
Does the Food Ingredient Pectin Provide a Risk for Patients Allergic to Non-Specific Lipid-Transfer Proteins?

 , , , , , and
, , , , , and 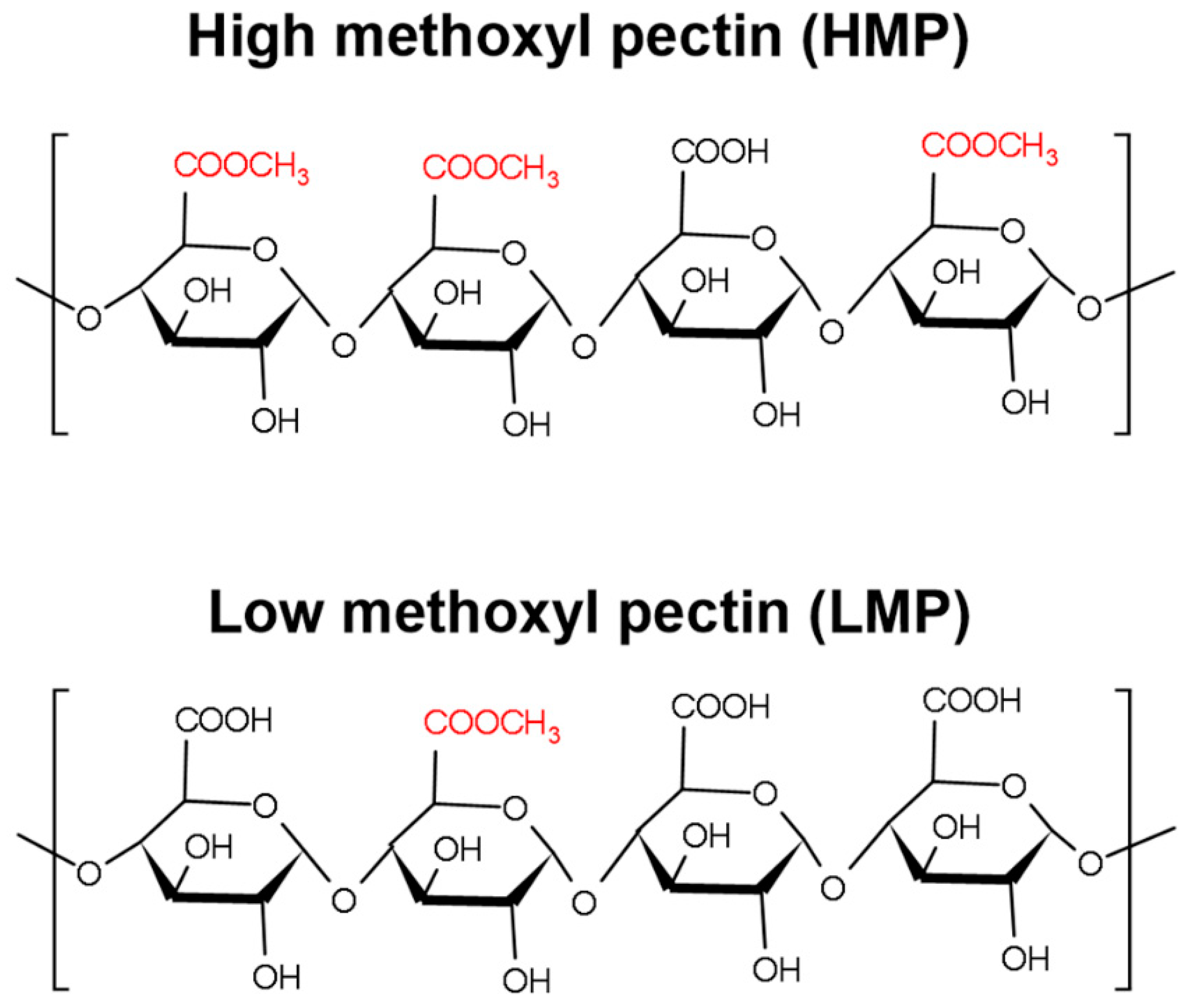{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
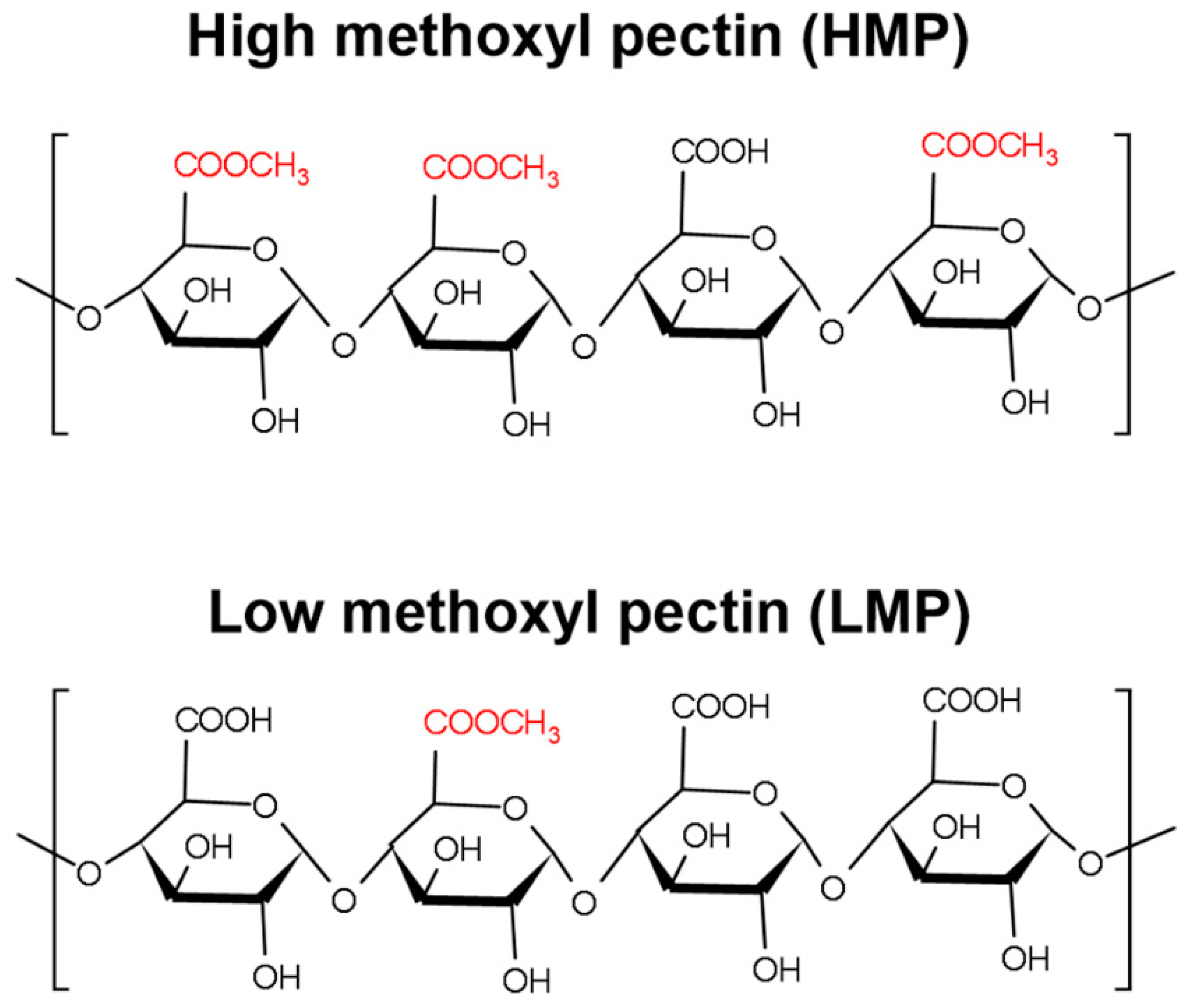
2.1. Material
2.2. Methods
2.2.1. Pectin Sample Preparation
2.2.2. Estimation of the Analytical nsLTP Threshold
2.2.3. Pru p 3 Specific IgE ELISA
2.2.4. SDS-PAGE and Immunoblotting
2.2.5. Size Exclusion Chromatography
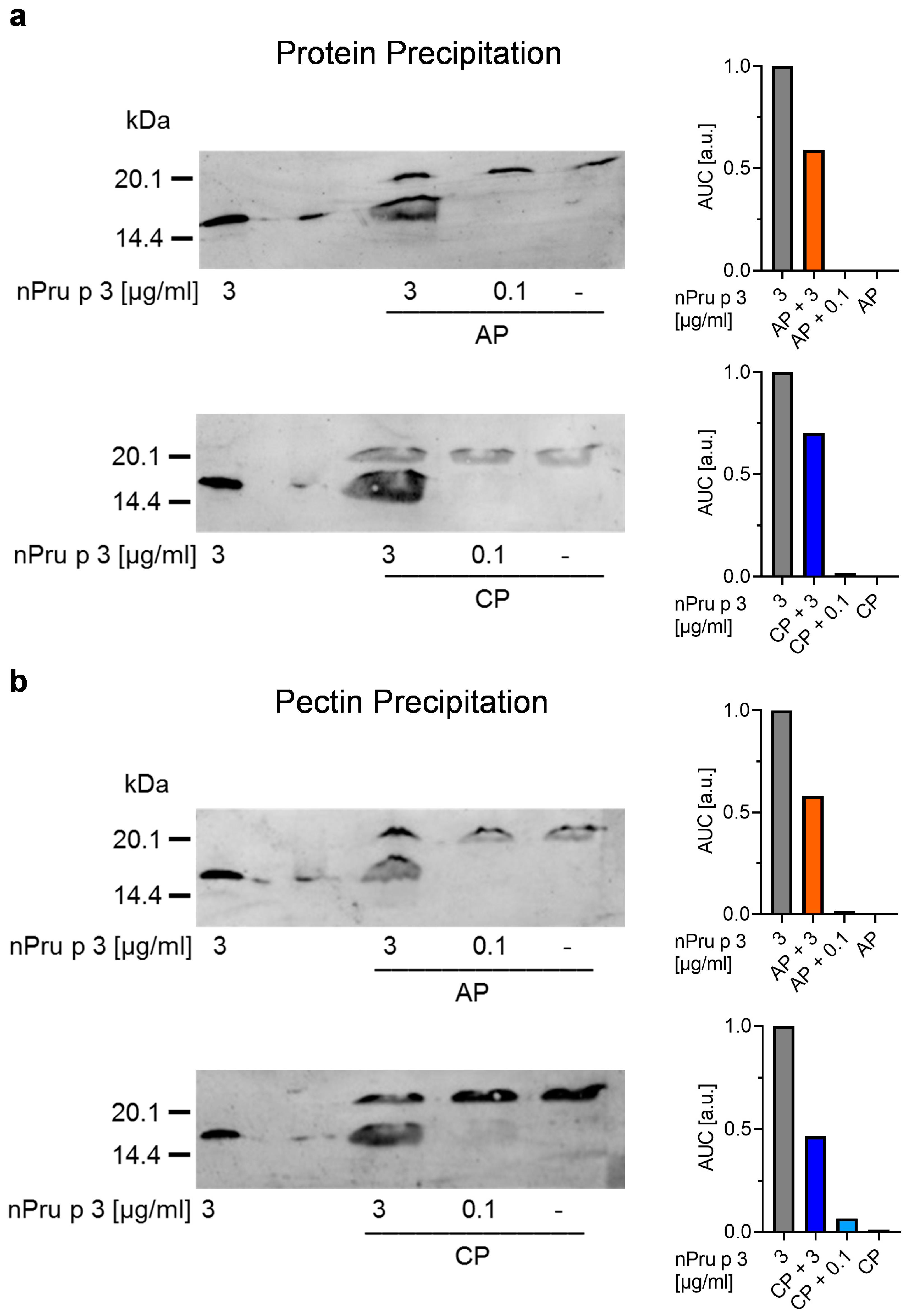
2.2.6. Protein Precipitation
2.2.7. Pectin Precipitation
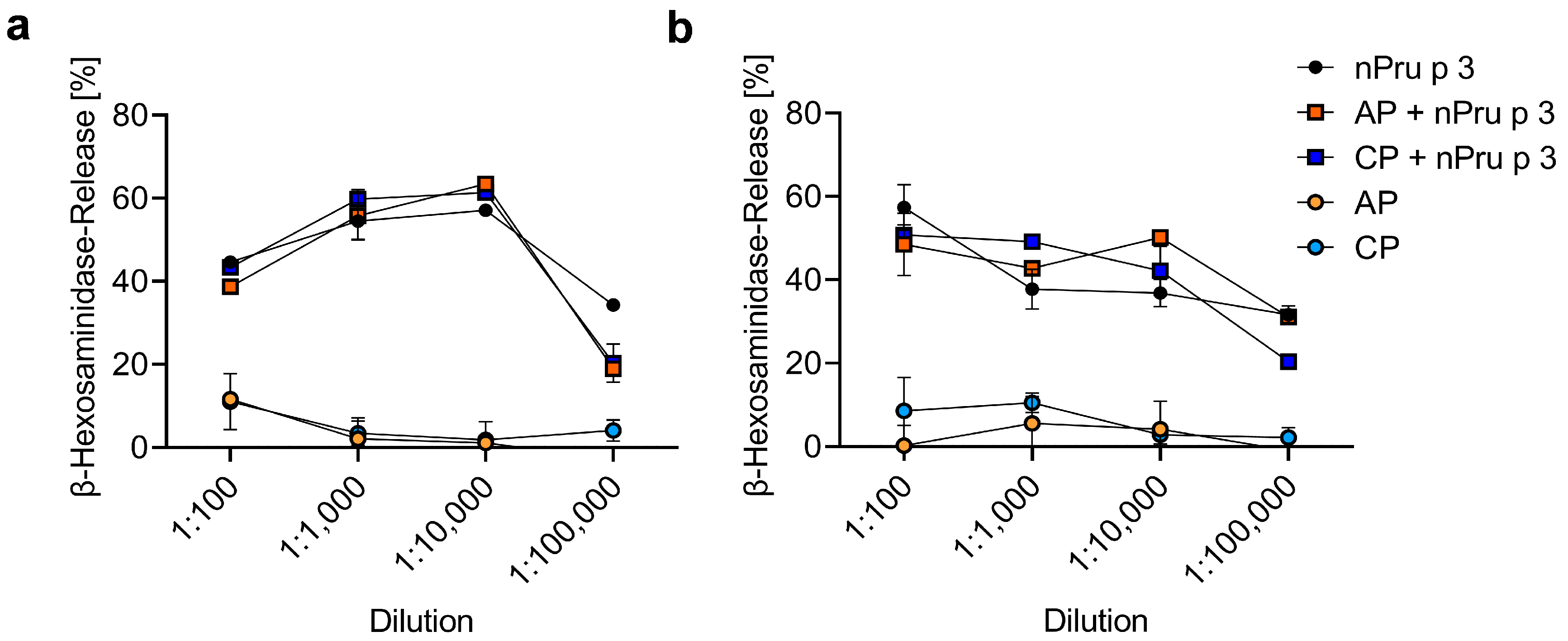
2.2.8. β-Hexosaminidase Release from Humanized Rat Basophil Leukemia (huRBL) Cells
2.2.9. Statistical Analysis
3. Results
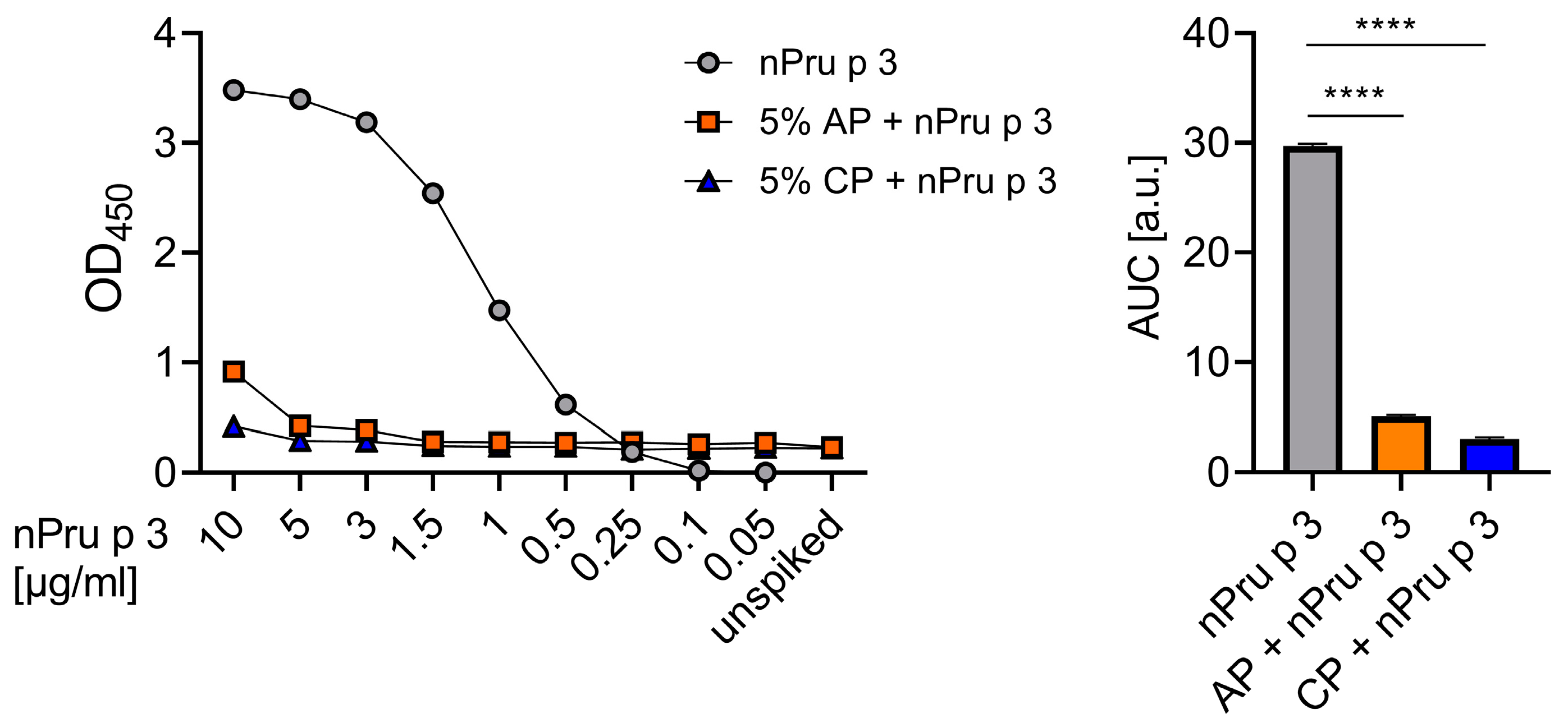
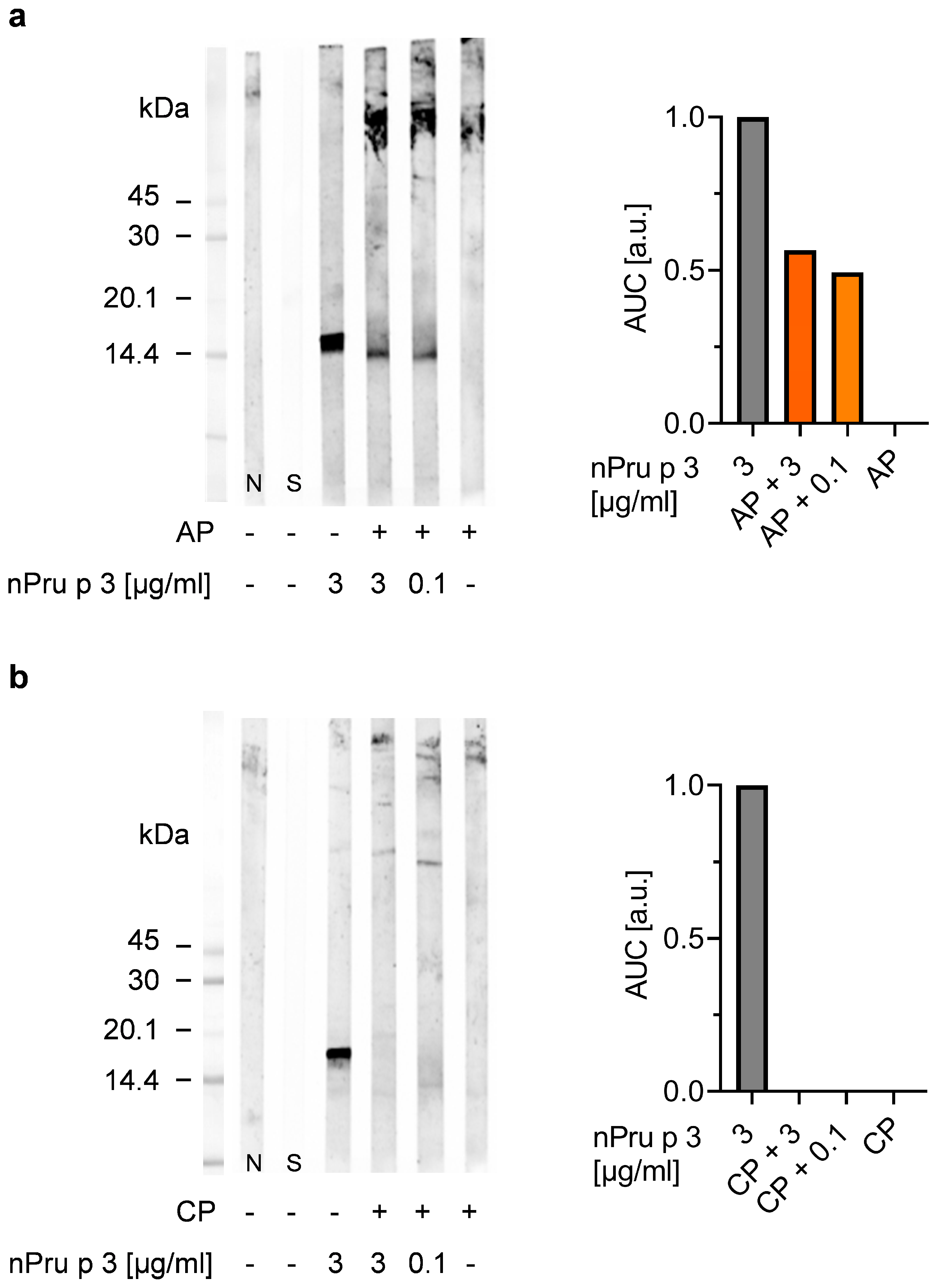
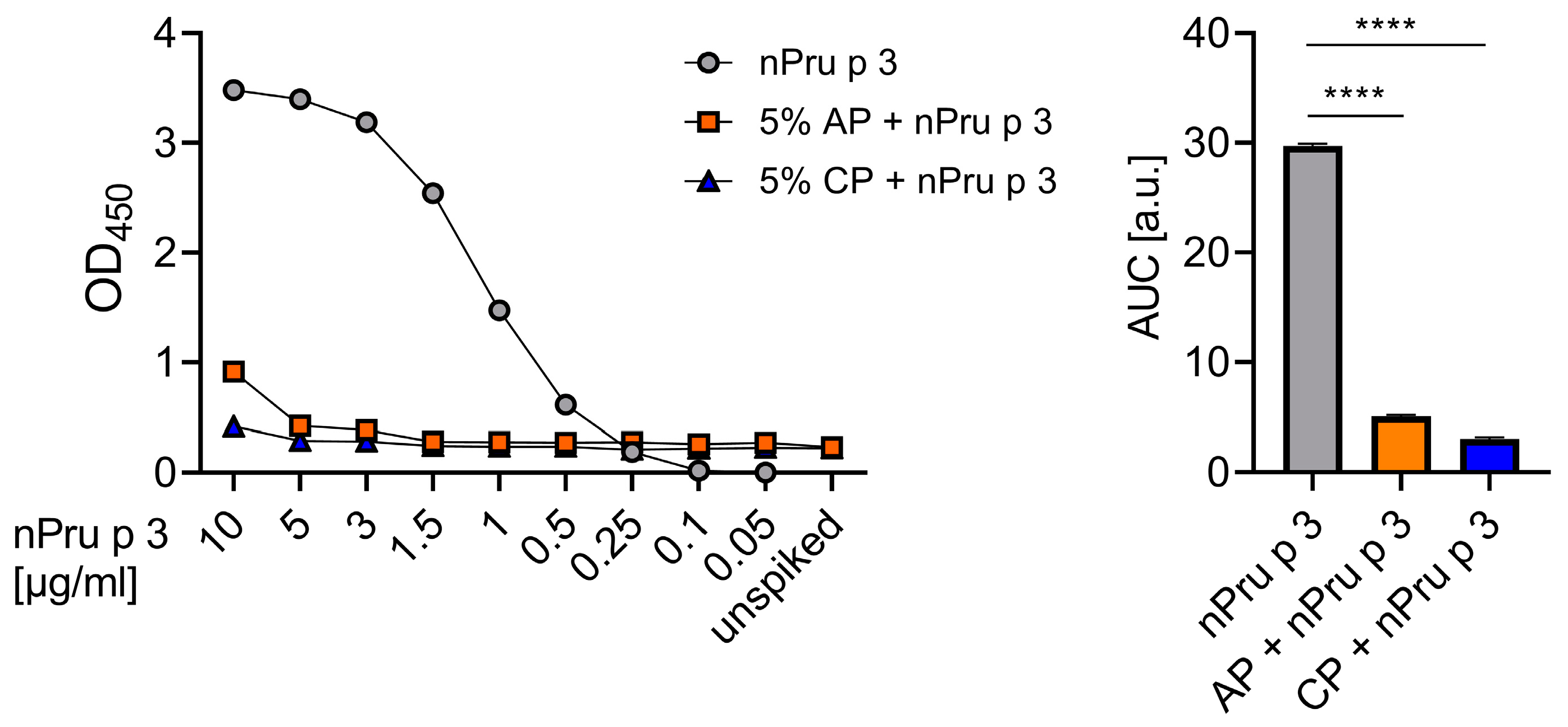
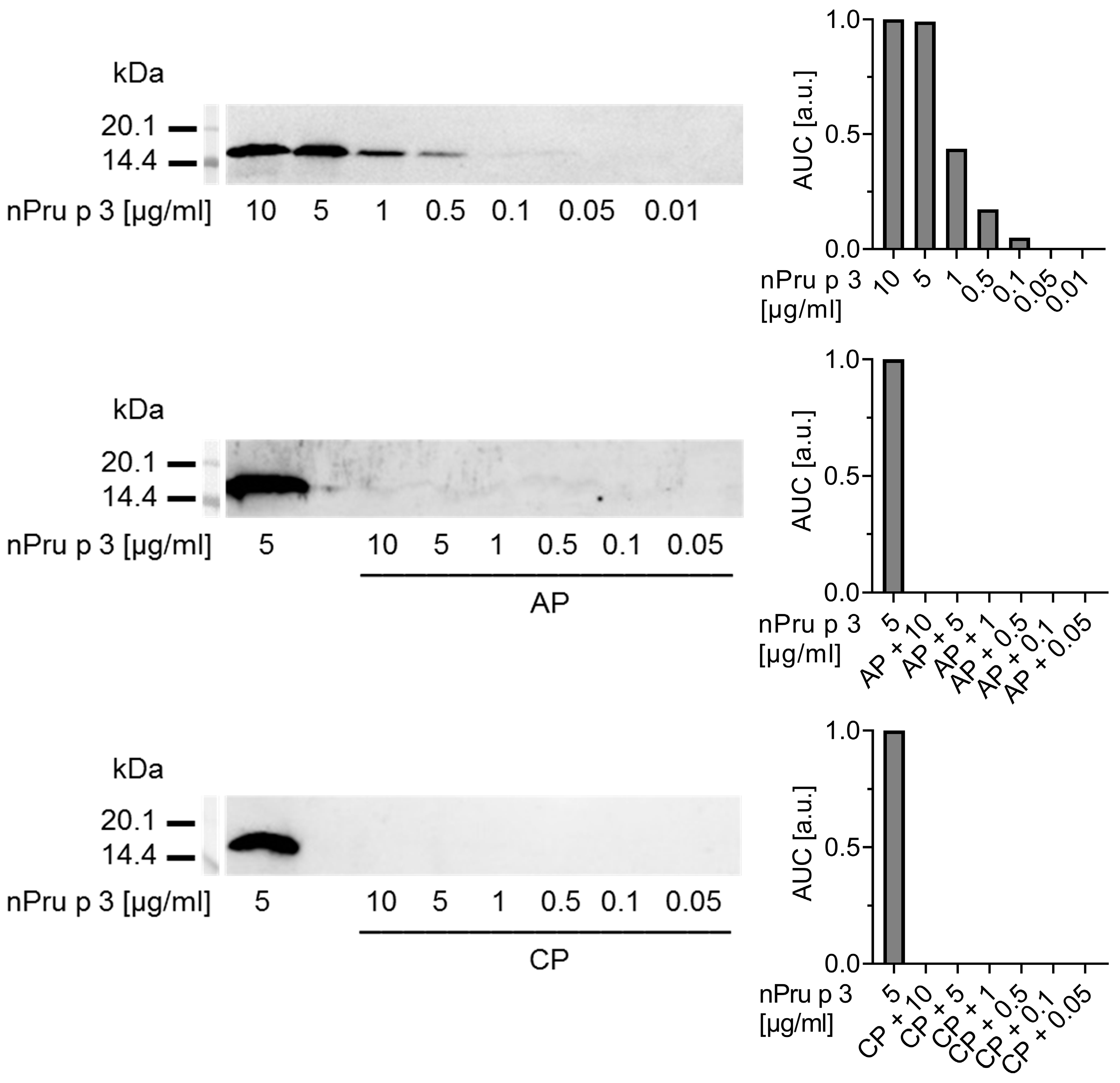
3.1. Detection of nsLTP Is Hampered in the Pectin Matrix
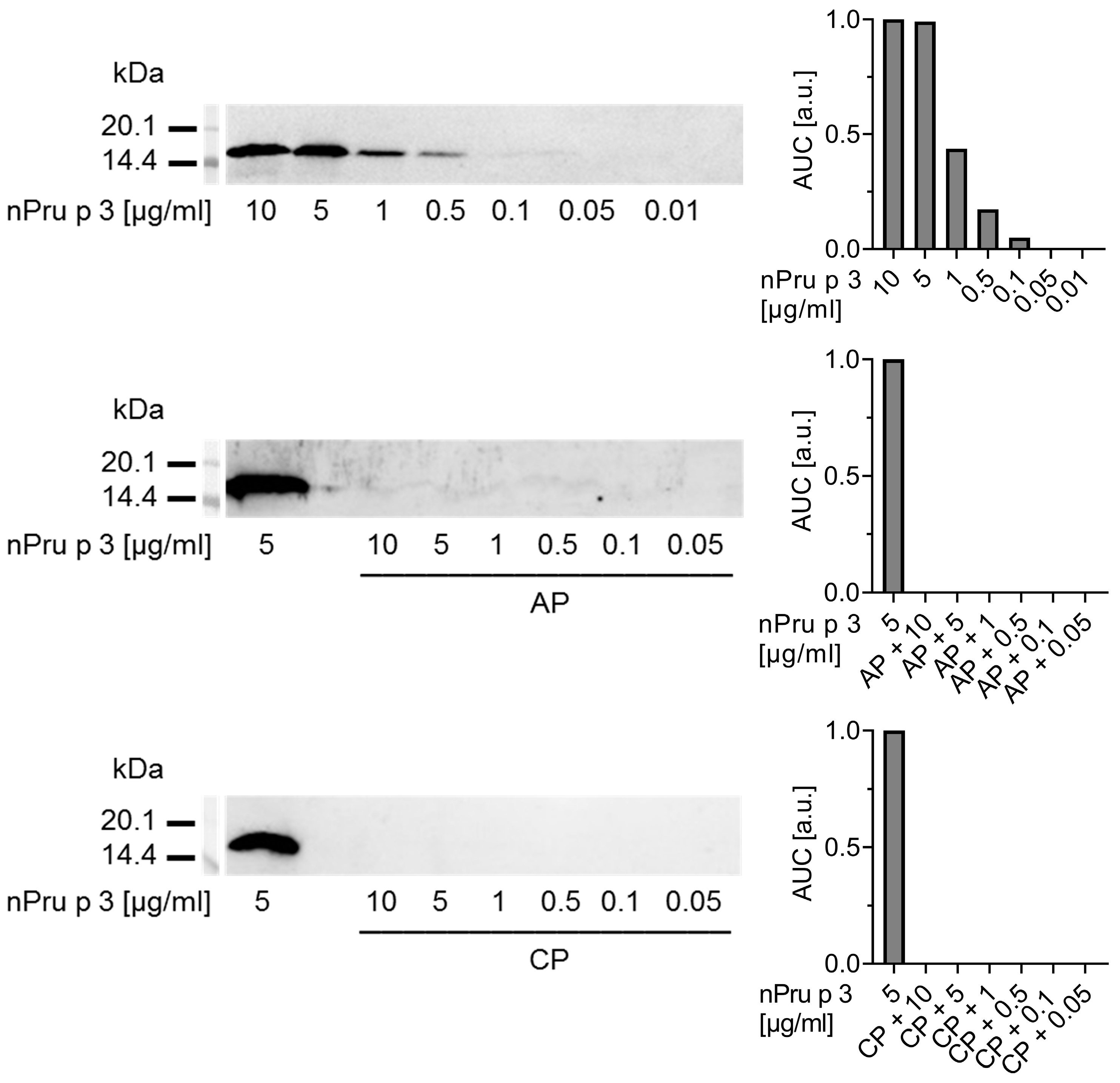
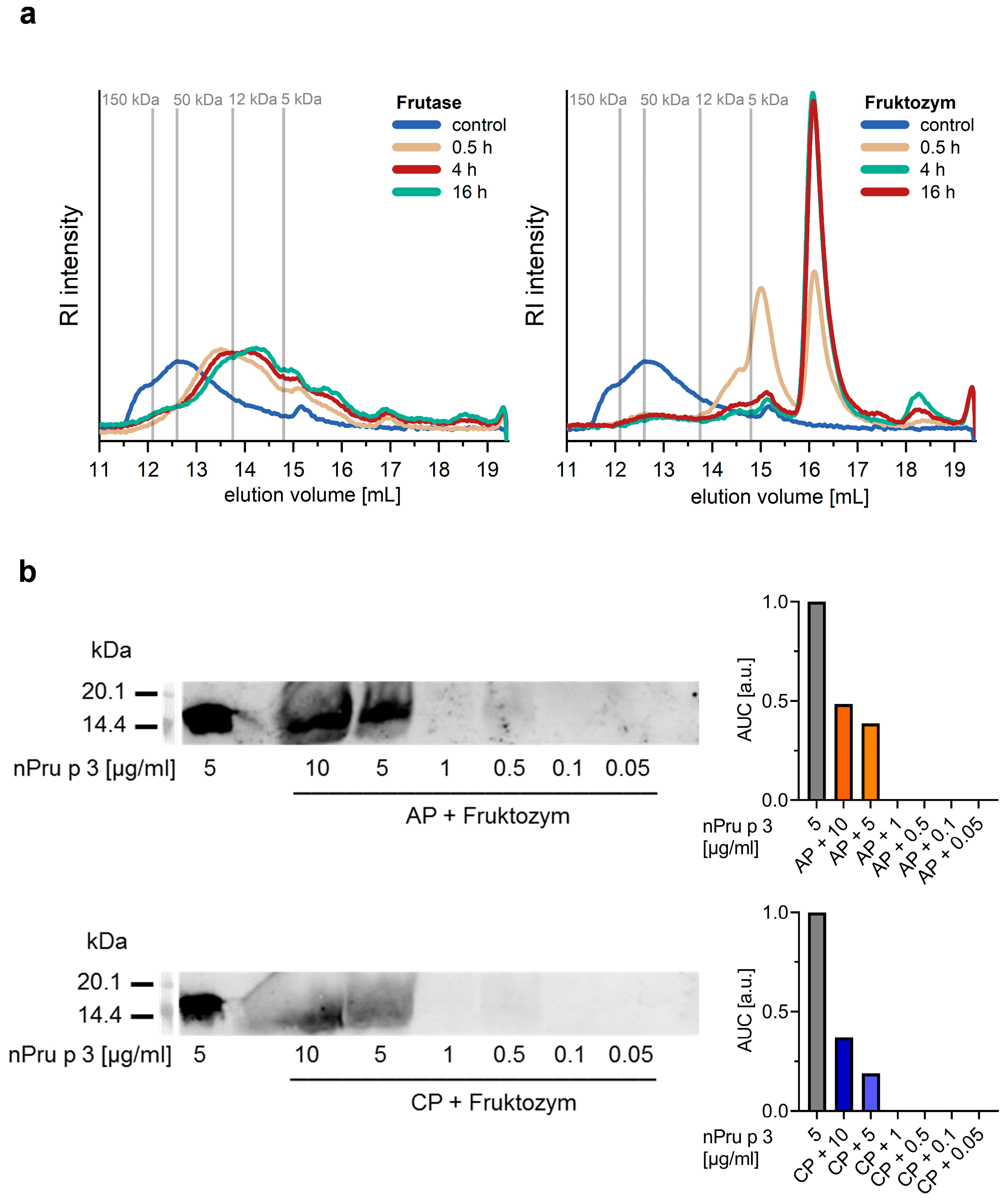
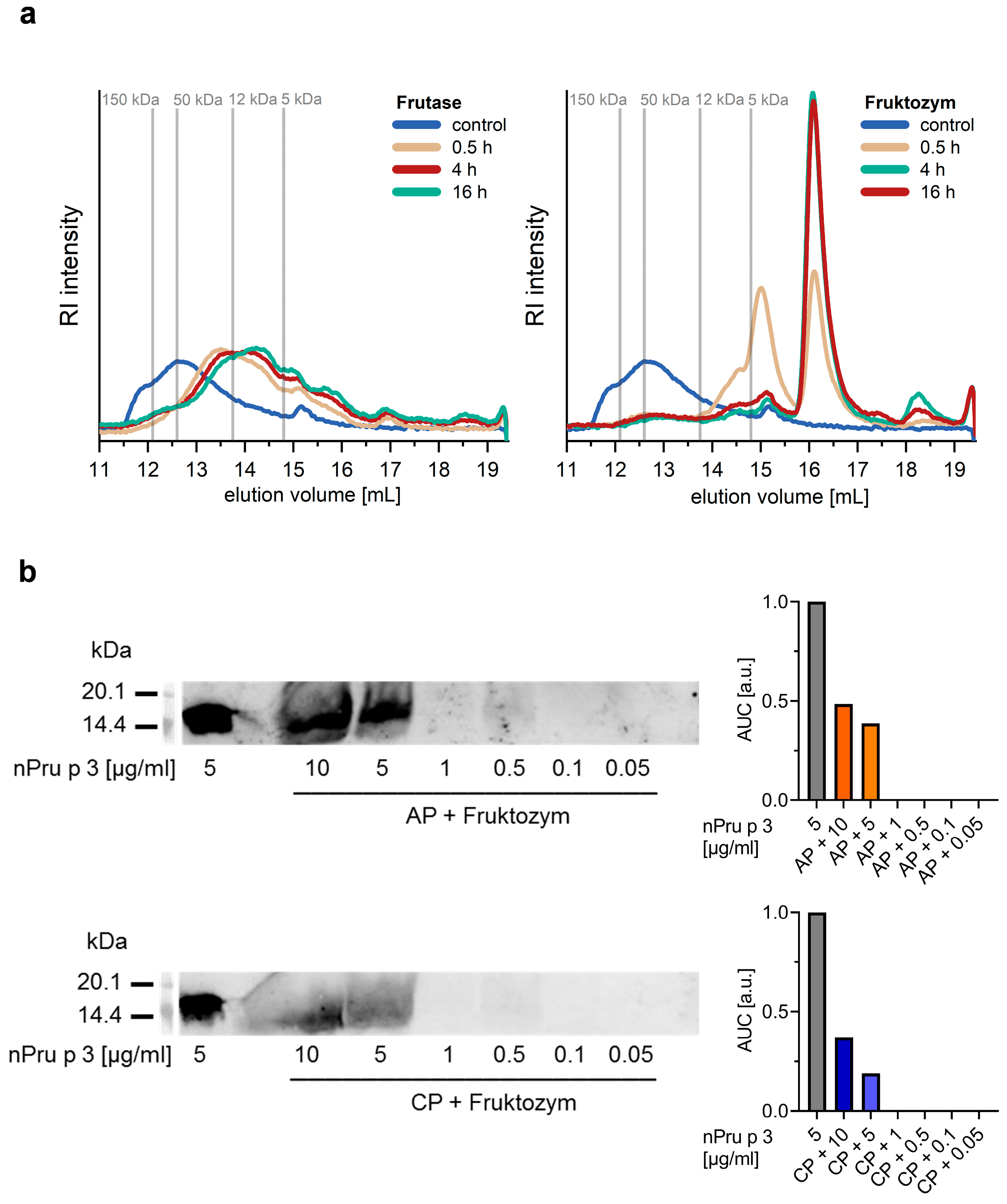
3.2. Enzymatic Pre-Treatment of Pectin Increases the Analytical Sensitivity for nsLTP Detection
3.3. Enzymatic Treatment of AP Allows Detection of nsLTP Corresponding to Clinical Thresholds
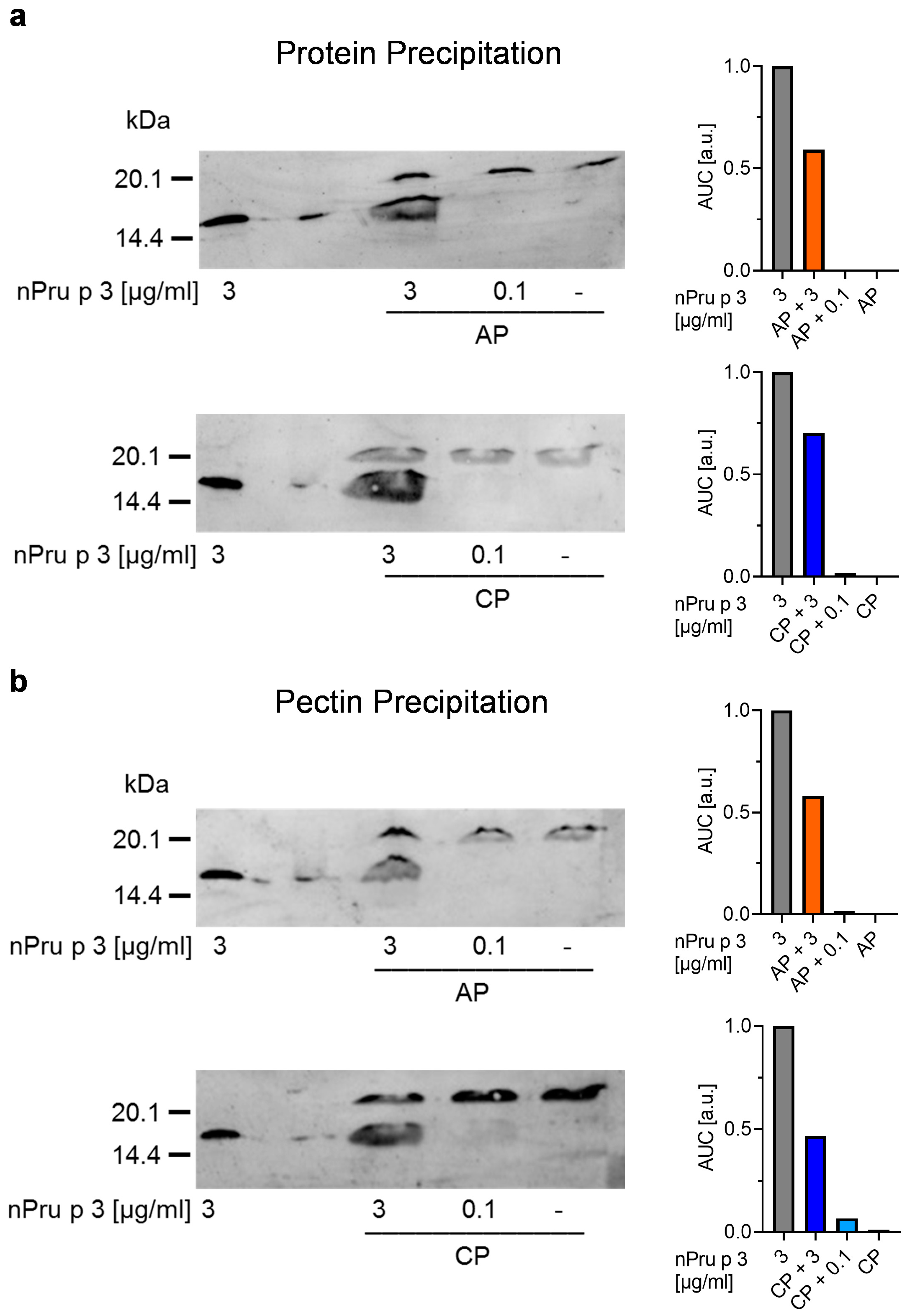
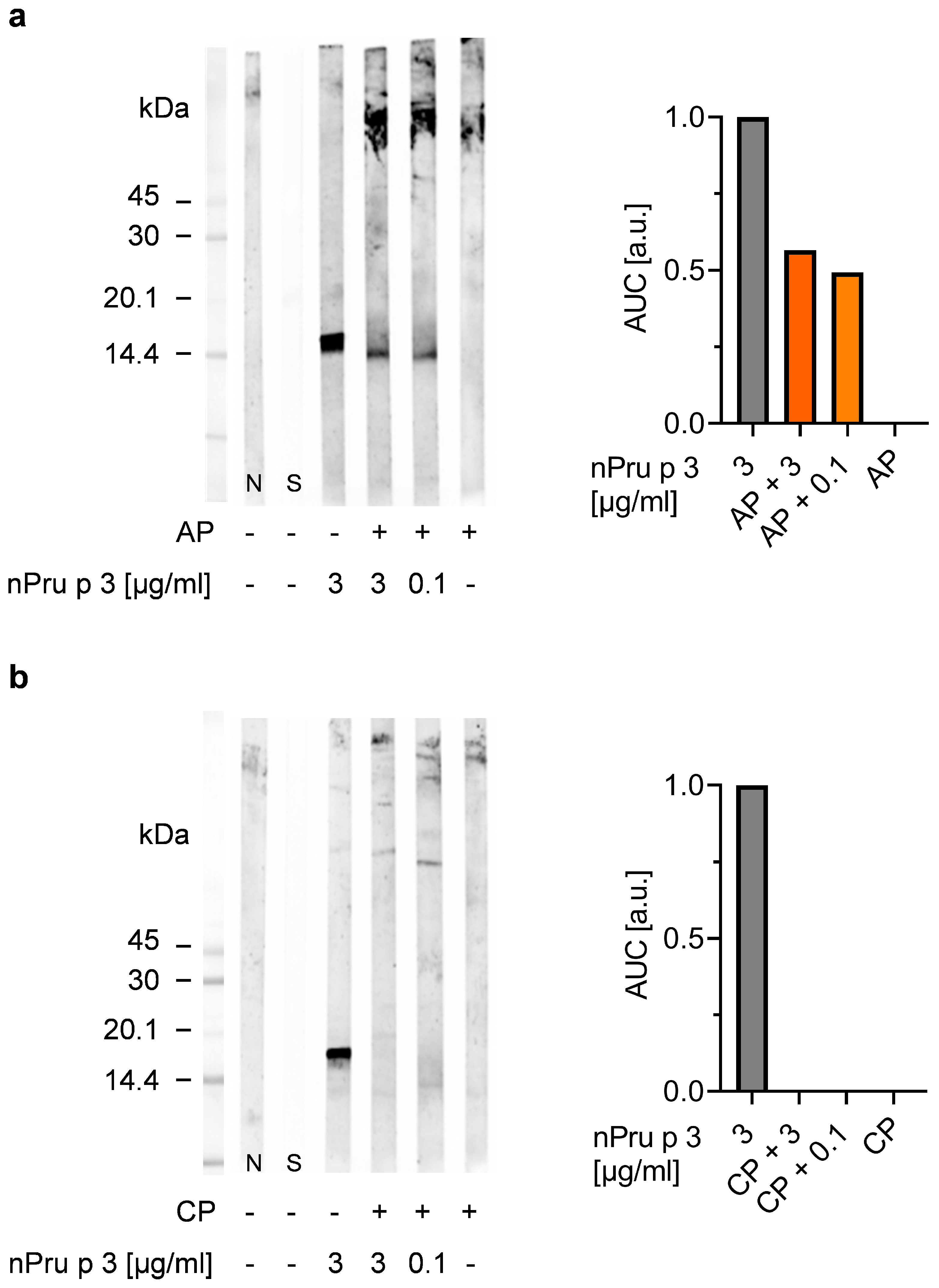
3.4. Precipitation of Protein and Pectin Allows Detection of nsLTP Concentration in Pectin Corresponding to Threshold for Anaphylactic Reactions
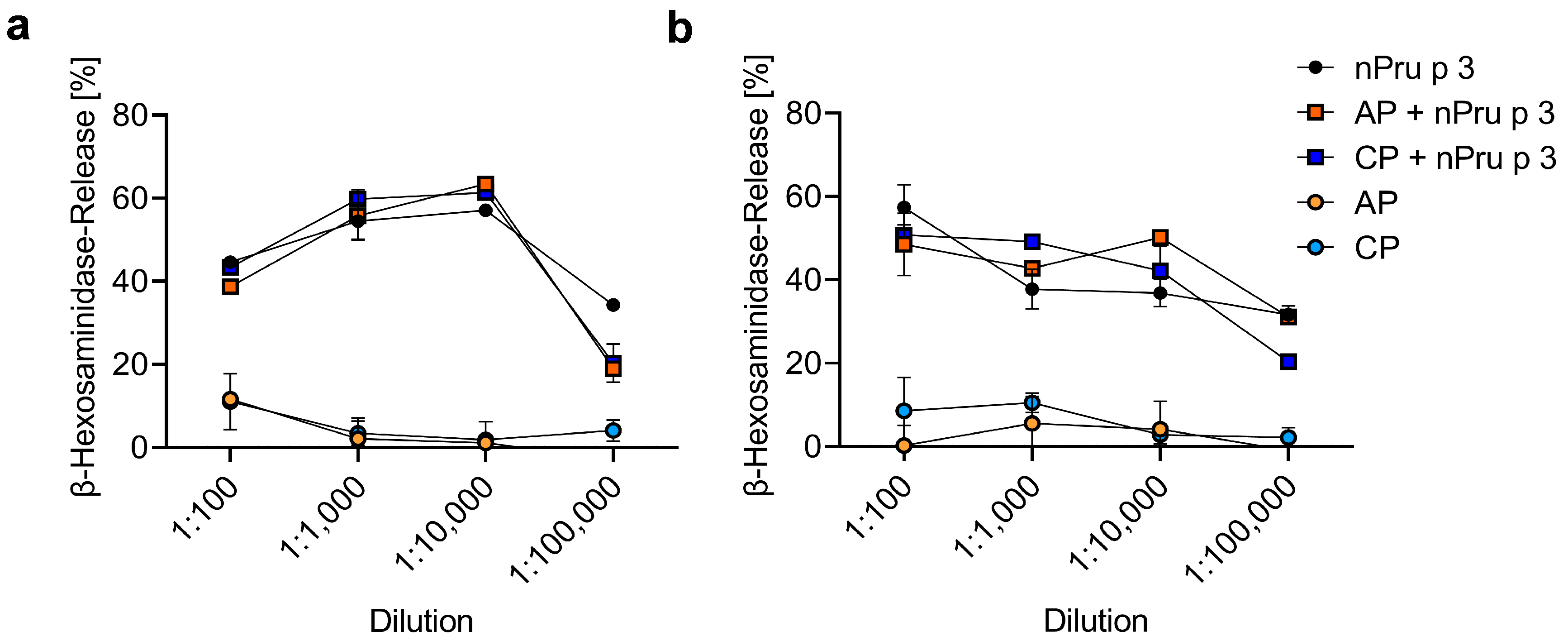
3.5. huRBL Assay Allows Detection of Spiked nPru p 3 Concentrations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AP | apple-derived pectin |
| CP | citrus-derived pectin |
| DE | degree of esterification |
| FCS | fetal calf serum |
| GalA | galacturonic acid |
| HG | homogalacturonan |
| HMP | high methoxyl pectin |
| LMP | low methoxyl pectin |
| MW | molecular weight |
| nPru p 3 | natural Pru p 3 |
| nsLTP | non-specific lipid transfer protein |
| OAS | oral allergy syndrome |
| RG | rhamnogalacturonan |
| XGA | xylogalacturonan |
References
- Chen, J.; Liang, R.; Liu, W.; Li, T.; Liu, C.; Wu, S.; Wang, Z. Pectic-oligosaccharides prepared by dynamic high-pressure microfluidization and their in vitro fermentation properties. Carbohydr. Polym. 2013, 91, 175–182. [Google Scholar] [CrossRef]
- Mualikrishna, G.; Tharanathan, R.N. Characterization of pectic polysaccharides from pulse husks. Food Chem. 1994, 50, 87–89. [Google Scholar] [CrossRef]
- Cornuault, V.; Posé, S.; Knox, J.P. Disentangling pectic homogalacturonan and rhamnogalacturonan-I polysaccharides: Evidence for sub-populations in fruit parenchyma systems. Food Chem. 2018, 246, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Dranca, F.; Oroian, M. Extraction, purification and characterization of pectin from alternative sources with potential technological applications. Food Res. Int. 2018, 113, 327–350. [Google Scholar] [CrossRef] [PubMed]
- Beukema, M.; Jermendi, É.; Koster, T.; Kitaguchi, K.; de Haan, B.J.; van den Berg, M.A.; Faas, M.M.; Schols, H.A.; de Vos, P. Attenuation of doxorubicin-induced small intestinal mucositis by pectins is dependent on pectin’s methyl-ester number and distribution. Mol. Nutr. Food Res. 2021, 65, e2100222. [Google Scholar] [CrossRef]
- Wai, W.W.; Alkarkhi, A.F.M.; Easa, A.M. Optimization of pectin extraction from durian rind (Durio zibethinus) using response surface methodology. J. Food Sci. 2009, 74, C637-41. [Google Scholar] [CrossRef]
- Willats, W.G.; Knox, J.P.; Mikkelsen, J.D. Pectin: New insights into an old polymer are starting to gel. Trends Food Sci. Technol. 2006, 17, 97–104. [Google Scholar] [CrossRef]
- Vanitha, T.; Khan, M. Role of pectin in food processing and food packaging. In Pectins-Extraction, Purification, Characterization and Applications; Masuelli, M., Ed.; IntechOpen: London, UK, 2020; ISBN 978-1-78984-071-1. [Google Scholar]
- Laurent, M.A.; Boulenguer, P. Stabilization mechanism of acid dairy drinks (ADD) induced by pectin. Food Hydrocoll. 2003, 17, 445–454. [Google Scholar] [CrossRef]
- Shah, B.R.; Li, B.; Al Sabbah, H.; Xu, W.; Mráz, J. Effects of prebiotic dietary fibers and probiotics on human health: With special focus on recent advancement in their encapsulated formulations. Trends Food Sci. Technol. 2020, 102, 178–192. [Google Scholar] [CrossRef]
- Burkitt, D.; Walker, A.; Painter, N. Effect of dietary fibre on stools and transit-times, and its role in the causation of disease. Lancet 1972, 300, 1408–1411. [Google Scholar] [CrossRef]
- Schwartz, S.E.; Levine, R.A.; Singh, A.; Scheidecker, J.R.; Track, N.S. Sustained pectin ingestion delays gastric emptying. Gastroenterology 1982, 83, 812–817. [Google Scholar] [CrossRef]
- Chang, J.H.; Kim, M.S.; Kim, T.W.; Lee, S.S. Effects of soybean supplementation on blood glucose, plasma lipid levels, and erythrocyte antioxidant enzyme activity in type 2 diabetes mellitus patients. Nutr. Res. Pract. 2008, 2, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Shtriker, M.G.; Hahn, M.; Taieb, E.; Nyska, A.; Moallem, U.; Tirosh, O.; Madar, Z. Fenugreek galactomannan and citrus pectin improve several parameters associated with glucose metabolism and modulate gut microbiota in mice. Nutrition 2018, 46, 134–142.e3. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; He, Y.; Wang, F.; Zhang, H.; de Vos, P.; Sun, J. Low-methoxyl lemon pectin attenuates inflammatory responses and improves intestinal barrier integrity in caerulein-induced experimental acute pancreatitis. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Chung, W.S.F.; Meijerink, M.; Zeuner, B.; Holck, J.; Louis, P.; Meyer, A.S.; Wells, J.M.; Flint, H.J.; Duncan, S.H. Prebiotic potential of pectin and pectic oligosaccharides to promote anti-inflammatory commensal bacteria in the human colon. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Niu, Y.; Wu, H.; Sun, Y.; Li, Q.; Kong, X.; Liu, L.; Mei, Q. Modified apple polysaccharides could induce apoptosis in colorectal cancer cells. J. Food Sci. 2010, 75, H224–H229. [Google Scholar] [CrossRef]
- Zhang, Z.; Shi, L.; Pang, W.; Liu, W.; Li, J.; Wang, H.; Shi, G. Dietary fiber intake regulates intestinal microflora and inhibits ovalbumin-induced allergic airway inflammation in a mouse model. PLoS ONE 2016, 11, e0147778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Blanco-Pérez, F.; Steigerwald, H.; Schülke, S.; Vieths, S.; Toda, M.; Scheurer, S. The dietary fiber pectin: Health benefits and potential for the treatment of allergies by modulation of gut microbiota. Curr. Allergy Asthma Rep. 2021, 21, 1–19. [Google Scholar] [CrossRef]
- Washio, K.; Nakamura, M.; Sato, N.; Hori, M.; Matsubara, K.; Ogura, K.; Oda, Y.; Fukunaga, A.; Yagami, A.; Matsunaga, K. Anaphylaxis in a pectin- and cashew nut-allergic child caused by a citrus bath. Allergol. Int. 2021, 2, 1323–8930. [Google Scholar] [CrossRef]
- Ferdman, R.M.; Ong, P.Y.; Church, J.A. Pectin anaphylaxis and possible association with cashew allergy. Ann. Allergy Asthma Immunol. 2006, 97, 759–760. [Google Scholar] [CrossRef]
- Capucilli, P.; Kennedy, K.; Kazatsky, A.M.; Cianferoni, A.; Spergel, J.M. Fruit for thought: Anaphylaxis to fruit pectin in foods. J. Allergy Clin. Immunol. Pract. 2019, 7, 719–720. [Google Scholar] [CrossRef]
- Salminen, T.A.; Blomqvist, K.; Edqvist, J. Lipid transfer proteins: Classification, nomenclature, structure, and function. Planta 2016, 244, 971–997. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Rivas, M. The place of lipid transfer proteins (LTP) in the cross-reactivity of plant foods. Rev. Française D’allergologie 2009, 49, 433–436. [Google Scholar] [CrossRef]
- Skypala, I.J.; Asero, R.; Barber, D.; Cecchi, L.; Diaz Perales, A.; Hoffmann-Sommergruber, K.; Pastorello, E.A.; Swoboda, I.; Bartra, J.; Ebo, D.G.; et al. Non-specific lipid-transfer proteins: Allergen structure and function, cross-reactivity, sensitization, and epidemiology. Clin. Transl. Allergy 2021, 11, e12010. [Google Scholar] [CrossRef]
- Asero, R.; Antonicelli, L.; Arena, A.; Bommarito, L.; Caruso, B.; Crivellaro, M.; de Carli, M.; Della Torre, E.; Della Torre, F.; Heffler, E.; et al. EpidemAAITO: Features of food allergy in Italian adults attending allergy clinics: A multi-centre study. Clin. Exp. Allergy 2009, 39, 547–555. [Google Scholar] [CrossRef]
- Pascal, M.; Muñoz-Cano, R.; Reina, Z.; Palacín, A.; Vilella, R.; Picado, C.; Juan, M.; Sánchez-López, J.; Rueda, M.; Salcedo, G.; et al. Lipid transfer protein syndrome: Clinical pattern, cofactor effect and profile of molecular sensitization to plant-foods and pollens. Clin. Exp. Allergy 2012, 42, 1529–1539. [Google Scholar] [CrossRef]
- Mothes-Luksch, N.; Raith, M.; Stingl, G.; Focke-Tejkl, M.; Razzazi-Fazeli, E.; Zieglmayer, R.; Wöhrl, S.; Swoboda, I. Pru p 3, a marker allergen for lipid transfer protein sensitization also in Central Europe. Allergy 2017, 72, 1415–1418. [Google Scholar] [CrossRef] [Green Version]
- Skypala, I.J.; Bartra, J.; Ebo, D.G.; Antje Faber, M.; Fernández-Rivas, M.; Gomez, F.; Luengo, O.; Till, S.J.; Asero, R.; Barber, D.; et al. The diagnosis and management of allergic reactions in patients sensitized to non-specific lipid transfer proteins. Allergy 2021, 76, 2433–2446. [Google Scholar] [CrossRef] [PubMed]
- Azofra, J.; Berroa, F.; Gastaminza, G.; Saiz, N.; Gamboa, P.M.; Vela, C.; García, B.E.; Lizarza, S.; Echenagusia, M.A.; Joral, A.; et al. Lipid transfer protein syndrome in a non-mediterranean area. Int. Arch. Allergy Immunol. 2016, 169, 181–188. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to pectins and reduction of post-prandial glycaemic responses (ID 786), maintenance of normal blood cholesterol concentrations (ID 818) and increase in satiety leading to a reduction in energy intake (ID 4692) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2010, 8, 1747. [Google Scholar] [CrossRef]
- Fernández-Rivas, M.; González-Mancebo, E.; Rodríguez-Pérez, R.; Benito, C.; Sánchez-Monge, R.; Salcedo, G.; Alonso, M.D.; Rosado, A.; Tejedor, M.A.; Vila, C.; et al. Clinically relevant peach allergy is related to peach lipid transfer protein, Pru p 3, in the Spanish population. J. Allergy Clin. Immunol. 2003, 112, 789–795. [Google Scholar] [CrossRef]
- Toda, M.; Reese, G.; Gadermaier, G.; Schulten, V.; Lauer, I.; Egger, M.; Briza, P.; Randow, S.; Wolfheimer, S.; Kigongo, V.; et al. Protein unfolding strongly modulates the allergenicity and immunogenicity of Pru p 3, the major peach allergen. J. Allergy Clin. Immunol. 2011, 128, 1022–1030.e17. [Google Scholar] [CrossRef] [PubMed]
- Björkstén, F.; Halmepuro, L.; Hannuksela, M.; Lahti, A. Extraction and properties of apple allergens. Allergy 1980, 35, 671–677. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Kyhse-Andersen, J. Electroblotting of multiple gels: A simple apparatus without buffer tank for rapid transfer of proteins from polyacrylamide to nitrocellulose. J. Biochem. Biophys. Methods 1984, 10, 203–209. [Google Scholar] [CrossRef]
- Niu, L.; Zhang, H.; Wu, Z.; Wang, Y.; Liu, H.; Wu, X.; Wang, W. Modified TCA/acetone precipitation of plant proteins for proteomic analysis. PLoS ONE 2018, 13, e0202238. [Google Scholar] [CrossRef] [Green Version]
- Lević, L.; Tekić, M.; Djurić, M.; Kuljanin, T. CaCl2, CuSO4 and AlCl3 & NaHCO3 as possible pectin precipitants in sugar juice clarification. Int. J. Food Sci. Tech. 2007, 42, 609–614. [Google Scholar] [CrossRef]
- Vogel, L.; Holzhauser, T.; Vieths, S. Development of a biological assay to determine the allergenic potential of foods. J. Verbr. Lebensm. 2006, 1, 317–324. [Google Scholar] [CrossRef]
- Hartz, C.; Del San Miguel-Moncín, M.M.; Cisteró-Bahíma, A.; Fötisch, K.; Metzner, K.J.; Fortunato, D.; Lidholm, J.; Vieths, S.; Scheurer, S. Molecular characterisation of Lac s 1, the major allergen from lettuce (Lactuca sativa). Mol. Immunol. 2007, 44, 2820–2830. [Google Scholar] [CrossRef]
- Mortensen, A.; Aguilar, F.; Crebelli, R.; Di Domenico, A.; Dusemund, B.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Lambré, C.; et al. Re-evaluation of pectin (E 440i) and amidated pectin (E 440ii) as food additives. EFSA J. 2017, 15, e04866. [Google Scholar] [CrossRef]
- Polovic, N.; Blanusa, M.; Gavrovic-Jankulovic, M.; Atanaskovic-Markovic, M.; Burazer, L.; Jankov, R.; Cirkovic Velickovic, T. A matrix effect in pectin-rich fruits hampers digestion of allergen by pepsin in vivo and in vitro. Clin. Exp. Allergy 2007, 37, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Zeuner, B.; Thomsen, T.B.; Stringer, M.A.; Krogh, K.B.R.M.; Meyer, A.S.; Holck, J. Comparative characterization of aspergillus pectin lyases by discriminative substrate degradation profiling. Front. Bioeng. Biotechnol. 2020, 8, 873. [Google Scholar] [CrossRef] [PubMed]
- Yavuz-Düzgün, M.; Zeeb, B.; Dreher, J.; Özçelik, B.; Weiss, J. The impact of esterification degree and source of pectins on complex coacervation as a tool to mask the bitterness of potato protein isolates. Food Biophys. 2020, 15, 376–385. [Google Scholar] [CrossRef]
- Gawkowska, D.; Cybulska, J.; Zdunek, A. Structure-related gelling of pectins and linking with other natural compounds: A Review. Polymers 2018, 10, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaul, S.; Lüttkopf, D.; Kastner, B.; Vogel, L.; Höltz, G.; Vieths, S.; Hoffmann, A. Mediator release assays based on human or murine immunoglobulin E in allergen standardization. Clin. Exp. Allergy 2007, 37, 141–150. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steigerwald, H.; Blanco-Perez, F.; Albrecht, M.; Bender, C.; Wangorsch, A.; Endreß, H.-U.; Bunzel, M.; Mayorga, C.; Torres, M.J.; Scheurer, S.; et al. Does the Food Ingredient Pectin Provide a Risk for Patients Allergic to Non-Specific Lipid-Transfer Proteins? Foods 2022, 11, 13. https://doi.org/10.3390/foods11010013
Steigerwald H, Blanco-Perez F, Albrecht M, Bender C, Wangorsch A, Endreß H-U, Bunzel M, Mayorga C, Torres MJ, Scheurer S, et al. Does the Food Ingredient Pectin Provide a Risk for Patients Allergic to Non-Specific Lipid-Transfer Proteins? Foods. 2022; 11(1):13. https://doi.org/10.3390/foods11010013
Chicago/Turabian StyleSteigerwald, Hanna, Frank Blanco-Perez, Melanie Albrecht, Caroline Bender, Andrea Wangorsch, Hans-Ulrich Endreß, Mirko Bunzel, Cristobalina Mayorga, Maria José Torres, Stephan Scheurer, and et al. 2022. "Does the Food Ingredient Pectin Provide a Risk for Patients Allergic to Non-Specific Lipid-Transfer Proteins?" Foods 11, no. 1: 13. https://doi.org/10.3390/foods11010013
APA StyleSteigerwald, H., Blanco-Perez, F., Albrecht, M., Bender, C., Wangorsch, A., Endreß, H.-U., Bunzel, M., Mayorga, C., Torres, M. J., Scheurer, S., & Vieths, S. (2022). Does the Food Ingredient Pectin Provide a Risk for Patients Allergic to Non-Specific Lipid-Transfer Proteins? Foods, 11(1), 13. https://doi.org/10.3390/foods11010013