
Products Released from Structurally Different Dextrans by Bacterial and Fungal Dextranases


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General
2.2. Molecular Cloning and Heterologous Expression
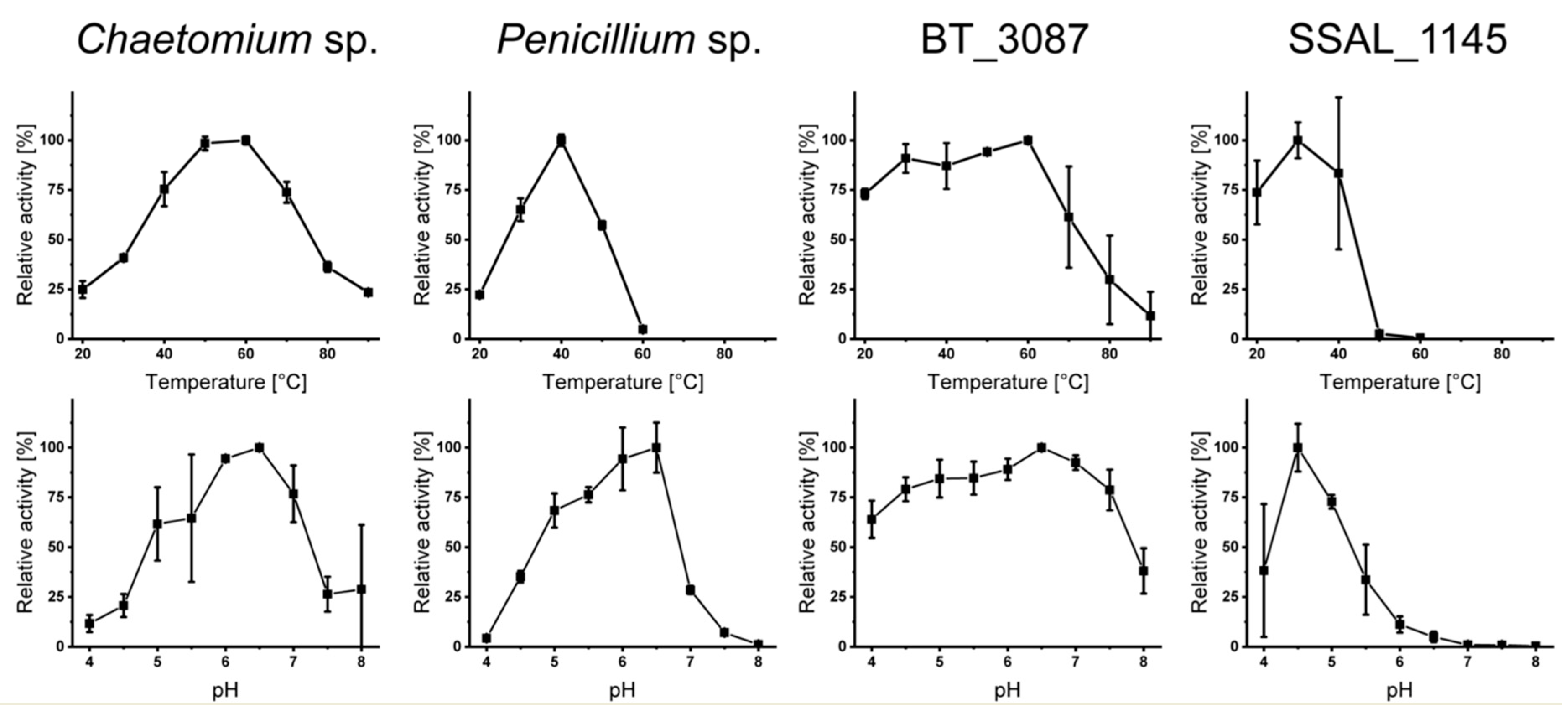
2.3. Optimum Incubation Conditions
2.4. Dextran Production
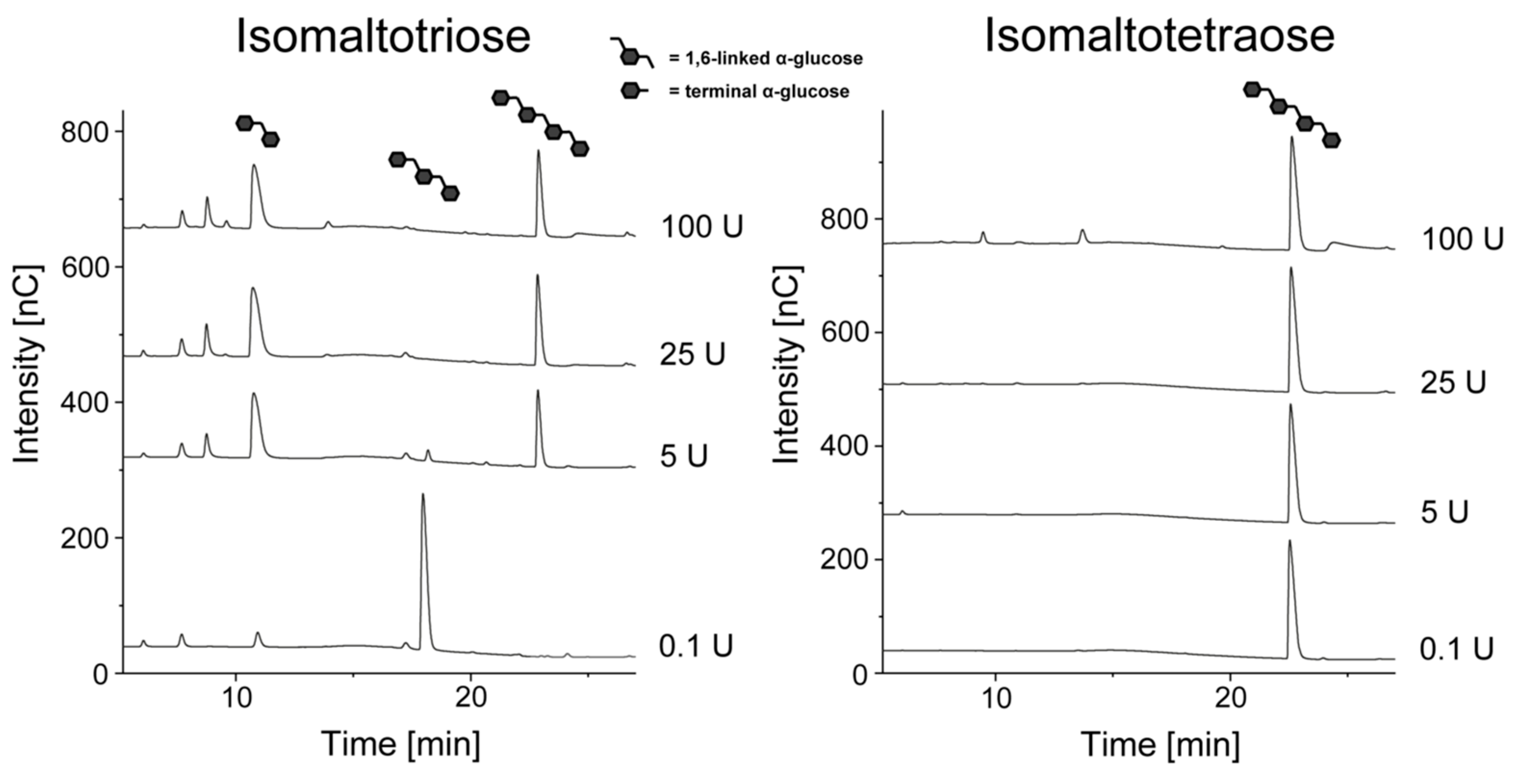
2.5. Dextranase-Mediated Hydrolysis
2.6. High Performance Anion Exchange Chromatography
3. Results and Discussion
3.1. Dextranase Production and Characterization
3.2. Hydrolysis of Linear Dextrans
3.3. Hydrolysis of O3- and O4-Branched Dextrans
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Naessens, M.; Cerdobbel, A.; Soetaert, W.; Vandamme, E. Leuconostoc dextransucrase and dextran: Production, properties and applications. J. Chem. Technol. Biotechnol. 2005, 80, 845–860. [Google Scholar] [CrossRef]
- Leemhuis, H.; Pijning, T.; Dobruchowska, J.M.; Van Leeuwen, S.S.; Kralj, S.; Dijkstra, B.W.; Dijkhuizen, L. Glucansucrases: Three-dimensional structures, reactions, mechanism, α-glucan analysis and their implications in biotechnology and food applications. J. Biotechnol. 2013, 163, 250–272. [Google Scholar] [CrossRef] [Green Version]
- Torino, M.I.; Font, G.; Mozzi, F. Biopolymers from lactic acid bacteria. Novel applications in foods and beverages. Front. Microbiol. 2015, 6, 834. [Google Scholar] [CrossRef] [Green Version]
- Zannini, E.; Waters, D.M.; Coffey, A.; Arendt, E.K. Production, properties, and industrial food application of lactic acid bacteria-derived exopolysaccharides. Appl. Microbiol. Biotechnol. 2016, 100, 1121–1135. [Google Scholar] [CrossRef]
- Khalikova, E.; Susi, P.; Korpela, T. Microbial Dextran-Hydrolyzing Enzymes: Fundamentals and Applications. Microbiol. Mol. Biol. Rev. 2005, 69, 306–325. [Google Scholar] [CrossRef] [Green Version]
- Katina, K.; Maina, N.H.; Juvonen, R.; Flander, L.; Johansson, L.; Virkki, L.; Tenkanen, M.; Laitila, A. In situ production and analysis of Weissella confusa dextran in wheat sourdough. Food Microbiol. 2009, 26, 734–743. [Google Scholar] [CrossRef]
- Maina, N.H.; Virkki, L.; Pynnönen, H.; Maaheimo, H.; Tenkanen, M. Structural Analysis of Enzyme-Resistant Isomaltooligosaccharides Reveals the Elongation of α-(1→3)-Linked Branches in Weissella confusa Dextran. Biomacromolecules 2011, 12, 409–418. [Google Scholar] [CrossRef]
- Fels, L.; Jakob, F.; Vogel, R.F.; Wefers, D. Structural characterization of the exopolysaccharides from water kefir. Carbohydr. Polym. 2018, 189, 296–303. [Google Scholar] [CrossRef]
- Xu, D.; Fels, L.; Wefers, D.; Behr, J.; Jakob, F.; Vogel, R.F. Lactobacillus hordei dextrans induce Saccharomyces cerevisiae aggregation and network formation on hydrophilic surfaces. Int. J. Biol. Macromol. 2018, 115, 236–242. [Google Scholar] [CrossRef]
- Bechtner, J.; Wefers, D.; Schmid, J.; Vogel, R.F.; Jakob, F. Identification and comparison of two closely related dextransucrases released by water kefir borne Lactobacillus hordei TMW 1.1822 and Lactobacillus nagelii TMW 1.1827. Microbiology 2019, 165, 956–966. [Google Scholar] [CrossRef]
- Münkel, F.; Wefers, D. Fine structures of different dextrans assessed by isolation and characterization of endo-dextranase liberated isomalto-oligosaccharides. Carbohydr. Polym. 2019, 215, 296–306. [Google Scholar] [CrossRef]
- Münkel, F.; Bechtner, J.; Eckel, V.; Fischer, A.; Herbi, F.; Jakob, F.; Wefers, D. Detailed Structural Characterization of Glucans Produced by Glucansucrases from Leuconostoc citreum TMW 2.1194. J. Agric. Food Chem. 2019, 67, 6856–6866. [Google Scholar] [CrossRef]
- Münkel, F.; Fischer, A.; Wefers, D. Structural characterization of mixed-linkage α-glucans produced by mutants of Lactobacillus reuteri TMW 1.106 dextransucrase. Carbohydr. Polym. 2020, 231, 115697. [Google Scholar] [CrossRef]
- Das, D.K.; Dutta, S.K. Purification, biochemical characterisation and mode of action of an extracellular endo-dextranase from the culture filtrate of Penicillium lilacinum. Int. J. Biochem. Cell Biol. 1996, 28, 107–113. [Google Scholar] [CrossRef]
- Hattori, A.; Ishibashi, K.; Minato, S. The purification and characterization of the dextranase of Chaetomium gracile. Agric. Biol. Chem. 1981, 45, 2409–2416. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A.M.; Andersson, R.; Stahlberg, J.; Kenne, L.; Jones, T.A. Dextranase from Penicillum minioluteum: Reaction course, crystal structure, and product complex. Structure 2003, 11, 1111–1121. [Google Scholar] [CrossRef]
- Sugiura, M.; Ito, A.; Ogiso, T.; Kato, K.; Asano, H. Studies on dextranase—Purification of dextranase from Penicillium funiculosum and its enzymatic properties. Biochim. Biophys. Acta 1973, 309, 357–362. [Google Scholar] [CrossRef]
- Virgen-Ortíz, J.; Ibarra-Junquera, V.; Escalanteminakata, P.; Ornelas-Paz, J.D.J.; Osunacastro, J.A.; González-Potes, A. Kinetics and thermodynamic of the purified dextranase from Chaetomium erraticum. J. Mol. Catal. B Enzym. 2015, 122, 80–86. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, N.; Tian, Y. Purification, characterization, and biocatalytic potential of a novel dextranase from Chaetomium globosum. Biotechnol. Lett. 2018, 40, 1407–1418. [Google Scholar] [CrossRef]
- Yilan, O.; Sun, X.; Du, K.; Ouyang, Y.; Wu, C.; Xu, N.; Linhardt, R.J.; Zhang, Z. UP-HILIC-MS/MS to Determine the Action Pattern of Penicillium sp. Dextranase. J. Am. Soc. Mass Spectrom. 2015, 26, 1174–1185. [Google Scholar] [CrossRef]
- Erhardt, F.A.; Stammen, S.; Jördening, H.-J. Production, characterization and (co-)immobilization of dextranase from Penicillium aculeatum. Biotechnol. Lett. 2008, 30, 1069–1073. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.J.; Dewar, M.D. The action pattern of Penicillium lilacinum dextranase. Carbohydr. Res. 1975, 39, 303–315. [Google Scholar] [CrossRef]
- Hiraoka, N.; Tsuji, H.; Fukumoto, J.; Yamamoto, T.; Tsuru, D. Studies on Mold Dextranases: Some Physicochemical Properties and Substrate Specificity of Dextranases Obtained from Aspergillus carneus and Penicillium luteum. Int. J. Pept. Protein Res. 2009, 5, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, M.; Prabhu, K.A. Studies on dextranase from Penicillium aculeatum. Enzyme Microb. Technol. 1984, 6, 217–220. [Google Scholar]
- Kim, Y.-M.; Yamamoto, E.; Kang, M.-S.; Nakai, H.; Saburi, W.; Okuyama, M.; Mori, H.; Funane, K.; Momma, M.; Fujimoto, Z.; et al. Bacteroides thetaiotaomicron VPI-5482 glycoside hydrolase family 66 homolog catalyzes dextranolytic and cyclization reactions. FEBS J. 2012, 279, 3185–3191. [Google Scholar] [CrossRef]
- Igarashi, T.; Morisaki, H.; Goto, N. Molecular Characterization of Dextranase from Streptococcus rattus. Microbiol. Immunol. 2004, 48, 155–162. [Google Scholar] [CrossRef]
- Wanda, S.Y.; Curtiss, R. Purification and characterization of Streptococcus sobrinus dextranase produced in recombinant Escherichia coli and sequence analysis of the dextranase gene. J. Bacteriol. 1994, 176, 3839–3850. [Google Scholar] [CrossRef] [Green Version]
- Pulkownik, A.; Walker, G.J. Purification and substrate specificity of an endo-dextranase of Streptococcus mutans K1-R. Carbohydr. Res. 1977, 54, 237–251. [Google Scholar] [CrossRef]
- Van Bueren, A.L.; Saraf, A.; Martens, E.C.; Dijkhuizen, L. Differential Metabolism of Exopolysaccharides from Probiotic Lactobacilli by the Human Gut Symbiont Bacteroides thetaiotaomicron. Appl. Environ. Microbiol. 2015, 81, 3973–3983. [Google Scholar] [CrossRef] [Green Version]
- Rühmkorf, C.; Rübsam, H.; Becker, T.; Bork, C.; Voiges, K.; Mischnick, P.; Brandt, M.J.; Vogel, R.F. Effect of structurally different microbial homoexopolysaccharides on the quality of gluten-free bread. Eur. Food Res. Technol. 2012, 235, 139–146. [Google Scholar] [CrossRef]
- Aslanidis, C.; De Jong, P.J. Ligation-independent cloning of PCR products (LIC-PCR). Nucleic Acids Res. 1990, 18, 6069–6074. [Google Scholar] [CrossRef] [PubMed]
- Burgess-Brown, N.A.; Sharma, S.; Sobott, F.; Loenarz, C.; Oppermann, U.; Gileadi, O. Codon optimization can improve expression of human genes in Escherichia coli: A multi-gene study. Protein Expr. Purif. 2008, 59, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Stols, L.; Gu, M.Y.; Dieckman, L.; Raffen, R.; Collart, F.R.; Donnelly, M.I. A new vector for high-throughput, ligation-independent cloning encoding a tobacco etch virus protease cleavage site. Protein Expr. Purif. 2002, 25, 8–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rühmkorf, C.; Bork, C.; Mischnick, P.; Rübsam, H.; Becker, T.; Vogel, R.F. Identification of Lactobacillus curvatus TMW 1.624 dextransucrase and comparative characterization with Lactobacillus reuteri TMW 1.106 and Lactobacillus animalis TMW 1.971 dextransucrases. Food Microbiol. 2013, 34, 52–61. [Google Scholar] [CrossRef]
- Sawai, T.; Niwa, Y. Transisomaltosylation activity of a bacterial isomalto-dextranase. Agric. Biol. Chem. 1975, 39, 1077–1083. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pittrof, S.L.; Kaufhold, L.; Fischer, A.; Wefers, D. Products Released from Structurally Different Dextrans by Bacterial and Fungal Dextranases. Foods 2021, 10, 244. https://doi.org/10.3390/foods10020244
Pittrof SL, Kaufhold L, Fischer A, Wefers D. Products Released from Structurally Different Dextrans by Bacterial and Fungal Dextranases. Foods. 2021; 10(2):244. https://doi.org/10.3390/foods10020244
Chicago/Turabian StylePittrof, Silke L., Larissa Kaufhold, Anja Fischer, and Daniel Wefers. 2021. "Products Released from Structurally Different Dextrans by Bacterial and Fungal Dextranases" Foods 10, no. 2: 244. https://doi.org/10.3390/foods10020244