3.1. Rat Food Intake, Body Weight and Fecal Nitrogen
Rats fed the blackcurrant and green tea diet with pectin (8%) had significantly lower food intake compared to those on other diets (
p < 0.001) (
Table 2). After 28 days, rat final body weights were similar between the diet groups (
Table 2). However, rats fed blackcurrant and green tea diets gained less body weight than those rats fed the control diet. The reduction in rat food intake and weight gain with blackcurrant and green tea is likely due to the astringent or bitter properties of the polyphenols. The precipitation of salivary proteins on the tongue by polyphenols results in a feeling of constriction, roughness and dryness on the palate, therefore reducing the palatability of the diet to the rats [
15]. Additionally, non-digestible carbohydrates and polyphenols can influence the neuropeptides involved in satiety [
16]. Although the underlying mechanisms remain unknown in the current study, combining a blackcurrant and green tea diet with high soluble fiber (8% pectin) was effective in reducing rat food intake.
Protein intake was lower in rats fed a blackcurrant and green tea diet (
p = 0.021) (
Table 3). Rat fecal nitrogen outputs were significantly higher in blackcurrant and green tea diets with pectin (4 or 8%) when compared to the control diet (
p < 0.001) (
Table 3). The apparent nitrogen balance and protein digestibility measures were significantly different between the experimental diet groups (
p < 0.001). Rats fed a blackcurrant and green tea diet with 8% pectin had high fecal nitrogen output and low protein digestibility, indicating that polyphenols may have bound to the dietary and endogenous proteins such as the digestive proteases, affecting protease activity and subsequently protein digestibility in the gut [
17].
3.2. Polyphenols in Urine, Feces and Plasma of Rats
The absorption and metabolism of polyphenols was determined by the presence of polyphenol parent components and their metabolites in biological samples (urine, feces and plasma) from rats fed blackcurrant and green tea diets with or without pectin (
Table 4). Relative peak areas of polyphenols detected in rat urine, feces and plasma are presented in
Supplementary Tables S1–S3.
There were no catechins, anthocyanins, flavonols or phenolic acids detected in the control diet and those rats fed the control diet. For the rats given the blackcurrant and green tea diets, a total of eight catechins, five anthocyanins and four flavonols were identified.
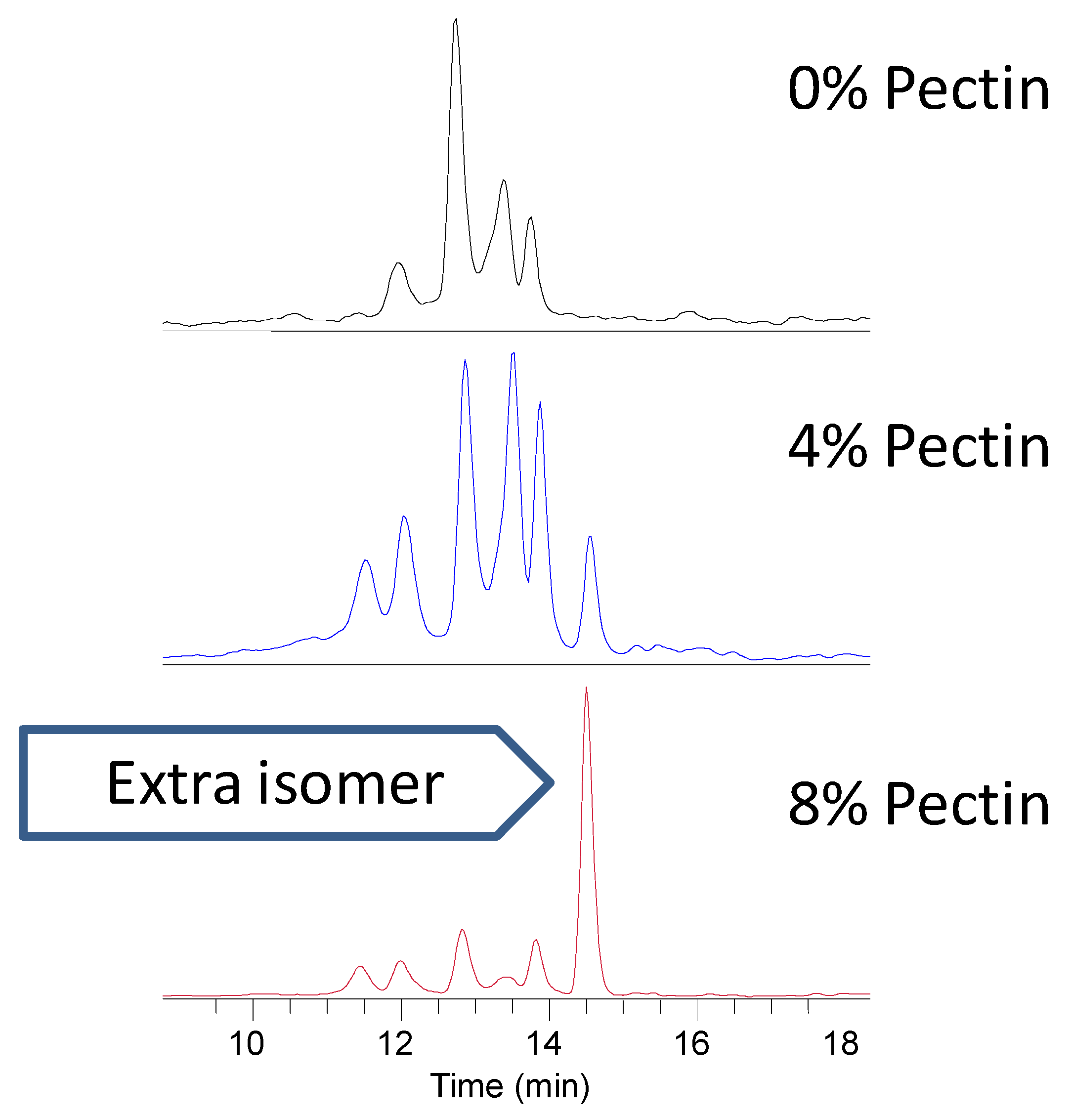
Parent catechins and flavonols were not detected in the urine obtained from rats fed the blackcurrant and green tea diets, but there were parent anthocyanins present. Catechin, anthocyanin and flavonol metabolites were present in the rat urine. Adding pectin to the blackcurrant and green tea diet affected the digestion and absorption of catechins, as shown by their presence or absence in the urine. The inclusion of soluble fiber in the diet may have promoted further metabolism via methylation, as shown by the additional methyl gallocatechin glucuronide isomer and variable isomer profiles with the addition of 4 and 8% pectin to the blackcurrant and green tea diets (
Figure 1). In the feces, catechins were only present as the parent compounds, while for the anthocyanins, there were both parent compounds and metabolites. Catechin gallate and epicatechin were not found in the feces of rats fed blackcurrant and green tea with 8% pectin. Petunidin and peonidin rutinosides were only found in the feces of rats that consumed blackcurrant and green tea without pectin. Two phenolic acids (3, 4 and 2, 4-dihydroxybenzoic acid) were present in the feces and urine of rats fed blackcurrant and green tea diets with and without pectin, but absent in the blood plasma. In the plasma, we found no anthocyanin parent compounds or their metabolites in rats fed blackcurrant and green tea diets. The only catechin parent compound present in the plasma was catechin. Gallocatechin glucuronide was present in the plasma of rats fed a blackcurrant and green tea diet without pectin but was absent in rats fed the diets supplemented with 4 and 8% pectin. The only flavonol parent compound present in the plasma was quercetin rutinoside.
Catechins are one of the important polyphenols found in tea and include an abundance of epigallocatechin gallate and epigallocatechin. Previous studies in animals and humans have shown that catechins have low absorption rates and therefore reduced bioavailability in blood circulation [
18]. When substantial quantities of catechins pass from the small intestine to the large intestine, they can be catabolized by the bacteria to produce phenolic acids before being excreted in the feces. This could be the reason for the absence of epicatechin and catechin gallate in the feces of rats that consumed a blackcurrant and green tea diet containing 8% pectin. Similarly, there were some anthocyanins detected in the urine but not the feces of rats fed the diets containing pectin (4 or 8%). The anthocyanin-rich blackcurrant extract in the current study contained mainly delphinidin-3-glucoside, delphinidin-3-rutinoside, cyanidin-3-glucoside and cyanidin-3-rutinoside [
8]. With some exceptions, anthocyanins appeared to be largely absorbed into the blood circulation, removed by the kidneys and consequently detected in the rat urine.
Inclusion of dietary fiber in a blackcurrant and green tea diet offers potential health benefits. This is evident from the observed presence and absence of polyphenols observed in the urine, feces and plasma of these rats. The addition of pectin, a fermentable fiber, affected the anthocyanin, catechin and flavonol metabolism, as indicated by the absence of some of the parent compounds and their metabolites in the urine, feces and plasma of rats fed diet with 4 and 8% pectin. Most polyphenols display limited bioavailability, but adding pectin to the diet can improve the polyphenol absorption by delaying the gastric emptying, which extends absorption in the small intestine, therefore enhancing the bioavailability of polyphenols [
19,
20,
21]. Polyphenols that escape digestion in the small intestine reach the large intestine, where they can act as a nutrient source to some resident microorganisms, thus inducing changes in the microbiota composition. There is growing evidence in the literature that polyphenols are extensively metabolized and further converted by the gut microbiota into bioactive molecules that are absorbed through the gut barrier, entering blood circulation and peripheral organs, contributing to host physiological functions [
16,
22]. In previous studies, we found that blackcurrant and pectin altered the microbiota composition and increased short-chain fatty acids (SCFAs) concentrations in the gut, which can impact overall health and wellbeing [
8,
23].
It is worth mentioning that food source (cultivar, growing environment), chemical structure, conjugation, food matrix, food processing conditions, interactions with other compounds and host physiology can influence the bioavailability of polyphenols [
3]. Future studies should investigate the role of the gut microbiota in polyphenol transformation and their contributions to health promotion. It also remains unknown how the polyphenol absorption and metabolism are affected by the formation of chemical complexes and colloidal structures between the polyphenols and dietary fiber within the food matrix and during the processes of digestion and absorption.
3.3. Plasma Corticosterone, TRAP and Uric Acid
There was no statistically significant difference in corticosterone concentrations between the diet groups, but there was a marked decrease in corticosterone concentrations in the rats fed blackcurrant and green tea diets containing pectin (4% or 8%) (
Table 5). Polyphenols and non-digestible carbohydrate components in the diet can influence corticosterone levels. An earlier study in rats has reported the normalization of corticosterone concentrations by polyphenols [
24]. Similarly, saccharolytic fermentation of dietary fiber occurs in the gut, releasing SCFAs that can also lower corticosterone in blood plasma [
25].
Plasma TRAP concentrations tended to be higher in rats fed blackcurrant and green tea diets with either 4 or 8% pectin compared to the control diet (
p = 0.046) (
Table 6). The TRAP assay has been widely used to determine the antioxidant potential of food constituents [
26]. Amongst fruit, high antioxidant capacity was found in berries, including blackcurrants [
27,
28]. In the current study, the combination of phenolic-rich plant extracts and pectin enhanced the plasma antioxidant capacity in rats. Further research in humans is needed to substantiate the health-promoting properties of the dietary combination of blackcurrant, green tea and pectin.
Endogenous plasma antioxidants are responsible for the homeostatic regulation of antioxidant status in blood. Uric acid is thought to contribute the greatest effect on TRAP [
29]. In the present study, there was no correlation between TRAP and uric acid measurements (R
2 = 0.05,
p = 0.756). Plasma uric acid concentrations did not differ significantly between the diet groups (
Table 6). However, rats that consumed a diet with 4 or 8% pectin tended to have lower uric acid, highlighting the beneficial role of dietary fiber in suppressing uric acid concentrations [
30].


 ,
, 


{kind=link}
{kind=link}