wanklyn, C.J.; Zamble, D.B. Microbial Nickel: Cellular Uptake and Delivery to Enzyme Centers. Curr. Opin. Chem. Biol. 2017, 37, 80–88. [Google Scholar] [CrossRef]
- Witte, C.-P. Urea Metabolism in Plants. Plant Sci. 2011, 180, 431–438. [Google Scholar] [CrossRef]
- Polacco, J.C.; Mazzafera, P.; Tezotto, T. Opinion—Nickel and Urease in Plants: Still Many Knowledge Gaps. Plant Sci. 2013, 199–200, 79–90. [Google Scholar] [CrossRef]
- Cao, F.-Q.; Werner, A.K.; Dahncke, K.; Romeis, T.; Liu, L.-H.; Witte, C.-P. Identification and Characterization of Proteins Involved in Rice Urea and Arginine Catabolism. Plant Physiol. 2010, 154, 98–108. [Google Scholar] [CrossRef]
- Myrach, T.; Zhu, A.; Witte, C.-P. The Assembly of the Plant Urease Activation Complex and the Essential Role of the Urease Accessory Protein G (UreG) in Delivery of Nickel to Urease. J. Biol. Chem. 2017, 292, 14556–14565. [Google Scholar] [CrossRef]
- Witte, C.-P.; Rosso, M.G.; Romeis, T. Identification of Three Urease Accessory Proteins that are Required for Urease Activation in Arabidopsis. Plant Physiol. 2005, 139, 1155–1162. [Google Scholar] [CrossRef]
- Freyermuth, S.K.; Bacanamwo, M.; Polacco, J.C. The Soybean Eu3 Gene Encodes an Ni-Binding Protein Necessary for Urease Activity. Plant J. 2000, 21, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, J.C. The Emerging Role of Urease as a General Microbial Virulence Factor. PLoS Pathog. 2014, 10, e1004062. [Google Scholar] [CrossRef] [PubMed]
- Safavi, M.; Sabourian, R.; Foroumadi, A. Treatment of Helicobacter pylori Infection: Current and Future Insights. World J. Clin. Cases 2016, 4, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Koohi-moghadam, M.; Wang, R.; Chang, Y.; Woo, P.C.Y.; Wang, J.; Li, H.; Sun, H. Metallochaperone UreG Serves as a New Target for Design of Urease Inhibitor: A Novel Strategy for Development of Antimicrobials. PLoS Biol. 2018, 16, e2003887. [Google Scholar] [CrossRef] [PubMed]
- Eschweiler, J.D.; Farrugia, M.A.; Dixit, S.M.; Hausinger, R.P.; Ruotolo, B.T. A Structural Model of the Urease Activation Complex Derived from Ion Mobility-Mass Spectrometry and Integrative Modeling. Structure 2018, 26, 599–606.e3. [Google Scholar] [CrossRef] [PubMed]
- Ligabue-Braun, R.; Real-Guerra, R.; Carlini, C.R.; Verli, H. Evidence-Based Docking of the Urease Activation Complex. J. Biomol. Struct. Dyn. 2013, 31, 854–861. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nim, Y.S.; Wong, K.-B. The Maturation Pathway of Nickel Urease. Inorganics 2019, 7, 85. https://doi.org/10.3390/inorganics7070085
Nim YS, Wong K-B. The Maturation Pathway of Nickel Urease. Inorganics. 2019; 7(7):85. https://doi.org/10.3390/inorganics7070085
Chicago/Turabian StyleNim, Yap Shing, and Kam-Bo Wong. 2019. "The Maturation Pathway of Nickel Urease" Inorganics 7, no. 7: 85. https://doi.org/10.3390/inorganics7070085
APA StyleNim, Y. S., & Wong, K.-B. (2019). The Maturation Pathway of Nickel Urease. Inorganics, 7(7), 85. https://doi.org/10.3390/inorganics7070085