

Characterization of SHH, SOX3, WNT3A and WNT9B Proteins in Human Non-Syndromic Cleft Lip and Palate Tissue




Abstract
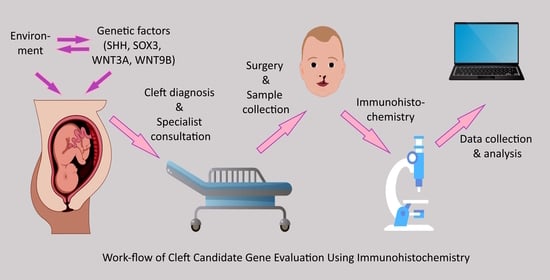
:
1. Introduction
2. Materials and Methods
2.1. Characterization of Patient Groups and Tissue Material
2.2. Immunohistochemistry and Slide Evaluation
2.3. Statistical Methods
3. Results
3.1. SHH Immunohistochemistry
3.2. SOX3 Immunohistochemistry
3.3. WNT3A Immunohistochemistry
3.4. WNT9B Immunohistochemistry
3.5. Correlations in Cleft-Affected Tissue
3.5.1. Correlations in Controls
3.5.2. Correlations in UCL Tissue
3.5.3. Correlations in BCL Tissue
3.5.4. Correlations in CP Tissue
4. Discussion
4.1. Immunohistochemistry
4.2. Correlations
4.2.1. Control Tissue Correlations
4.2.2. UCL Tissue Correlations
4.2.3. BCL Tissue Correlations
4.2.4. CP Tissue Correlations
4.3. Limitations of the Study
5. Conclusions
- The statistically significant decrease of SHH in bilateral cleft lip connective tissue and cleft palate tissue could imply that SHH dysfunction might be more associated with the morphogenesis and tissue growth of these specific types of clefts postnatally and less with unilateral cleft lip.
- The statistically significant decrease of SOX3, WNT3A and WNT9B proteins in all evaluated non-syndromic cleft-affected tissue types could indicate similarities in cleft morphogenesis and growth mechanisms within the postnatal cleft lip and palate tissue, possibly due to interlinked signaling pathway disruption, such as disrupted WNT signaling.
- The presence of similar correlations between SHH, SOX3, WNT3A, and WNT9B protein-containing cells within different cleft-affected tissue types could indicate the existence of similar signaling mechanisms and interactions within each separate cleft type postnatally, while the possible differences in correlations between cleft types might be affected by other regulatory factors mediating and modifying SHH, SOX3, WNT3A, and WNT9B function.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Impellizzeri, A.; Giannantoni, I.; Polimeni, A.; Barbato, E.; Galluccio, G. Epidemiological Characteristic of Orofacial Clefts and Its Associated Congenital Anomalies: Retrospective Study. BMC Oral Health 2019, 19, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.K.; Bui, A.H.; Taioli, E. Epidemiology of Cleft Lip and Palate. In Designing Strategies for Cleft Lip and Palate Care; InTech: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Nasreddine, G.; El Hajj, J.; Ghassibe-Sabbagh, M. Orofacial Clefts Embryology, Classification, Epidemiology, and Genetics. Mutat. Res. Rev. Mutat. Res. 2021, 787, 108373. [Google Scholar] [CrossRef] [PubMed]
- Alois, C.I.; Ruotolo, R.A. An Overview of Cleft Lip and Palate. J. Am. Acad. Physician Assist. 2020, 33, 17–20. [Google Scholar] [CrossRef]
- Martinelli, M.; Palmieri, A.; Carinci, F.; Scapoli, L. Non-Syndromic Cleft Palate: An Overview on Human Genetic and Environmental Risk Factors. Front. Cell Dev. Biol. 2020, 8, 592271. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, K.; Zhang, S.; Sun, B.; Garland, M.A.; Ji, Y.; Zhou, C.J. Genetics and Signaling Mechanisms of Orofacial Clefts. Birth Defects Res. 2020, 112, 1588–1634. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Jia, Z.; Shi, Y.; Du, Q.; Shi, J.; Wang, Z.; Mou, Y.; Wang, Q.; Zhang, B.; Wang, Q.; et al. Genetic Factors Define CPO and CLO Subtypes of Nonsyndromicorofacial Cleft. PLoS Genet. 2019, 15, e1008357. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Dai, Y.; Iwata, J.; Zhao, Z.; Jia, P. An Integrative, Genomic, Transcriptomic and Network-Assisted Study to Identify Genes Associated with Human Cleft Lip with or without Cleft Palate. BMC Med. Genom. 2020, 13 (Suppl. 5), 39. [Google Scholar] [CrossRef]
- Shibano, M.; Watanabe, A.; Takano, N.; Mishima, H.; Kinoshita, A.; Yoshiura, K.-I.; Shibahara, T. Target Capture/next-Generation Sequencing for Nonsyndromic Cleft Lip and Palate in the Japanese Population. Cleft Palate Craniofac. J. 2020, 57, 80–87. [Google Scholar] [CrossRef]
- Dąbrowska, J.; Biedziak, B.; Bogdanowicz, A.; Mostowska, A. Identification of Novel Risk Variants of Non-Syndromic Cleft Palate by Targeted Gene Panel Sequencing. J. Clin. Med. 2023, 12, 2051. [Google Scholar] [CrossRef]
- Smarius, B.; Loozen, C.; Manten, W.; Bekker, M.; Pistorius, L.; Breugem, C. Accurate Diagnosis of Prenatal Cleft Lip/Palate by Understanding the Embryology. World J. Methodol. 2017, 7, 93–100. [Google Scholar] [CrossRef]
- Wang, K.H.; Heike, C.L.; Clarkson, M.D.; Mejino, J.L.V.; Brinkley, J.F.; Tse, R.W.; Birgfeld, C.B.; Fitzsimons, D.A.; Cox, T.C. Evaluation and Integration of Disparate Classification Systems for Clefts of the Lip. Front. Physiol. 2014, 5, 163. [Google Scholar] [CrossRef] [Green Version]
- Konstantonis, D.; Nassika, M.; Athanasiou, M.; Vastardis, H. Subphenotypes in Non-Syndromic Orofacial Cleft Patients Based on the Tooth Agenesis Code (TAC). Children 2022, 9, 437. [Google Scholar] [CrossRef]
- Roth, D.M.; Bayona, F.; Baddam, P.; Graf, D. Craniofacial Development: Neural Crest in Molecular Embryology. Head Neck Pathol. 2021, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Rodriguez, G.; Han, X.; Janečková, E.; Kahng, S.; Song, B.; Chai, Y. Regulatory Mechanisms of Soft Palate Development and Malformations. J. Dent. Res. 2019, 98, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.S.; Goudy, S.L. Cellular and Molecular Mechanisms of Cleft Palate Development: CLP Development. Laryngoscope Investig. Otolaryngol. 2019, 4, 160–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.R.; Chung, H.M.; Matsuk, V.; Fink, D.M.; Stebbins, M.J.; Palecek, S.P.; Shusta, E.V.; Lipinski, R.J. Sonic Hedgehog Signaling in Cranial Neural Crest Cells Regulates Microvascular Morphogenesis in Facial Development. Front. Cell Dev. Biol. 2020, 8, 590539. [Google Scholar] [CrossRef]
- Chong, H.J.; Young, N.M.; Hu, D.; Jeong, J.; McMahon, A.P.; Hallgrimsson, B.; Marcucio, R.S. Signaling by SHH Rescues Facial Defects Following Blockade in the Brain. Dev. Dyn. 2012, 241, 247–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramyan, J. Hedgehog Signaling and Embryonic Craniofacial Disorders. J. Dev. Biol. 2019, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Schock, E.N.; LaBonne, C. Sorting Sox: Diverse Roles for Sox Transcription Factors during Neural Crest and Craniofacial Development. Front. Physiol. 2020, 11, 606889. [Google Scholar] [CrossRef]
- Gordon, R.J.; Levine, M.A. Genetic Disorders of Parathyroid Development and Function. Endocrinol. Metab. Clin. North Am. 2018, 47, 809–823. [Google Scholar] [CrossRef]
- Alatzoglou, K.S.; Kelberman, D.; Dattani, M.T. The Role of SOX Proteins in Normal Pituitary Development. J. Endocrinol. 2009, 200, 245–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alatzoglou, K.S.; Azriyanti, A.; Rogers, N.; Ryan, F.; Curry, N.; Noakes, C.; Bignell, P.; Hall, G.W.; Littooij, A.S.; Saunders, D.; et al. SOX3 Deletion in Mouse and Human Is Associated with Persistence of the Craniopharyngeal Canal. J. Clin. Endocrinol. Metab. 2014, 99, E2702–E2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Gil, N.; Ugartondo, N.; Grinberg, D.; Balcells, S. Wnt Pathway Extracellular Components and Their Essential Roles in Bone Homeostasis. Genes 2022, 13, 138. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Lu, Y.; Liu, X.; Huang, X.; Keller, E.T.; Qian, C.-N.; Zhang, J. Wnt3a: Functions and Implications in Cancer. Chin. J. Cancer 2015, 34, 554–562. [Google Scholar] [CrossRef] [Green Version]
- Marchini, M.; Hu, D.; Lo Vercio, L.; Young, N.M.; Forkert, N.D.; Hallgrímsson, B.; Marcucio, R. Wnt Signaling Drives Correlated Changes in Facial Morphology and Brain Shape. Front. Cell Dev. Biol. 2021, 9, 644099. [Google Scholar] [CrossRef]
- Slavec, L.; Karas Kuželički, N.; Locatelli, I.; Geršak, K. Genetic Markers for Non-Syndromic Orofacial Clefts in Populations of European Ancestry: A Meta-Analysis. Sci. Rep. 2022, 12, 1214. [Google Scholar] [CrossRef]
- Jin, Y.-R.; Han, X.H.; Nishimori, K.; Ben-Avraham, D.; Oh, Y.J.; Shim, J.-W.; Yoon, J.K. Canonical WNT/β-Catenin Signaling Activated by WNT9b and RSPO2 Cooperation Regulates Facial Morphogenesis in Mice. Front. Cell Dev. Biol. 2020, 8, 264. [Google Scholar] [CrossRef]
- Jin, Y.-R.; Han, X.H.; Taketo, M.M.; Yoon, J.K. Wnt9b-Dependent FGF Signaling Is Crucial for Outgrowth of the Nasal and Maxillary Processes during Upper Jaw and Lip Development. Development 2012, 139, 1821–1830. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, K.; Kumari, P.; Sepulveda Rincon, L.; Gu, R.; Ji, Y.; Kumar, S.; Zhou, C.J. Wnt Signaling in Orofacial Clefts: Crosstalk, Pathogenesis and Models. Dis. Model. Mech. 2019, 12, dmm037051. [Google Scholar] [CrossRef] [Green Version]
- Fontoura, C.; Silva, R.M.; Granjeiro, J.M.; Letra, A. Association of WNT9B Gene Polymorphisms with Nonsyndromic Cleft Lip with or without Cleft Palate in Brazilian Nuclear Families. Cleft Palate Craniofac. J. 2015, 52, 44–48. [Google Scholar] [CrossRef] [Green Version]
- Chiquet, B.T.; Blanton, S.H.; Burt, A.; Ma, D.; Stal, S.; Mulliken, J.B.; Hecht, J.T. Variation in WNT Genes Is Associated with Non-Syndromic Cleft Lip with or without Cleft Palate. Hum. Mol. Genet. 2008, 17, 2212–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevanovic, M.; Drakulic, D.; Lazic, A.; Ninkovic, D.S.; Schwirtlich, M.; Mojsin, M. SOX Transcription Factors as Important Regulators of Neuronal and Glial Differentiation during Nervous System Development and Adult Neurogenesis. Front. Mol. Neurosci. 2021, 14, 654031. [Google Scholar] [CrossRef]
- Kormish, J.D.; Sinner, D.; Zorn, A.M. Interactions between SOX Factors and Wnt/Beta-Catenin Signaling in Development and Disease. Dev. Dyn. 2010, 239, 56–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Zevallos, S.E.; Rizzoti, K.; Jeong, Y.; Lovell-Badge, R.; Epstein, D.J. Disruption of SoxB1-Dependent Sonic Hedgehog Expression in the Hypothalamus Causes Septo-Optic Dysplasia. Dev. Cell 2012, 22, 585–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, S.M.; Raine, L.; Fanger, H. The Use of Antiavidin Antibody and Avidin-Biotin-Peroxidase Complex in Immunoperoxidase Technics. Am. J. Clin. Pathol. 1981, 75, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Pilmane, M.; Luts, A.; Sundler, F. Changes in Neuroendocrine Elements in Bronchial Mucosa in Chronic Lung Disease in Adults. Thorax 1995, 50, 551–554. [Google Scholar] [CrossRef] [Green Version]
- Fedchenko, N.; Reifenrath, J. Different Approaches for Interpretation and Reporting of Immunohistochemistry Analysis Results in the Bone Tissue-a Review. Diagn. Pathol. 2014, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- Manola, I.; Mataic, A.; Drvar, D.L.; Pezelj, I.; Dzombeta, T.R.; Kruslin, B. Peritumoral Clefting and Expression of MMP-2 and MMP-9 in Basal Cell Carcinoma of the Skin. In Vivo 2020, 34, 1271–1275. [Google Scholar] [CrossRef]
- Vitenberga, Z.; Pilmane, M.; Babjoniševa, A. The Evaluation of Inflammatory, Anti-Inflammatory and Regulatory Factors Contributing to the Pathogenesis of COPD in Airways. Pathol. Res. Pr. 2019, 215, 97–105. [Google Scholar] [CrossRef]
- Kurosaka, H. The Roles of Hedgehog Signaling in Upper Lip Formation. Biomed Res. Int. 2015, 2015, 901041. [Google Scholar] [CrossRef] [Green Version]
- Everson, J.L.; Fink, D.M.; Yoon, J.W.; Leslie, E.J.; Kietzman, H.W.; Ansen-Wilson, L.J.; Chung, H.M.; Walterhouse, D.O.; Marazita, M.L.; Lipinski, R.J. Sonic Hedgehog Regulation of Foxf2 Promotes Cranial Neural Crest Mesenchyme Proliferation and Is Disrupted in Cleft Lip Morphogenesis. Development 2017, 144, 2082–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, N.L.; Brookes, K.J.; Dixon, M.J. Ectopic Hedgehog Signaling Causes Cleft Palate and Defective Osteogenesis. J. Dent. Res. 2018, 97, 1485–1493. [Google Scholar] [CrossRef]
- Rizzoti, K.; Lovell-Badge, R. SOX3 Activity during Pharyngeal Segmentation Is Required for Craniofacial Morphogenesis. Development 2007, 134, 3437–3448. [Google Scholar] [CrossRef] [Green Version]
- Yao, T.; Yang, L.; Li, P.-Q.; Wu, H.; Xie, H.-B.; Shen, X.; Xie, X.-D. Association of Wnt3A Gene Variants with Non-Syndromic Cleft Lip with or without Cleft Palate in Chinese Population. Arch. Oral Biol. 2011, 56, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Dworkin, S.; Boglev, Y.; Owens, H.; Goldie, S. The Role of Sonic Hedgehog in Craniofacial Patterning, Morphogenesis and Cranial Neural Crest Survival. J. Dev. Biol. 2016, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Dixon, M.J.; Marazita, M.L.; Beaty, T.H.; Murray, J.C. Cleft Lip and Palate: Understanding Genetic and Environmental Influences. Nat. Rev. Genet. 2011, 12, 167–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.; Ryan, R.C.; Zhang, Z.; Bullard, S.A.; Bush, J.O.; Maltby, K.M.; Lidral, A.C.; Jiang, R. Expression of Wnt9b and Activation of Canonical Wnt Signaling during Midfacial Morphogenesis in Mice. Dev. Dyn. 2006, 235, 1448–1454. [Google Scholar] [CrossRef] [Green Version]
- Sethi, J.K.; Vidal-Puig, A. Wnt Signalling and the Control of Cellular Metabolism. Biochem. J. 2010, 427, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, R.; Bush, J.O.; Lidral, A.C. Development of the Upper Lip: Morphogenetic and Molecular Mechanisms. Dev. Dyn. 2006, 235, 1152–1166. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Hao, H.; Reynolds, K.; McMahon, M.; Zhou, C.J. Wnt Signaling in Neural Crest Ontogenesis and Oncogenesis. Cells 2019, 8, 1173. [Google Scholar] [CrossRef] [Green Version]
- Sagai, T.; Amano, T.; Maeno, A.; Ajima, R.; Shiroishi, T. SHH Signaling Mediated by a Prechordal and Brain Enhancer Controls Forebrain Organization. Proc. Natl. Acad. Sci. USA 2019, 116, 23636–23642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Sears, M.J.; Zhang, Z.; Li, H.; Salhab, I.; Krebs, P.; Xing, Y.; Nah, H.-D.; Williams, T.; Carstens, R.P. Cleft Lip and Cleft Palate in Esrp1 Knockout Mice Is Associated with Alterations in Epithelial-Mesenchymal Crosstalk. Development 2020, 147, dev187369. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, E.; Li, B.; Zewdu, R.; Wells, V.; Hebert, J.M.; Karner, C.; Anderson, M.J.; Williams, T.; Dixon, J.; Dixon, M.J.; et al. A Conserved Pbx-Wnt-P63-Irf6 Regulatory Module Controls Face Morphogenesis by Promoting Epithelial Apoptosis. Dev. Cell 2011, 21, 627–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaMantia, A.-S. Why Does the Face Predict the Brain? Neural Crest Induction, Craniofacial Morphogenesis, and Neural Circuit Development. Front. Physiol. 2020, 11, 610970. [Google Scholar] [CrossRef]
- Levi, B.; Brugman, S.; Wong, V.W.; Grova, M.; Longaker, M.T.; Wan, D.C. Palatogenesis: Engineering, Pathways and Pathologies: Engineering, Pathways and Pathologies. Organogenesis 2011, 7, 242–254. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Tanegashima, K.; Ro, H.; Dawid, I.B. Lrig3 Regulates Neural Crest Formation in Xenopus by Modulating Fgf and Wnt Signaling Pathways. Development 2008, 135, 1283–1293. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SHH | SOX3 | WNT3A | WNT9B | |||||
|---|---|---|---|---|---|---|---|---|
| Epithelium | Connective Tissue | Epithelium | Connective Tissue | Epithelium | Connective Tissue | Epithelium | Connective Tissue | |
| Controls | ++/+++ | ++ | +++/++++ | +++ | ++/+++ | +++ | +++/++++ | +++/++++ |
| Unilateral Cleft Lip | ++/+++ | ++/+++ | ++/+++ | ++ | +/++ | + | ++/+++ | +++ |
| Bilateral Cleft Lip | ++ | +/++ | ++ | ++–++/+++ | + | + | ++/+++ | ++/+++ |
| Cleft Palate | +/++ | +/++ | +/++ | ++ | + | +/++ | ++ | ++/+++ |
| H | 43.261 | 40.626 | 34.178 | 12.838 | 19.849 | 16.259 | 26.492 | 20.123 |
| p | <0.001 | <0.001 | <0.001 | 0.005 | <0.001 | 0.001 | <0.001 | <0.001 |
| Strength of Correlation | Correlations between Factors in UCL Group | rs | p |
|---|---|---|---|
| Strong (0.6–0.8) | WNT9B in epithelium and WNT9B in connective tissue | 0.662 | <0.001 |
| Moderate (0.4–0.6) | SOX3 in epithelium and SOX3 in connective tissue | 0.599 | <0.001 |
| SHH in epithelium and WNT9B in epithelium | 0.578 | <0.001 | |
| SOX3 in epithelium and WNT9B in epithelium | 0.524 | 0.001 | |
| WNT3A in epithelium and WNT9B in epithelium | 0.521 | 0.001 | |
| SHH in epithelium and WNT3A epithelium | 0.510 | 0.001 | |
| SOX3 in epithelium and WNT3A in epithelium | 0.496 | 0.002 | |
| SOX3 in epithelium and WNT9B in connective tissue | 0.445 | 0.007 | |
| SHH in connective tissue and WNT3A in epithelium | 0.444 | 0.007 | |
| SHH in connective tissue and SOX3 in connective tissue | 0.432 | 0.008 | |
| SHH in epithelium and SOX3 in epithelium | 0.415 | 0.012 | |
| Weak (0.2–0.4) | SOX3 in connective tissue and WNT9B in connective tissue | 0.392 | 0.018 |
| SHH in connective tissue and WNT3A in connective tissue | 0.384 | 0.021 | |
| SOX3 in connective tissue and WNT3A in epithelium | 0.375 | 0.024 | |
| SHH in epithelium and WNT9B in connective tissue | 0.374 | 0.025 | |
| SHH in connective tissue and SOX3 in epithelium | 0.341 | 0.042 |
| Strength of Correlation | Correlations between Factors in BCL Group | rs | p |
|---|---|---|---|
| Very strong (0.8–1.0) | SHH in epithelium and WNT9B in epithelium | 0.812 | 0.001 |
| Strong (0.6–0.8) | SOX3 in epithelium and WNT9B in epithelium | 0.713 | 0.009 |
| WNT3A in epithelium and WNT9B in connective tissue | 0.687 | 0.014 | |
| WNT3A in epithelium and WNT9B in epithelium | 0.659 | 0.020 | |
| SHH in connective tissue and SOX3 in epithelium | 0.617 | 0.032 | |
| Moderate (0.4–0.6) | SHH in connective tissue and WNT3A in epithelium | 0.591 | 0.043 |
| SHH in epithelium and SOX3 in epithelium | 0.580 | 0.048 |
| Strength of Correlation | Correlations between Factors in CP Group | rs | p |
|---|---|---|---|
| Positive correlations | |||
| Very strong (0.8–1.0) | SOX3 in epithelium and WNT9B in epithelium | 0.832 | <0.001 |
| Moderate (0.4–0.6) | SHH in connective tissue and SOX3 in connective tissue | 0.440 | 0.025 |
| Weak (0.2–0.4) | SOX3 in connective tissue and WNT9B in connective tissue | 0.397 | 0.050 |
| Negative correlations | |||
| Moderate (−0.6…−0.4) | SOX3 in connective tissue and WNT3A in epithelium | −0.458 | 0.028 |
| SHH in epithelium and SOX3 in connective tissue | −0.417 | 0.043 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaivads, M.; Akota, I.; Pilmane, M. Characterization of SHH, SOX3, WNT3A and WNT9B Proteins in Human Non-Syndromic Cleft Lip and Palate Tissue. Dent. J. 2023, 11, 151. https://doi.org/10.3390/dj11060151
Vaivads M, Akota I, Pilmane M. Characterization of SHH, SOX3, WNT3A and WNT9B Proteins in Human Non-Syndromic Cleft Lip and Palate Tissue. Dentistry Journal. 2023; 11(6):151. https://doi.org/10.3390/dj11060151
Chicago/Turabian StyleVaivads, Mārtiņš, Ilze Akota, and Māra Pilmane. 2023. "Characterization of SHH, SOX3, WNT3A and WNT9B Proteins in Human Non-Syndromic Cleft Lip and Palate Tissue" Dentistry Journal 11, no. 6: 151. https://doi.org/10.3390/dj11060151
APA StyleVaivads, M., Akota, I., & Pilmane, M. (2023). Characterization of SHH, SOX3, WNT3A and WNT9B Proteins in Human Non-Syndromic Cleft Lip and Palate Tissue. Dentistry Journal, 11(6), 151. https://doi.org/10.3390/dj11060151