Effect of Unsaturated Fatty Acids on Intra-Metabolites and Aroma Compounds of Saccharomyces cerevisiae in Wine Fermentation



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Fermentation Medium and Yeast Strains
2.3. Fermentation Conditions
2.4. Analysis Methods
2.4.1. Major Fermentation Products Analyses
2.4.2. Fatty Acids Analyses
2.4.3. Volatile Compounds Analyses
2.5. Metabolome Analysis
2.6. Real-Time qPCR Assay
2.7. Statistical Analysis
3. Results
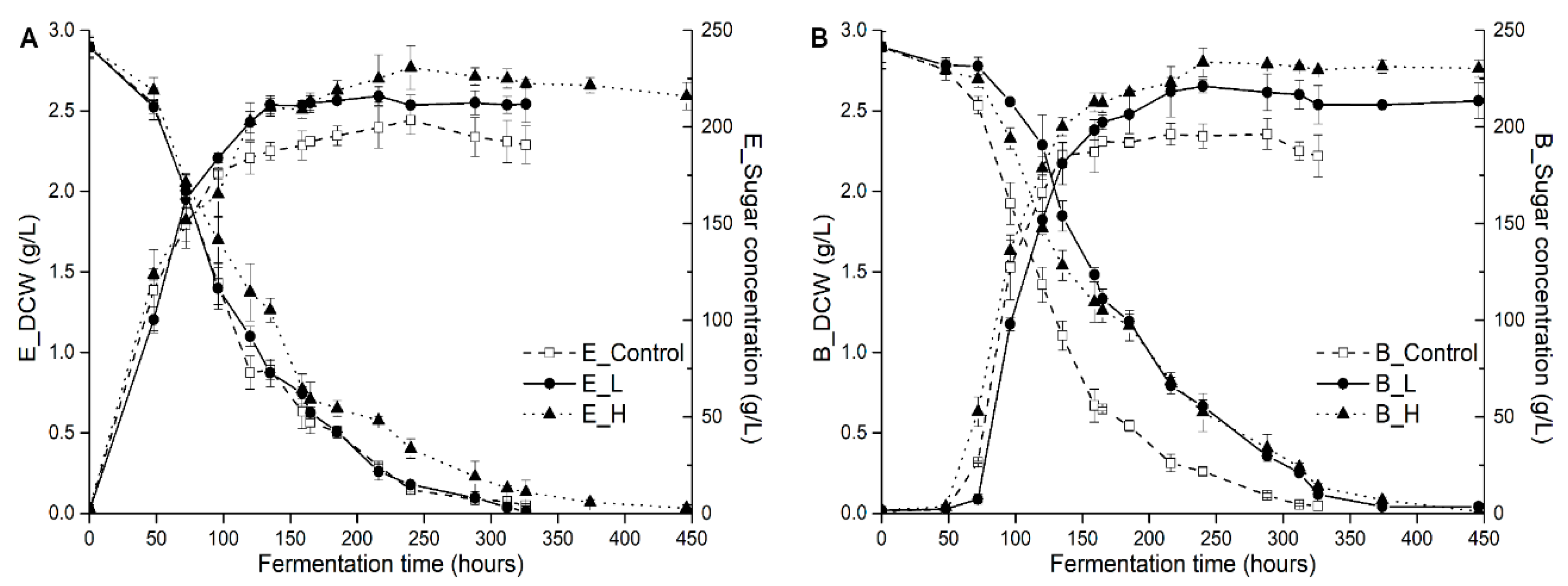
3.1. Effects of Pre-Fermentative Addition of UFAs Mixture on Yeast Growths, Fermentation Activity and Major Fermentation Products
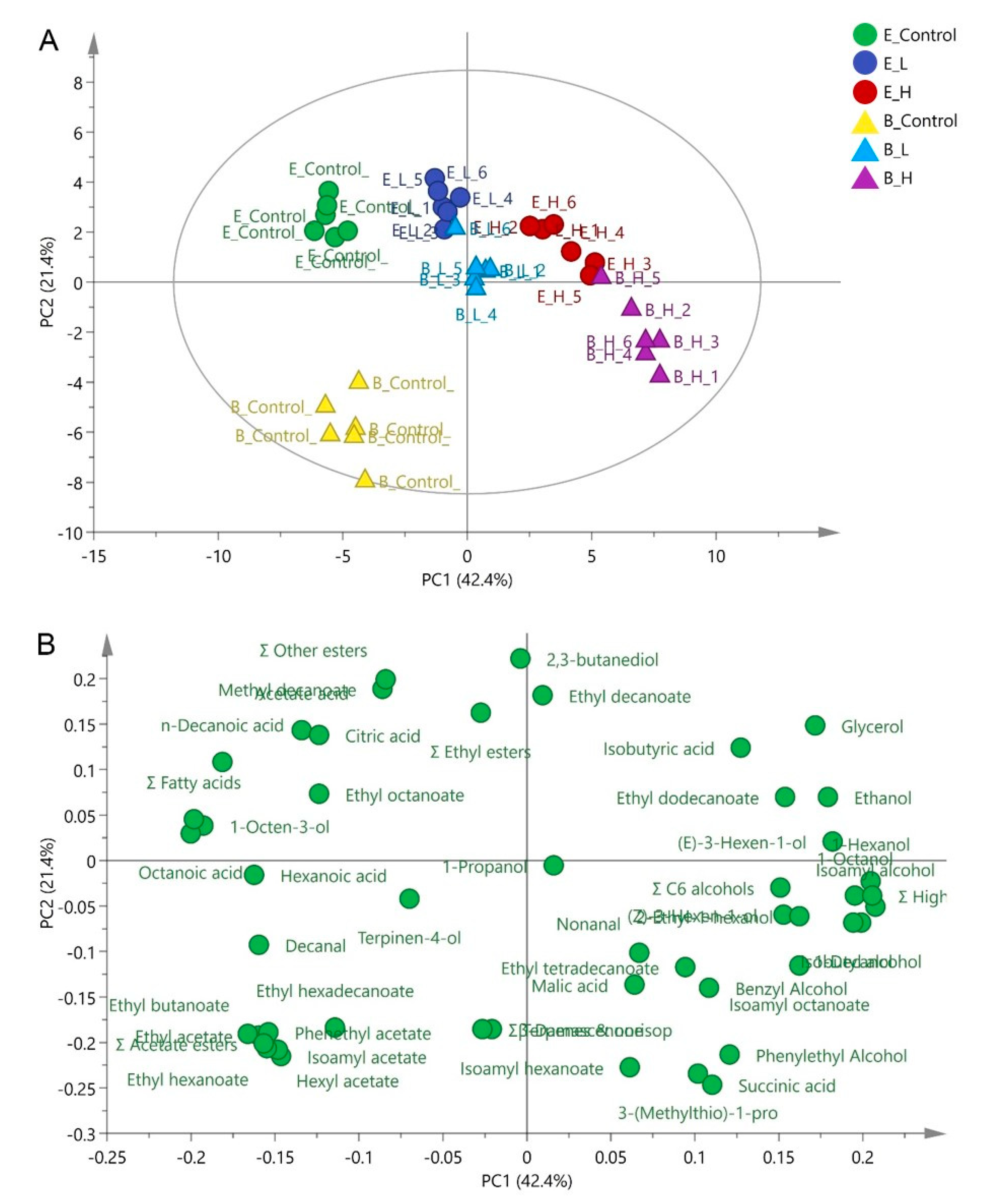
3.2. Effects of Pre-Fermentative Addition of UFAs Mixture on Volatile Compounds Production
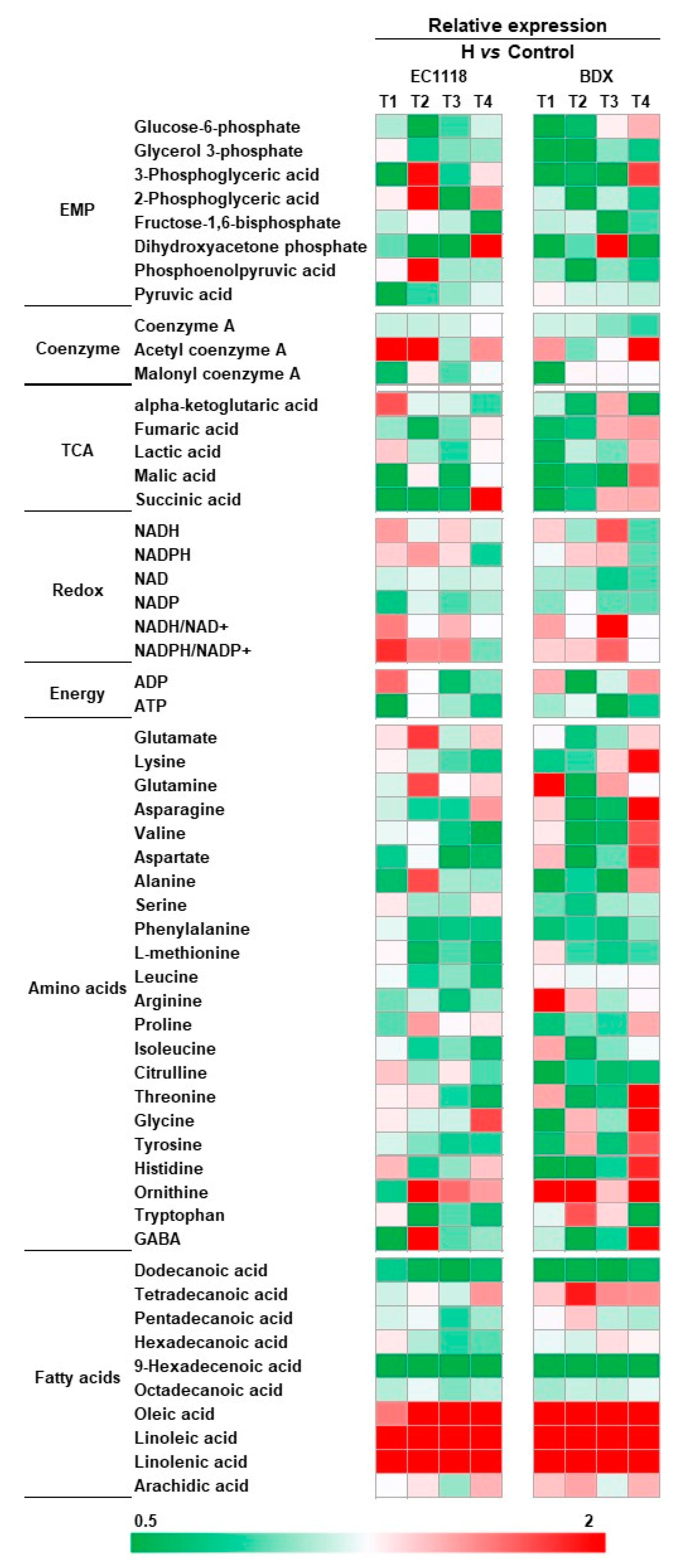
3.3. Effects of Pre-Fermentative Addition of UFAs Mixture on Intra-Metabolite Profile
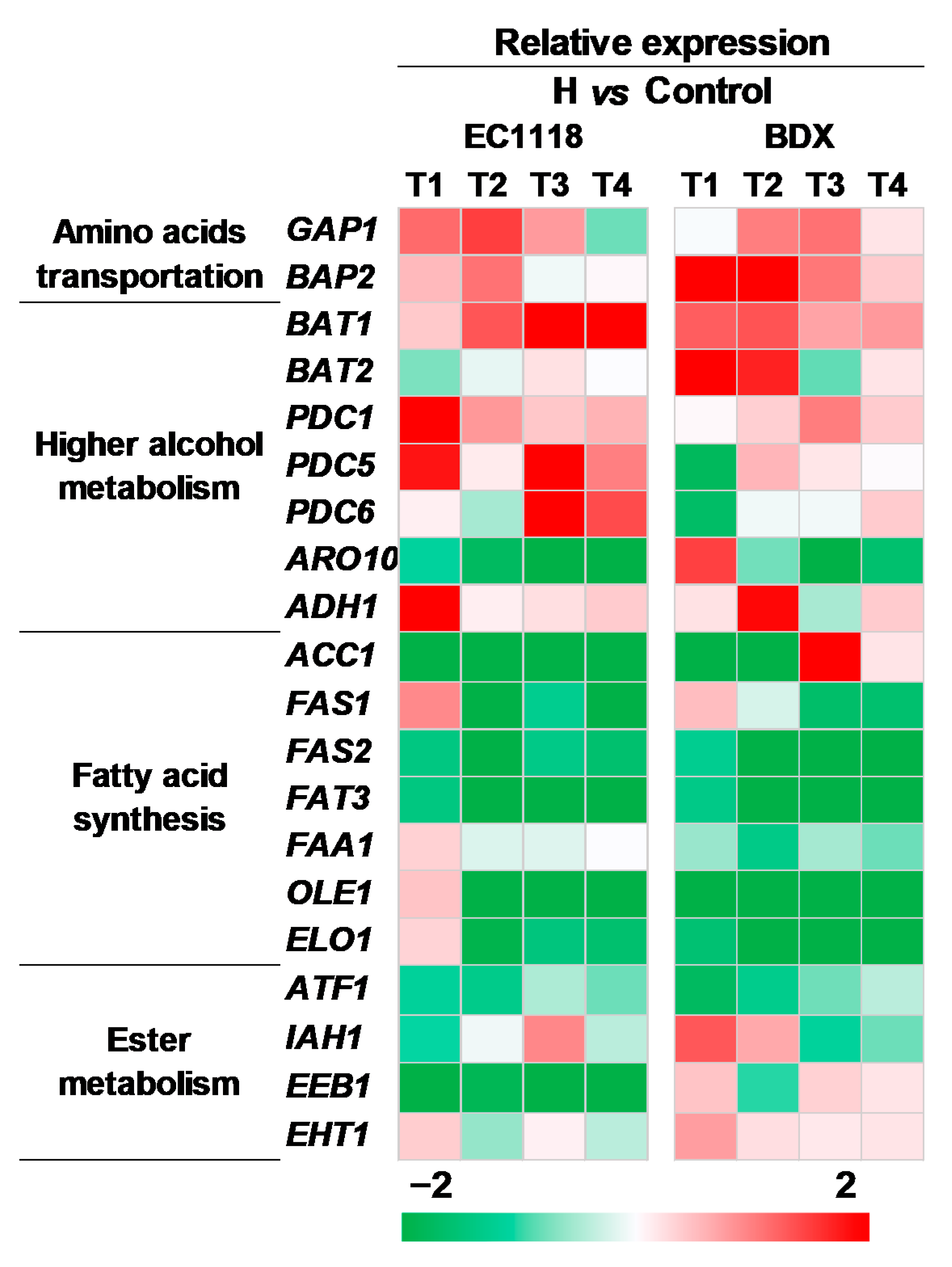
3.4. Effects of Pre-Fermentative Addition of UFAs Mixture Genes Expression Related to Aroma Compounds Formation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Antalick, G.; Suklje, K.; Blackman, J.W.; Meeks, C.; Deloire, A.; Schmidtke, L.M. Influence of grape composition on red wine ester profile: Comparison between Cabernet Sauvignon and Shiraz cultivars from Australian warm climate. J. Agric. Food Chem. 2015, 63, 4664–4672. [Google Scholar] [CrossRef] [PubMed]
- Mato, I.; Suárez-Luque, S.; Huidobro, J.F. Simple determination of main organic acids in grape juice and wine by using capillary zone electrophoresis with direct UV detection. Food Chem. 2007, 102, 104–112. [Google Scholar] [CrossRef]
- Deed, N.K.; van Vuuren, H.J.J.; Gardner, R.C. Effects of nitrogen catabolite repression and di-ammonium phosphate addition during wine fermentation by a commercial strain of S. cerevisiae. Appl. Microbiol. Biotechnol. 2011, 89, 1537–1549. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.M.; van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [Green Version]
- Calderbank, J.; Keenan, M.H.; Rose, A.H. Plasma-membrane phospholipid unsaturation affects expression of the general amino-acid permease in Saccharomyces cerevisiae Y185. J. Gen. Microbiol. 1985, 131, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.T.; Yu, K.J.; Li, Y.T.; Duan, C.Q.; Yan, G.L. The content of linoleic acid in grape must influences the aromatic effect of branched-chain amino acids addition on red wine. Food Res. Int. 2018, 114, 214–222. [Google Scholar] [CrossRef]
- Liu, P.T.; Zhang, B.Q.; Duan, C.Q.; Yan, G.L. Pre-fermentative supplementation of unsaturated fatty acids alters the effect of overexpressing ATF1 and EEB1 on esters biosynthesis in red wine. Lwt 2020, 120, 108925. [Google Scholar] [CrossRef]
- Rollero, S.; Mouret, J.-R.; Sanchez, I.; Camarasa, C.; Ortiz-Julien, A.; Sablayrolles, J.-M.; Dequin, S. Key role of lipid management in nitrogen and aroma metabolism in an evolved wine yeast strain. Microb. Cell Fact. 2016, 15, 32. [Google Scholar] [CrossRef]
- Pinu, F.R.; Edwards, P.J.B.; Gardner, R.C.; Villas-Boas, S.G. Nitrogen and carbon assimilation by Saccharomyces cerevisiae during Sauvignon blanc juice fermentation. FEMS Yeast Res. 2014, 14, 1206–1222. [Google Scholar] [CrossRef] [Green Version]
- Varela, C.; Torrea, D.; Schmidt, S.A.; Ancin-Azpilicueta, C.; Henschke, P.A. Effect of oxygen and lipid supplementation on the volatile composition of chemically defined medium and Chardonnay wine fermented with Saccharomyces cerevisiae. Food Chem. 2012, 135, 2863–2871. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.L.; Shi, Y.; Jiang, R.; Yang, Q.; Wang, Y.Q.; Liu, P.T.; Duan, C.Q.; Yan, G.L. Effects of adding unsaturated fatty acids on fatty acid composition of Saccharomyces cerevisiae and major volatile compounds in wine. S. Afr. J. Enol. Vitic. 2015, 36, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Rossouw, D.; Naes, T.; Bauer, F.F. Linking gene regulation and the exo-metabolome: A comparative transcriptomics approach to identify genes that impact on the production of volatile aroma compounds in yeast. BMC Genom. 2008, 9, 530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinu, F.R.; Villas-Boas, S.G.; Martin, D. Pre-fermentative supplementation of fatty acids alters the metabolic activity of wine yeasts. Food Res. Int. 2019, 121, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Ancín, C.; Ayestaran, B.; Garcia, A.; Garrido, J. Influence of vacuum filtration of Viura must on the concentration of fatty acids and their utilization in fermentation. Food Res. Int. 1997, 29, 763–770. [Google Scholar] [CrossRef]
- Varela, F.; Calderón, F.; González, M.C.; Colomo, B.; Suárez, J.A. Effect of clarification on the fatty acid composition of grape must and the fermentation kinetics of white wines. Eur. Food Res. Technol. 1999, 209, 439–444. [Google Scholar] [CrossRef]
- Tumanov, S.; Pinu, F.R.; Greenwood, D.R.; Villas-Bôas, S.G. Effect of free fatty acids and lipolysis on Sauvignon Blanc fermentation. Aust. J. Grape Wine Res. 2018, 24, 398–405. [Google Scholar] [CrossRef]
- GB, T. 2006 Analytical Methods of Wine and Fruit Wine; China Standard Press: Beijing, China, 2006. [Google Scholar]
- Lan, Y.B.; Qian, X.; Yang, Z.J.; Xiang, X.F.; Yang, W.X.; Liu, T.; Zhu, B.Q.; Pan, Q.H.; Duan, C.Q. Striking changes in volatile profiles at sub-zero temperatures during over-ripening of ‘Beibinghong’ grapes in Northeastern china. Food Chem. 2016, 212, 172–182. [Google Scholar] [CrossRef]
- Shao, S.; Zhou, T.; McGarvey, B.D. Comparative metabolomic analysis of Saccharomyces cerevisiae during the degradation of patulin using gas chromatography—mass spectrometry. Appl. Microbiol. Biotechnol. 2012, 94, 789–797. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real- time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Krivoruchko, A.; Zhang, Y.; Siewers, V.; Chen, Y.; Nielsen, J. Microbial acetyl-CoA metabolism and metabolic engineering. Metab. Eng. 2015, 28, 28–42. [Google Scholar] [CrossRef]
- Redón, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effect of lipid supplementation upon Saccharomyces cerevisiae lipid composition and fermentation performance at low temperature. Eur. Food Res. Technol. 2009, 228, 833–840. [Google Scholar] [CrossRef]
- Varela, C.; Pizarro, F.; Agosin, E. Biomass content governs fermentation rate in nitrogen-deficient wine musts. Appl. Environ. Microbiol. 2004, 70, 3392–3400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casu, F.; Pinu, F.R.; Stefanello, E.; Greenwood, D.R.; Villas-Bôas, S.G. The fate of linoleic acid on Saccharomyces cerevisiae metabolism under aerobic and anaerobic conditions. Metabolomics 2018, 14, 103. [Google Scholar] [CrossRef] [PubMed]
- Hettema, E.H.; van Roermund, C.W.; Distel, B.; van den Berg, M.; Vilela, C.; Rodrigues-Pousada, C.; Wanders, R.J.; Tabak, H.F. The ABC transporter proteins Pat1 and Pat2 are required for import of long-chain fatty acids into peroxisomes of Saccharomyces cerevisiae. EMBO J. 1996, 15, 3813–3822. [Google Scholar] [CrossRef] [PubMed]
- Snoek, I.S.I.; Steensma, H.Y. Factors involved in anaerobic growth of Saccharomyces cerevisiae. Yeast 2007, 24, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scanes, K.T.; Hohmann, S.; Prior, B.A. Glycerol production by the yeast Saccharomyces cerevisiae and its relevance to wine: A review. S. Afr. J. Enol. Vitic. 1998, 19, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Saint-Prix, F.; Bönquist, L.; Dequin, S. Functional analysis of the ALD gene family of Saccharomyces cerevisiae during anaerobic growth on glucose: The NADP+-dependent Ald6p and Ald5p isoforms play a major role in acetate formation. Microbiology 2004, 150, 2209–2220. [Google Scholar] [CrossRef]
- Eglinton, J.M.; Heinrich, A.J.; Pollnitz, A.P.; Langridge, P.; Henschke, P.A.; Lopes, M.D.B. Decreasing acetic acid accumulation by a glycerol overproducing strain of Saccharomyces cerevisiae by deleting the ALD6 aldehyde dehydrogenase gene. Yeast 2002, 19, 295–301. [Google Scholar] [CrossRef]
- McDonough, V.M.; Stukey, J.E.; Martin, C.E. Specificity of unsaturated fatty acid-regulated expression of the Saccharomyces cerevisiae OLE1 gene. J. Biol. Chem. 1992, 267, 5931–5936. [Google Scholar] [CrossRef]
- Rossouw, D.; Van Den Dool, A.H.; Jacobson, D.; Bauer, F.F. Comparative transcriptomic and proteomic profiling of industrial wine yeast strains. Appl. Environ. Microbiol. 2010, 76, 3911–3923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, T.; Perez, M.; Mouret, J.R.; Sanchez, I.; Sablayrolles, J.M.; Camarasa, C. Metabolic responses of Saccharomyces cerevisiae to valine and ammonium pulses during four-stage continuous wine fermentations. Appl. Environ. Microbiol. 2013, 79, 2749–2758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoondermark-Stolk, S.A.; Tabernero, M.; Chapman, J.; Ter Schure, E.G.; Verrips, C.T.; Verkleij, A.J.; Boonstra, J. Bat2p is essential in Saccharomyces cerevisiae for fusel alcohol production on the non-fermentable carbon source ethanol. FEMS Yeast Res. 2005, 5, 757–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casu, F.; Pinu, F.R.; Fedrizzi, B.; Greenwood, D.R.; Villas-Boas, S.G. The effect of linoleic acid on the Sauvignon blanc fermentation by different wine yeast strains. FEMS Yeast Res. 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilly, M.; Bauer, F.F.; Lambrechts, M.G.; Swiegers, J.H.; Cozzolino, D.; Pretorius, I.S. The effect of increased yeast alcohol acetyltransferase and esterase activity on the flavour profiles of wine and distillates. Yeast 2006, 23, 641–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saerens, S.M.G.; Verstrepen, K.J.; Van Laere, S.D.M.; Voet, A.R.D.; Van Dijck, P.; Delvaux, F.R.; Thevelein, J.M. The Saccharomyces cerevisiae EHT1 and EEB1 genes encode novel enzymes with medium-chain fatty acid ethyl ester synthesis and hydrolysis capacity. J. Biol. Chem. 2006, 281, 4446–4456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Liu, L.P.; Yang, M.; Ding, X.T.; Jia, S.R.; Dong, J.J.; Zhong, C. Enhancing medium-chain fatty acid ethyl ester production during beer fermentation through EEB1 and ETR1 overexpression in Saccharomyces pastorianus. J. Agric. Food Chem. 2019, 67, 5607–5613. [Google Scholar] [CrossRef]
- Vilanova, M.; Siebert, T.E.; Varela, C.; Pretorius, I.S.; Henschke, P.A. Effect of ammonium nitrogen supplementation of grape juice on wine volatiles and non-volatiles composition of the aromatic grape variety Albariño. Food Chem. 2012, 133, 124–131. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | E_Control | E_L | E_H | B_Control | B_L | B_H | p Value |
|---|---|---|---|---|---|---|---|
| Fermentation duration time (h) | 326 | 326 | 446 | 326 | 446 | 446 | |
| Maximum biomass (g DCW/L) | 2.44 ± 0.09 a | 2.59 ± 0.06 a | 2.77 ± 0.14 b | 2.36 ± 0.09 a | 2.65 ± 0.04 b | 2.80 ± 0.09 b | * |
| Maximum growth rate (g DCW/L/h) | 0.028 ± 0.001 a | 0.028 ± 0.001 a | 0.030 ± 0.002 a | 0.035 ± 0.001 b | 0.036 ± 0.001 b | 0.033 ± 0.002 ab | * |
| Maximum fermentation rate (g/L/h) | 1.94 ± 0.05 a | 1.96 ± 0.03 a | 1.61 ± 0.03 b | 1.93 ± 0.10 a | 1.86 ± 0.06 a | 1.61 ± 0.05 b | * |
| Glycerol (g/L) | 9.32 ± 0.3 a | 10.23 ± 0.2 b | 10.57 ± 0.18 c | 8.78 ± 0.27 a | 10.26 ± 0.06 b | 10.45 ± 0.21 c | * |
| Ethanol (% v/v) | 13.94 ± 0.25 a | 14.38 ± 0.05 b | 14.57 ± 0.05 b | 13.86 ± 0.46 a | 14.49 ± 0.08 b | 14.72 ± 0.19 b | * |
| Citric acid (g/L) | 0.22 ± 0.01 ab | 0.21 ± 0.01 a | 0.21 ± 0.01 ab | 0.21 ± 0.01 a | 0.21 ± 0.02 a | 0.21 ± 0.01 a | * |
| Malic acid (g/L) | 3.10 ± 0.06 a | 3.28 ± 0.22 a | 3.52 ± 0.03 b | 3.58 ± 0.22 b | 3.14 ± 0.07 a | 3.39 ± 0.29 a | * |
| Succinic acid (g/L) | 3.60 ± 0.04 a | 3.79 ± 0.22 ab | 3.87 ± 0.01 ab | 4.39 ± 0.23 c | 3.93 ± 0.05 b | 4.48 ± 0.20 c | * |
| Acetic acid (g/L) | 0.51 ± 0.07 a | 0.42 ± 0.02 b | 0.43 ± 0.01 b | 0.39 ± 0.03 b | 0.31 ± 0.01 c | 0.24 ± 0.04 d | * |
| Compounds | E_Control | E_L | E_H | B_Control | B_L | B_H | p Value | Thresholds |
|---|---|---|---|---|---|---|---|---|
| Isobutyl alcohol (mg/L) | 31.87 ± 2.92 a | 47.29 ± 6.01 b | 72.35 ± 9.53 c | 43.62 ± 7.23 b | 50.251 ± 7.48 b | 106.57 ± 11.05 d | * | 40 |
| Isoamyl alcohol (mg/L) | 295.05 ± 28.92 a | 411.09 ± 45.14 bc | 524.72 ± 65.58 d | 355.38 ± 13.66 b | 448.63 ± 56.93 c | 645.46 ± 72.21 e | * | 60 |
| 2-Phenylethanol (mg/L) | 62.26 ± 8.29 a | 72.11 ± 8.59 b | 75.47 ± 11.07 b | 91.04 ± 9.29 c | 87.55 ± 8.66 c | 106.83 ± 5.67 d | * | 10 |
| Total higher alcohols (mg/L) | 369.04 ± 61.55 a | 511.30 ± 48.09 b | 680.04 ± 82.44 c | 428.89 ± 63.66 a | 587.17 ± 71.45 b | 838.11 ± 117.40 d | * | |
| Total C6 alcohols (mg/L) | 0.49 ± 0.01 ab | 0.54 ± 0.01 ab | 0.62 ± 0.18 bc | 0.47 ± 0.03 a | 0.54 ± 0.01 ab | 0.74 ± 0.21 c | * | |
| Ethyl acetate (mg/L) | 48.84 ± 3.83 a | 37.40 ± 2.85 b | 34.87 ± 4.53 b | 67.40 ± 9.09 c | 37.68 ± 3.69 b | 34.21 ± 2.56 b | * | 7.50 |
| Isoamyl acetate (mg/L) | 1.79 ± 0.24 a | 0.88 ± 0.09 b | 0.65 ± 0.101 c | 3.51 ± 0.17 d | 0.89 ± 0.13 b | 1.00 ± 0.09 b | * | 0.16 |
| Phenethyl acetate (mg/L) | 0.59 ± 0.08 a | 0.31 ± 0.04 b | 0.23 ± 0.03 c | 0.76 ± 0.13 d | 0.35 ± 0.03 b | 0.37 ± 0.05 b | * | 0.25 |
| Total acetate esters (mg/L) | 51.29 ± 3.98 a | 38.64 ± 2.95 b | 35.81 ± 4.63 b | 73.29 ± 10.90 c | 38.97 ± 3.84 b | 35.53 ± 2.84 b | * | |
| Hexanoic acid (mg/L) | 47.78 ± 5.83 a | 29.70 ± 3.08 b | 34.981 ± 4.42 c | 41.32 ± 0.85 d | 26.98 ± 2.45 be | 24.45 ± 2.22 e | * | 0.42 |
| Octanoic acid (mg/L) | 98.59 ± 3.78 a | 58.06 ± 6.93 b | 42.565 ± 4.97 c | 73.63 ± 4.03 d | 43.30 ± 5.26 c | 22.13 ± 3.26 e | * | 0.5 |
| Decanoic acid (mg/L) | 77.23 ± 3.60 a | 48.48 ± 5.62 b | 30.48 ± 4.27 c | 42.43 ± 4.47 d | 25.34 ± 3.59 e | 11.79 ± 0.85 f | * | 1 |
| Isobutyric acid (mg/L) | 4.35 ± 0.68 a | 5.04 ± 0.75 b | 5.07 ± 0.46 b | 3.76 ± 0.44 c | 4.88 ± 0.55 ab | 4.93 ± 0.49 ab | * | 3 |
| Total medium-chain fatty acids (mg/L) | 224.91 ± 3.88 a | 140.44 ± 15.37 b | 106.24 ± 14.69 c | 165.37 ± 5.38 d | 100.50 ± 11.09 e | 58.13 ± 3.43 c | * | |
| Ethyl hexanoate (μg/L) | 335.98 ± 31.38 a | 219.4 ± 22.01 b | 162.94 ± 20.31 c | 528.81 ± 67.73 d | 205.82 ± 27.75 b | 211.26 ± 28.85 b | * | 80 |
| Ethyl octanoate (μg/L) | 619.74 ± 74.42 a | 397.47 ± 53.15 b | 351.04 ± 55.95 b | 395.2 ± 93.44 b | 408.8 ± 52.02 b | 380.99 ± 53.86 b | * | 2 |
| Ethyl decanoate (mg/L) | 1.93 ± 0.22 a | 1.49 ± 0.21 bc | 1.56 ± 0.21 b | 1.02 ± 0.08 d | 1.33 ± 0.15 c | 1.53 ± 0.18 b | * | 0.2 |
| Total ethyl esters (mg/L) | 3.49 ± 0.27 a | 2.45 ± 0.30 bc | 2.69 ± 0.24 b | 1.92 ± 0.11 d | 2.28 ± 0.35 c | 2.53 ± 0.29 bc | * | |
| Methyl octanoate (mg/L) | 6.16 ± 0.83 a | 5.68 ± 0.60 b | 3.61 ± 0.46 cd | 3.32 ± 0.35 c | 3.54 ± 0.19 cd | 3.92 ± 0.47 d | * | 0.2 |
| β-Damascenone (μg/L) | 218.57 ± 12.02 ab | 231.98 ± 25.61 abc | 205.01 ± 19.47 a | 259.31 ± 12.93 c | 247.94 ± 21.44 c | 238.19 ± 35.1 bc | * | 0.14 |
| 3-(Methylthio)-1-propanol (mg/L) | 1.14 ± 0.14 a | 1.72 ± 0.12 b | 1.73 ± 0.22 b | 2.96 ± 0.38 c | 1.75 ± 0.24 b | 3.68 ± 0.24 d | * | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, P.; Ivanova-Petropulos, V.; Duan, C.; Yan, G. Effect of Unsaturated Fatty Acids on Intra-Metabolites and Aroma Compounds of Saccharomyces cerevisiae in Wine Fermentation. Foods 2021, 10, 277. https://doi.org/10.3390/foods10020277
Liu P, Ivanova-Petropulos V, Duan C, Yan G. Effect of Unsaturated Fatty Acids on Intra-Metabolites and Aroma Compounds of Saccharomyces cerevisiae in Wine Fermentation. Foods. 2021; 10(2):277. https://doi.org/10.3390/foods10020277
Chicago/Turabian StyleLiu, Peitong, Violeta Ivanova-Petropulos, Changqing Duan, and Guoliang Yan. 2021. "Effect of Unsaturated Fatty Acids on Intra-Metabolites and Aroma Compounds of Saccharomyces cerevisiae in Wine Fermentation" Foods 10, no. 2: 277. https://doi.org/10.3390/foods10020277
APA StyleLiu, P., Ivanova-Petropulos, V., Duan, C., & Yan, G. (2021). Effect of Unsaturated Fatty Acids on Intra-Metabolites and Aroma Compounds of Saccharomyces cerevisiae in Wine Fermentation. Foods, 10(2), 277. https://doi.org/10.3390/foods10020277