
Microbial Communities and Physiochemical Properties of Four Distinctive Traditionally Fermented Vegetables from North China and Their Influence on Quality and Safety

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Physicochemical Analysis
2.3. DNA Eextraction and Polymerase Chain Reaction (PCR) Amplification
2.4. Illumina MiSeq Sequencing
2.5. Sequencing Data Processing
2.6. Statistical Analysis
3. Results
3.1. Physicochemical Properties of the Four Traditionally Fermented Vegetables
3.2. Diversity Index Comparison of the Four Traditionally Fermented Vegetables
3.3. Bacterial Community Comparisons
3.4. Bacterial Profiles of the Four Traditionally Fermented Vegetables
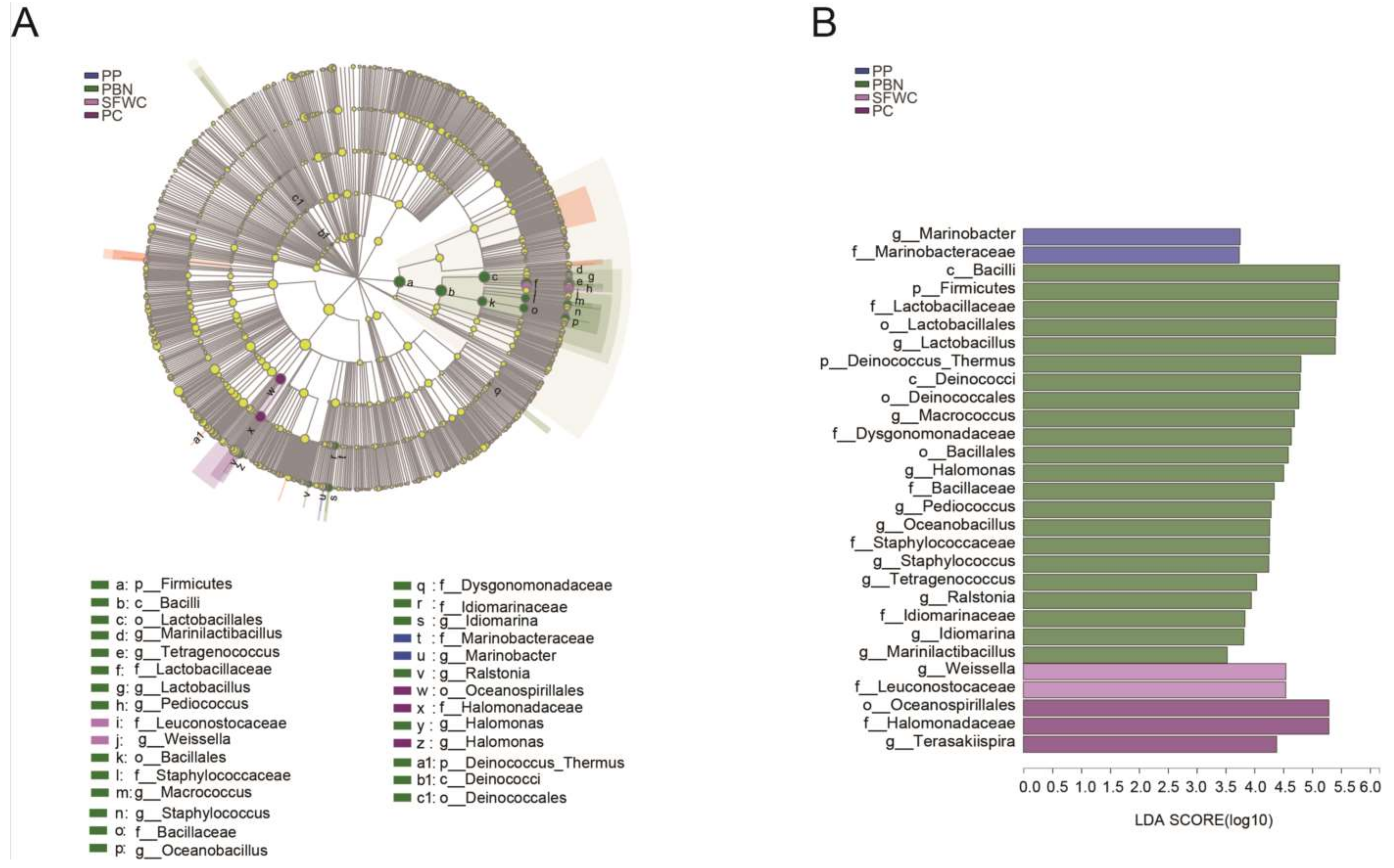
3.5. Differential Bacteria in the Four Traditionally Fermented Vegetables
3.6. Correlation of Bacterial Communities and the Physicochemical Indices of the Four Traditionally Fermented Vegetables
3.7. Predicted Functions of Metabolic
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, Z.; Peng, Z.; Huang, T.; Xiao, Y.; Li, J.; Xie, M.; Xiong, T. Comparison of bacterial diversity in traditionally homemade paocai and Chinese spicy cabbage. Food Microbiol. 2019, 83, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Zheng, W.; Huang, T.; Xiao, Y.; Xiong, T. Comparison of microbial communities and physiochemical characteristics of two traditionally fermented vegetables. Food Res. Int. 2019, 128, 108755. [Google Scholar] [CrossRef]
- Cao, J.; Yang, J.; Hou, Q.; Xu, H.; Zheng, Y.; Zhang, H.; Zhang, L. Assessment of bacterial profiles in aged, home-made Sichuan paocai brine with varying titratable acidity by PacBio SMRT sequencing technology. Food Control 2017, 78, 14–23. [Google Scholar] [CrossRef]
- Wang, L.; Pi, Z.; Liu, S.; Liu, Z.; Song, F. Targeted metabolome profiling by dual-probe microdialysis sampling and treatment using Gardenia jasminoides for rats with type 2 diabetes. Sci. Rep. 2017, 7, 10105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Zhang, X.; Liao, P.; Li, Z.; Li, W.; Li, X.; Wu, Y.; Pei, F. NMR spectroscopic-based metabonomic investigation on the acute biochemical effects induced by Ce(NO3)3 in rats. J. Inorg. Biochem. 2005, 99, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Chun, J.; Cha, C.J.; Park, W.S.; Jeon, C.O.; Bae, J.W. Bacterial community analysis during fermentation of ten representative kinds of kimchi with barcoded pyrosequencing. Food Microbiol. 2012, 30, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shao, Y. Effects of microbial diversity on nitrite concentration in pao cai, a naturally fermented cabbage product from China. Food Microbiol. 2018, 72, 185–192. [Google Scholar] [CrossRef]
- Chao, S.H.; Wu, R.J.; Watanabe, K.; Tsai, Y.C. Diversity of lactic acid bacteria in suan-tsai and fu-tsai, traditional fermented mustard products of Taiwan. Int. J. Food Microbiol. 2009, 135, 203–210. [Google Scholar] [CrossRef]
- Nguyen, D.T.L.; Van Hoorde, K.; Cnockaert, M.; De Brandt, E.; Aerts, M.; Binh Thanh, L.; Vandamme, P. A description of the lactic acid bacteria microbiota associated with the production of traditional fermented vegetables in Vietnam. Int. J. Food Microbiol. 2013, 163, 19–27. [Google Scholar] [CrossRef]
- Liu, Z.; Li, J.; Wei, B.; Huang, T.; Xiao, Y.; Peng, Z.; Xie, M.; Xiong, T. Bacterial community and composition in Jiang-shui and Suan-cai revealed by high-throughput sequencing of 16S rRNA. Int. J. Food Microbiol. 2019, 306, 108271. [Google Scholar] [CrossRef]
- Lee, M.; Song, J.H.; Jung, M.Y.; Lee, S.H.; Chang, J.Y. Large-scale targeted metagenomics analysis of bacterial ecological changes in 88 kimchi samples during fermentation. Food Microbiol. 2017, 66, 173–183. [Google Scholar] [CrossRef]
- Xiong, T.; Guan, Q.; Song, S.; Hao, M.; Xie, M. Dynamic changes of lactic acid bacteria flora during Chinese sauerkraut fermentation. Food Control 2012, 26, 178–181. [Google Scholar] [CrossRef]
- Anal, A.K.; Perpetuini, G.; Petchkongkaew, A.; Tan, R.; Waché, Y. Food safety risks in traditional fermented food from South-East Asia. Food Control 2019, 109, 106922. [Google Scholar] [CrossRef]
- Nout, M.; Bakshi, D.; Sarkar, P.K. Microbiological safety of kinema, a fermented soya bean food. Food Control 1998, 9, 357–362. [Google Scholar] [CrossRef]
- Hernández-Carrión, M.; Sanz, T.; Hernando, I.; Llorca, E.; Fiszman, S.M.; Quiles, A. New formulations of functional white sauces enriched with red sweet pepper: A rheological, microstructural and sensory study. Eur. Food Res. Technol. 2015, 240, 1187–1202. [Google Scholar] [CrossRef]
- Beato, V.M.; Sánchez, A.H.; De Castro, A.; Montaño, A. Effect of Processing and Storage Time on the Contents of Organosulfur Compounds in Pickled Blanched Garlic. J. Agr. Food Chem. 2012, 60, 3485. [Google Scholar] [CrossRef]
- He, Z.; Chen, H.; Wang, X.; Lin, X.; Ji, C.; Li, S.; Liang, H. Effects of different temperatures on bacterial diversity and volatile flavor compounds during the fermentation of suancai, a traditional fermented vegetable food from northeastern China. LWT-Food Sci. Technol. 2020, 118, 108773. [Google Scholar] [CrossRef]
- Wan, H.; Liu, T.; Su, C.; Ji, X.; Wang, L.; Zhao, Y.; Wang, Z. Evaluation of bacterial and fungal communities during the fermentation of Baixi sufu, a traditional spicy fermented bean curd. J. Sci. Food Agric. 2020, 100, 1448–1457. [Google Scholar] [CrossRef]
- Feng, K.; Sun, Q.; Deng, W.Q.; Zhang, S.; Qi-Sheng, Z.; Fan, Z.Y.; You, J.G.; Heng, L.I. A HPLC Method for Determination of Seven Kinds of Organic Acids in Broad Bean Paste With Different Fermentation Year. Food Fermen. Sci. Technol. 2019, 55, 128–141. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Jia, G. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Wang, Q. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Liu, G.; Huang, C.; Shen, L.; Li, C.; Liu, Y.; Liu, S.; Hu, B.; Chen, H. The bacterial diversity of ripened Guang’yuan Suancai and in vitro evaluation of potential probiotic lactic acid bacteria isolated from Suancai. LWT-Food Sci. Technol. 2017, 85, 175–180. [Google Scholar] [CrossRef]
- Liu, X.; Kuda, T.; Takahashi, H.; Kimura, B. Bacterial and fungal microbiota of spontaneously fermented Chinese products, Rubing milk cake and Yan-cai vegetable pickles. Food Microbiol. 2018, 72, 106. [Google Scholar] [CrossRef]
- Jeong, S.H.; Lee, H.J.; Jung, J.Y.; Lee, S.H.; Seo, H.Y.; Park, W.S.; Jeon, C.O. Effects of red pepper powder on microbial communities and metabolites during kimchi fermentation. Int. J. Food Microbiol. 2013, 160, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Jinhee, C.; Dongyun, L.; Changnam, Y.; Jongin, J.; Jeongho, K.; Hongui, H. Microbial population dynamics of kimchi, a fermented cabbage product. FEMS Microbiol. Lett. 2010, 257, 262–267. [Google Scholar]
- Lee, J.S.; Heo, G.Y.; Lee, J.W.; Oh, Y.J.; Ahn, J.S. Analysis of kimchi microflora using denaturing gradient gel electrophoresis. Int. J. Food Microbiol. 2005, 102, 143–150. [Google Scholar] [CrossRef]
- Zhou, Q.; Zang, S.; Zhao, Z.; Li, X. Dynamic changes of bacterial communities and nitrite character during northeastern Chinese sauerkraut fermentation. Food Sci. Biotechnol. 2018, 27, 79–85. [Google Scholar] [CrossRef]
- Namwong, S.; Tanasupawat, S.; Lee, K.C.; Lee, J.S. Oceanobacillus kapialis sp. nov., from fermented shrimp paste in Thailand. Int. J. Syst. Evol. Micr. 2009, 59, 2254–2259. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Oh, T.K.; Kim, W.; Yoon, J.H. Oceanobacillus locisalsi sp. nov. isolated from a marine solar saltern. Int. J. Syst. Evol. Micr. 2010, 60, 2758–2762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, B.; Saleem, M.; Naseer, G.; Fida, A. Significance of Staphylococcus hemolyticus in nosocomial associated Infections. Pak. J. Med. Sci. 2014, 4, 119–125. [Google Scholar]
- Pérez-Cataluña, A.; Salas-Massó, N.; Figueras, M.J. Arcobacter canalis sp. nov., isolated from a water canal contaminated with urban sewage. Int. J. Syst. Evol. Micr. 2018, 68, 1258–1264. [Google Scholar] [CrossRef]
- Jung, H.M.; Liu, Q.M.; Kim, J.K.; Lee, S.-T. Lactobacillus ginsenosidimutans sp. nov., isolated from kimchi with the ability to transform ginsenosides. Antonie Van Leeuwenhoek 2013, 103, 867–876. [Google Scholar] [CrossRef]
- Entani, E.; Masai, H.; Suzuki, K.I. Lactobacillus acetotolerans, a New Species from Fermented Vinegar Broth. Int. J. Syst. Bacteriol. 1986, 36, 544–549. [Google Scholar] [CrossRef]
- Yang, J.; Cao, J.; Xu, H.; Hou, Q.; Yu, Z.; Zhang, H.; Sun, Z. Bacterial diversity and community structure in Chongqing radish paocai brines revealed using PacBio single-molecule real-time sequencing technology. J. Sci. Food Agr. 2018, 98, 3234–3245. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Wood, B.J.B. Lactic Acid Bacteria: Biodiversity and Taxonomy; Wiley: Hoboken, NJ, USA, 2014; pp. 507–508. [Google Scholar]
- Tanasupawat, S.; Shida, O.; Okada, S.; Komagata, K. Lactobacillus acidipiscis sp. nov. and Weissella thailandensis sp. nov., isolated from fermented fish in Thailand. Int. J. Syst. Evol. Micr. 2000, 50 Pt 4, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, H.; Nagai, Y.; Miyoshi, K.; Kawamoto, T.; Watanabe, T. 3P-068 Pancreatic lipase inhibitory activity of lactic acid bacteria isolated from pickles, Lactobacillus acidipiscis TK12408. In Proceedings of the Congress of the Society for Biotechnology, Sapporo, Japan, 26–28 June 2013. [Google Scholar]
- Kim, M.; Chunt, J. Bacterial community structure in kimchi, a Korean fermented vegetable food, as revealed by 16S rRNA gene analysis. Int. J. Food Microbiol. 2005, 103, 91–96. [Google Scholar] [CrossRef]
- Park, S.E.; Seo, S.H.; Kim, E.J.; Na, C.S.; Son, H.S. Effects of different fermentation temperatures on metabolites of Kimchi. Food Biosci. 2018, 23, 100–106. [Google Scholar] [CrossRef]
- Li, H.; Li, Z.; Qu, J.; Wang, J. Bacterial diversity in traditional Jiaozi and sourdough revealed by high-throughput sequencing of 16S rRNA amplicons. LWT-Food Sci. Technol. 2017, 81, 319–325. [Google Scholar] [CrossRef]
- Li, Q.; Kang, J.; Ma, Z.; Li, X.; Liu, L.; Hu, X. Microbial succession and metabolite changes during traditional serofluid dish fermentation. LWT-Food Sci. Technol. 2017, 84, 771–779. [Google Scholar] [CrossRef]
- Collar, C.; Mascarós, A.F.; Barber, C.B.D. Amino Acid Metabolism by Yeasts and Lactic Acid Bacteria during Bread Dough Fermentation. J. Food Sci. 1992, 57, 1423–1427. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample_Name | Seq_Num | Average Length (bp) | OTU | Shannon | Simpson | Ace | Chao1 | Coverage | |
|---|---|---|---|---|---|---|---|---|---|
| PP | PP_83 | 50,570 | 428 | 109 | 1.472 | 0.377 | 216.499 | 154.556 | 0.999 |
| PP_254 | 48,552 | 422 | 322 | 3.241 | 0.092 | 362.423 | 377.313 | 0.998 | |
| PP_263 | 56,438 | 406 | 83 | 0.324 | 0.908 | 160.771 | 124.333 | 0.999 | |
| PP_318 | 62,935 | 415 | 59 | 1.019 | 0.448 | 159.878 | 102.875 | 0.999 | |
| PBN | PBN_81 | 54,631 | 424 | 452 | 3.635 | 0.061 | 489.033 | 497.643 | 0.998 |
| PBN_147 | 51,414 | 427 | 245 | 2.762 | 0.133 | 416.913 | 361.000 | 0.997 | |
| PBN_198 | 61,650 | 427 | 535 | 2.774 | 0.230 | 733.741 | 701.693 | 0.994 | |
| PBN_260 | 48,502 | 428 | 141 | 2.090 | 0.171 | 523.611 | 304.235 | 0.998 | |
| SFWC | SFWC_118 | 43,810 | 412 | 158 | 1.395 | 0.440 | 219.884 | 223.900 | 0.998 |
| SFWC_120 | 46,411 | 409 | 251 | 1.171 | 0.547 | 476.835 | 360.306 | 0.997 | |
| SFWC_238 | 46,848 | 414 | 287 | 1.465 | 0.451 | 625.914 | 464.349 | 0.996 | |
| SFWC_239 | 36,547 | 410 | 197 | 0.865 | 0.658 | 268.803 | 305.043 | 0.998 | |
| SFWC_257 | 61,458 | 407 | 94 | 0.416 | 0.826 | 262.913 | 154.882 | 0.999 | |
| PC | PC_132 | 53,280 | 421 | 86 | 1.825 | 0.217 | 140.644 | 123.800 | 0.999 |
| PC_133 | 50,564 | 424 | 142 | 1.616 | 0.343 | 198.557 | 180.607 | 0.999 | |
| PC_192 | 67,831 | 429 | 34 | 0.693 | 0.663 | 44.481 | 49.000 | 1.000 | |
| PC_237 | 63,784 | 416 | 419 | 2.644 | 0.227 | 804.149 | 662.019 | 0.995 | |
| PC_269 | 59,771 | 406 | 119 | 0.249 | 0.931 | 333.021 | 202.182 | 0.998 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, T.; Xie, X.; Wu, L.; Li, L.; Li, H.; Xi, Y.; Feng, Y.; Xue, L.; Chen, M.; Chen, X.; et al. Microbial Communities and Physiochemical Properties of Four Distinctive Traditionally Fermented Vegetables from North China and Their Influence on Quality and Safety. Foods 2022, 11, 21. https://doi.org/10.3390/foods11010021
Liang T, Xie X, Wu L, Li L, Li H, Xi Y, Feng Y, Xue L, Chen M, Chen X, et al. Microbial Communities and Physiochemical Properties of Four Distinctive Traditionally Fermented Vegetables from North China and Their Influence on Quality and Safety. Foods. 2022; 11(1):21. https://doi.org/10.3390/foods11010021
Chicago/Turabian StyleLiang, Tingting, Xinqiang Xie, Lei Wu, Longyan Li, Haixin Li, Yu Xi, Ying Feng, Liang Xue, Moutong Chen, Xuefeng Chen, and et al. 2022. "Microbial Communities and Physiochemical Properties of Four Distinctive Traditionally Fermented Vegetables from North China and Their Influence on Quality and Safety" Foods 11, no. 1: 21. https://doi.org/10.3390/foods11010021
APA StyleLiang, T., Xie, X., Wu, L., Li, L., Li, H., Xi, Y., Feng, Y., Xue, L., Chen, M., Chen, X., Zhang, J., Ding, Y., & Wu, Q. (2022). Microbial Communities and Physiochemical Properties of Four Distinctive Traditionally Fermented Vegetables from North China and Their Influence on Quality and Safety. Foods, 11(1), 21. https://doi.org/10.3390/foods11010021