
Oral Administration of Ethanolic Extract of Shrimp Shells-Loaded Liposome Protects against Aβ-Induced Memory Impairment in Rats

 ,
,  , ,
, , 
Abstract
:
1. Introduction
2. Materials and Methods
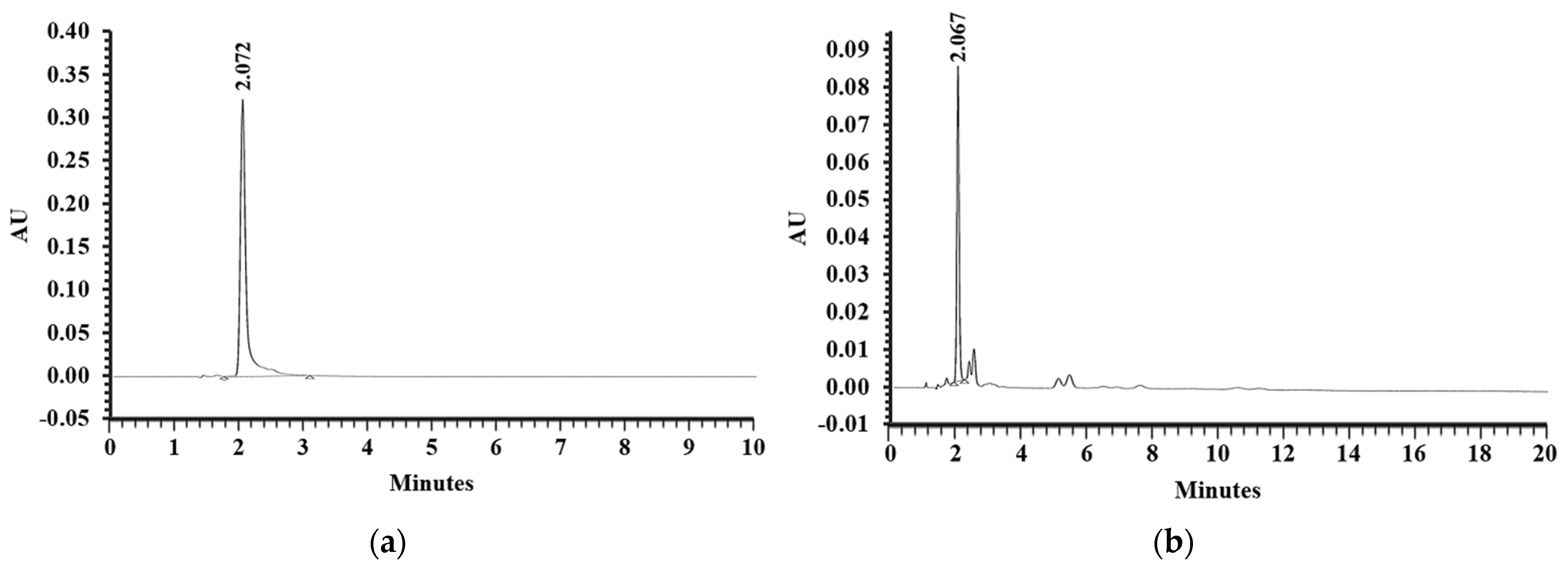
2.1. Extraction of EESS and RP-HPLC Analysis of AST Content in the EESS
2.2. Preparation of EESS-Loaded Liposome
2.3. Characterization of EESS-Loaded Liposome
2.3.1. Encapsulation Efficiency (EE)
2.3.2. Particle Size, Polydispersity Index and Zeta Potential
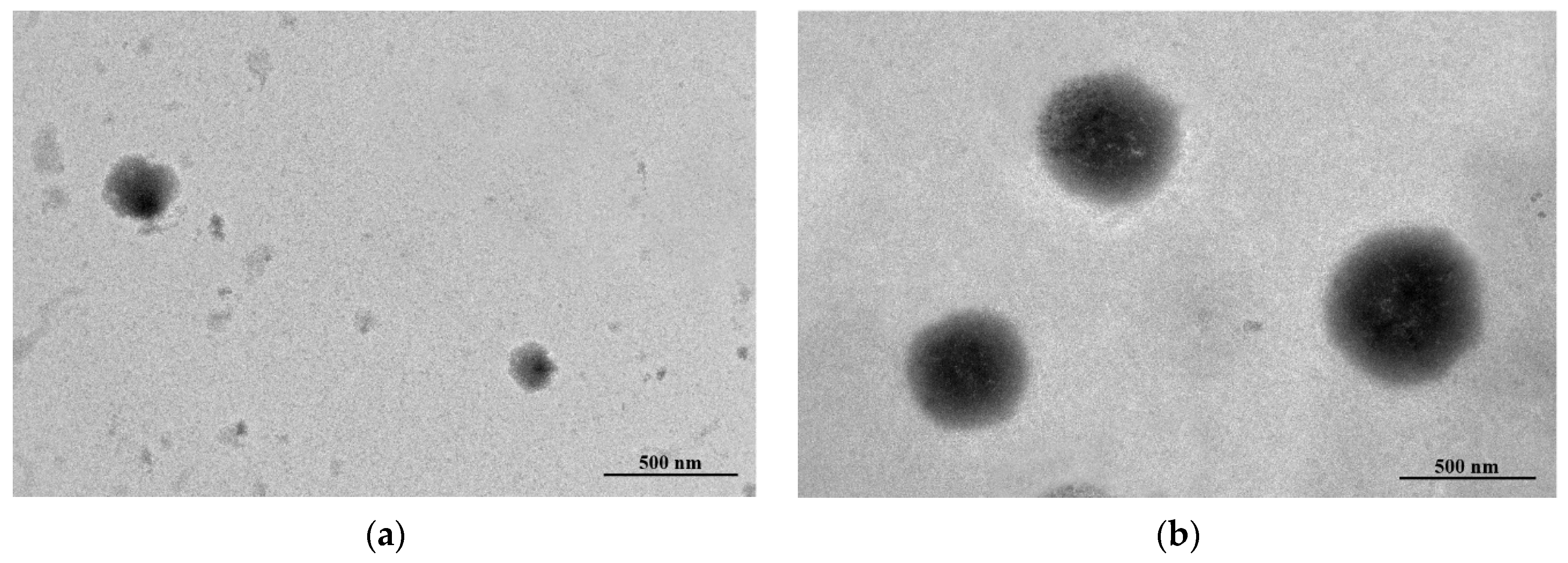
2.3.3. Morphology Study
2.4. Animals
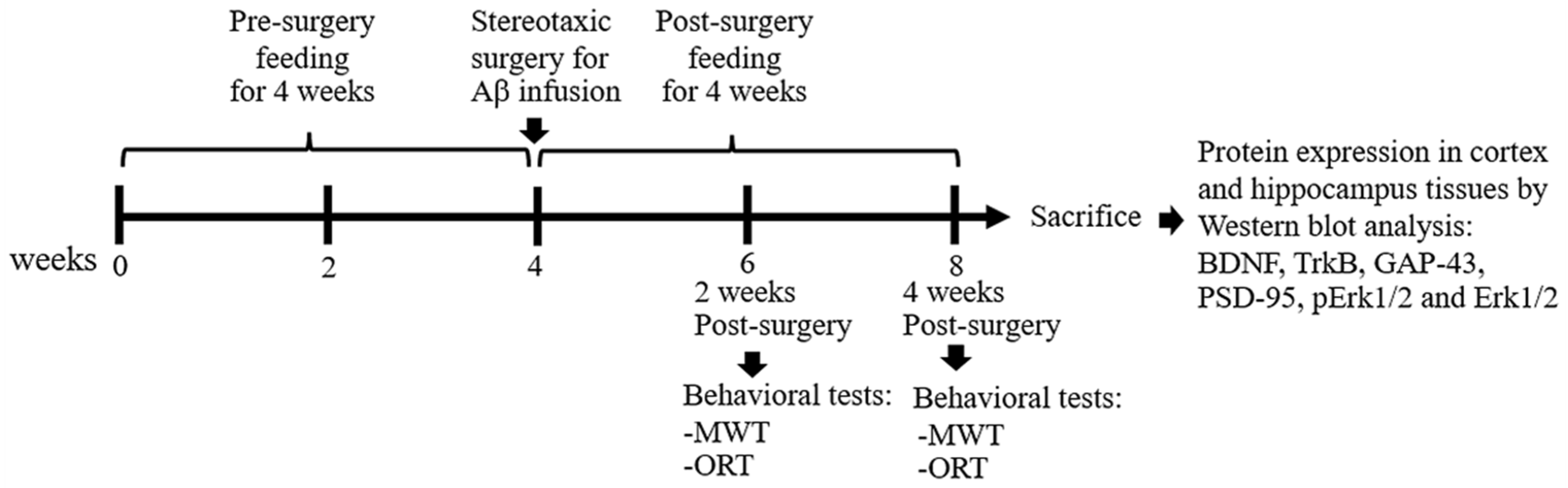
2.5. Experimental Design
2.6. Morris Water MAZE Test
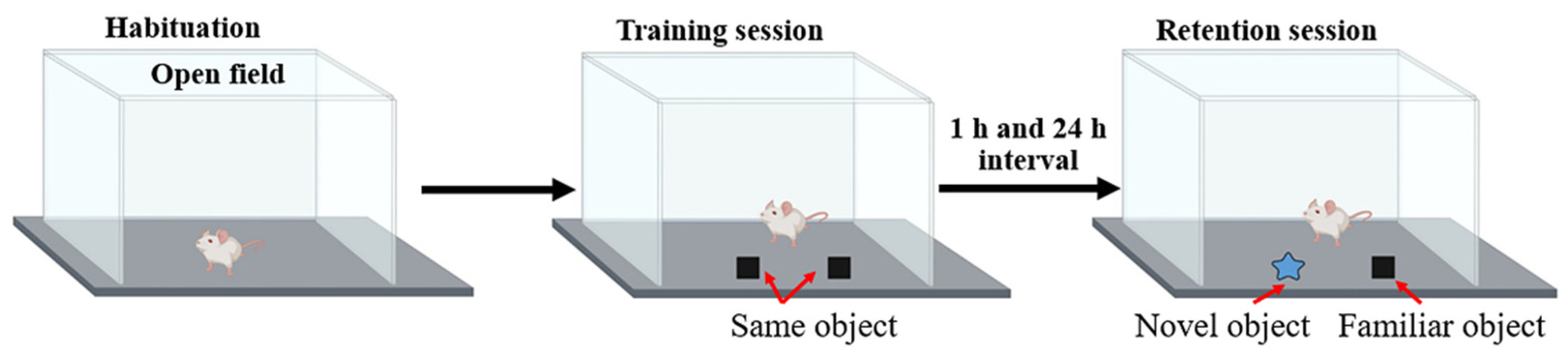
2.7. Object Recognition Test
2.8. Western Blot Analysis
2.9. Data Analysis
3. Results
3.1. AST Content in Ethanolic Extract from Shrimp Shells
3.2. Encapsulation Efficiency (EE), Particle Size, Zeta-Potential and Transmission Electron Microscopy (TEM) Study of EESS-Loaded Liposomes
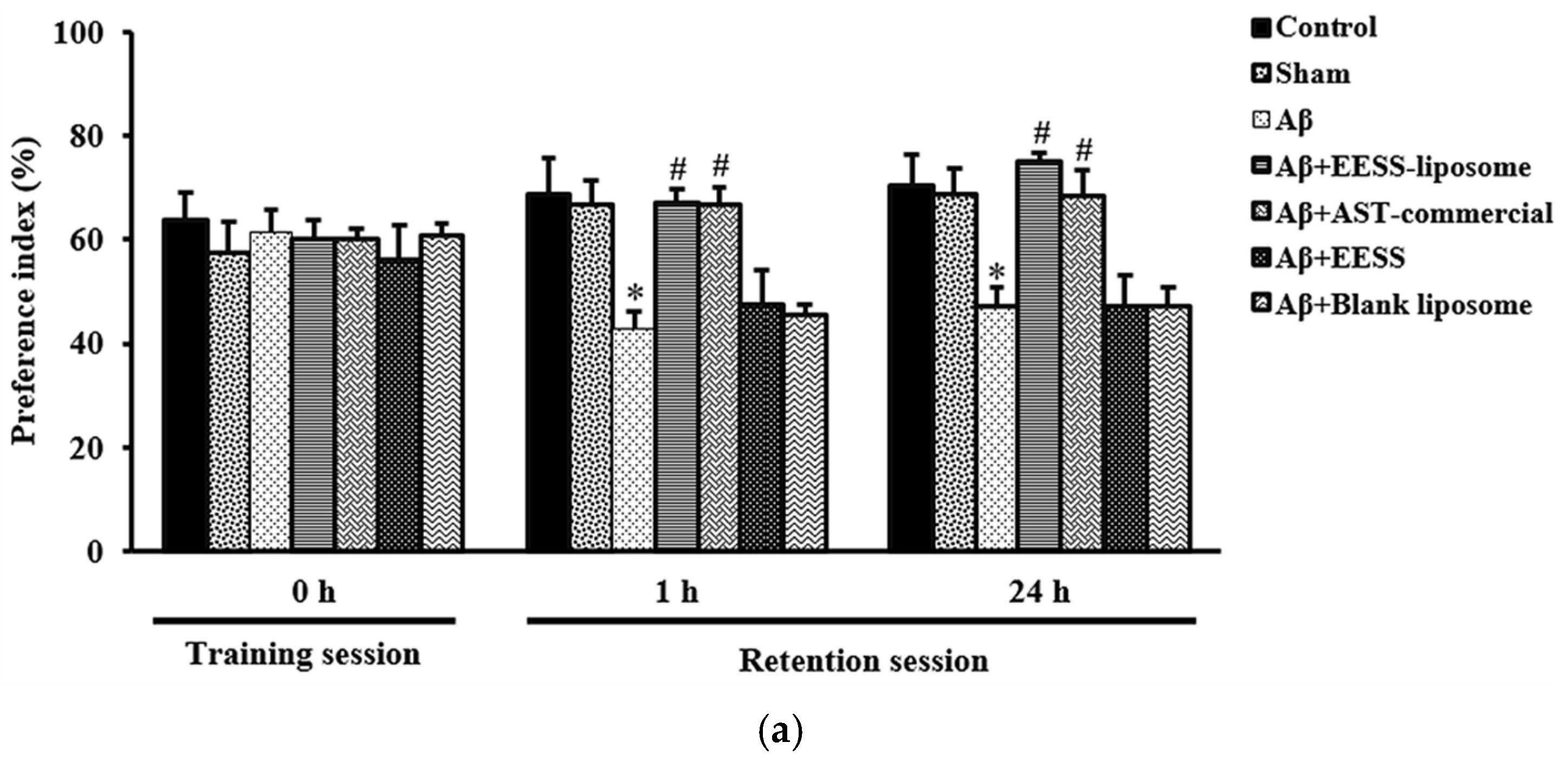
3.3. EESS-Loaded Liposome Improved Memory and Cognitive Ability in the Memory-Related Behavioral Tests
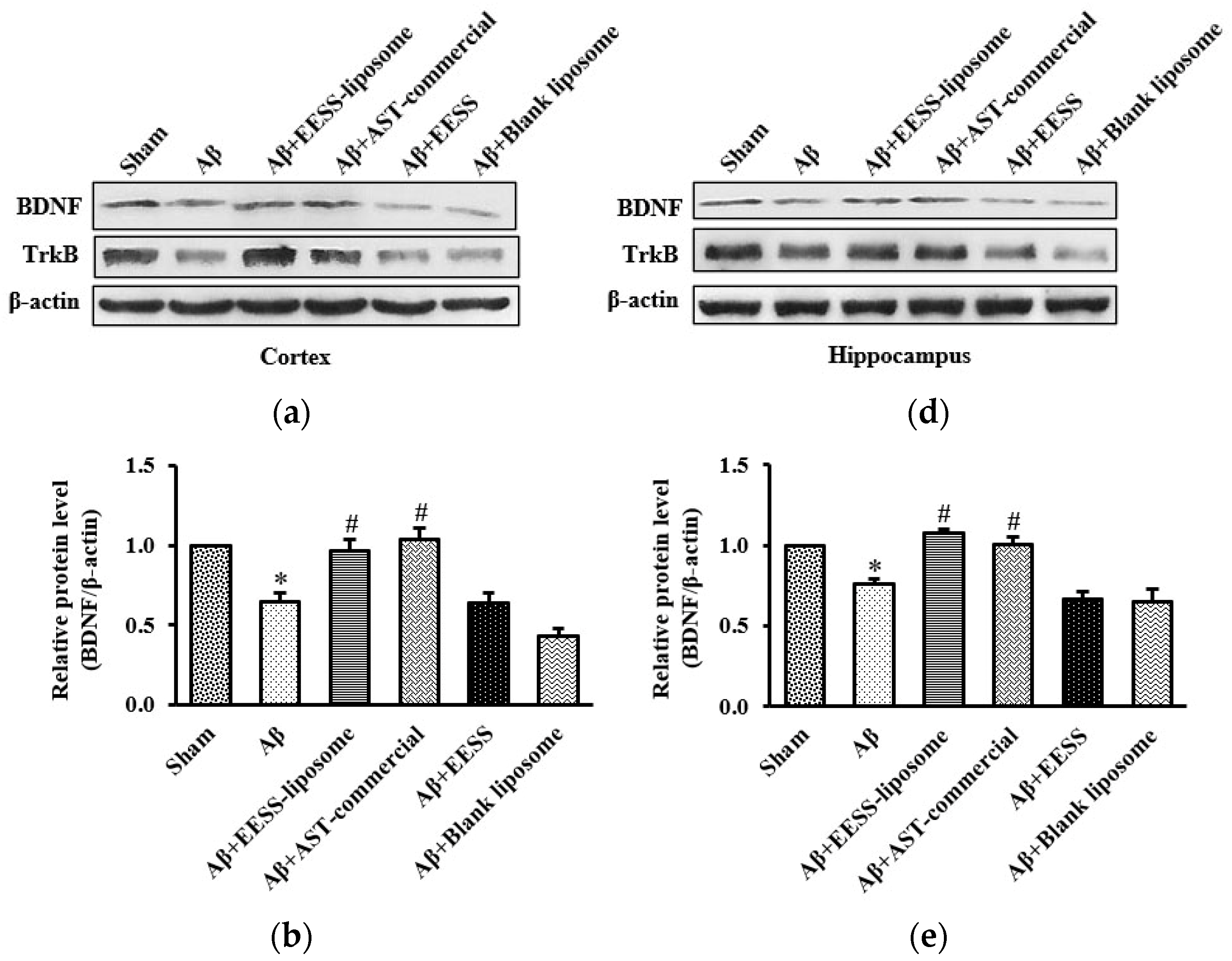
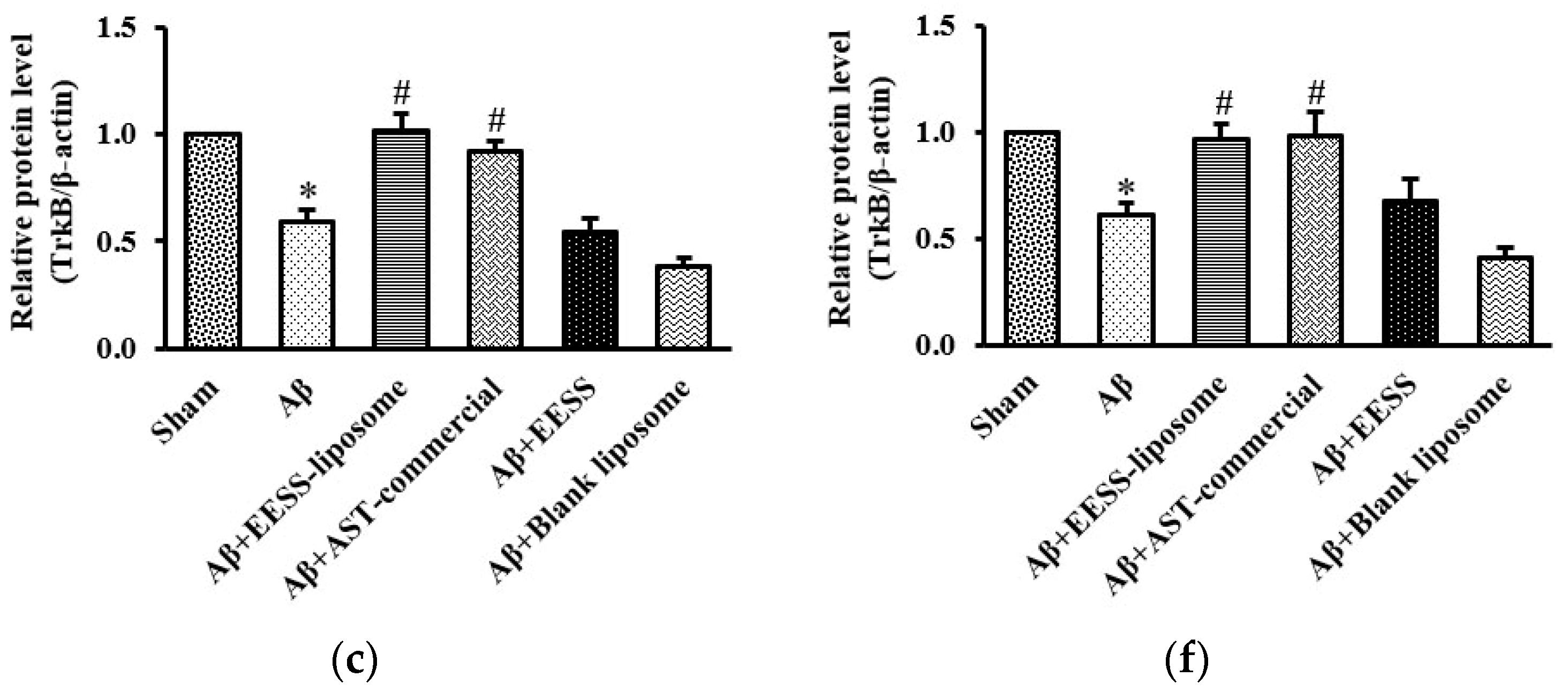
3.4. EESS-Loaded Liposome Increased the Levels of BDNF and TrkB in AD Rats
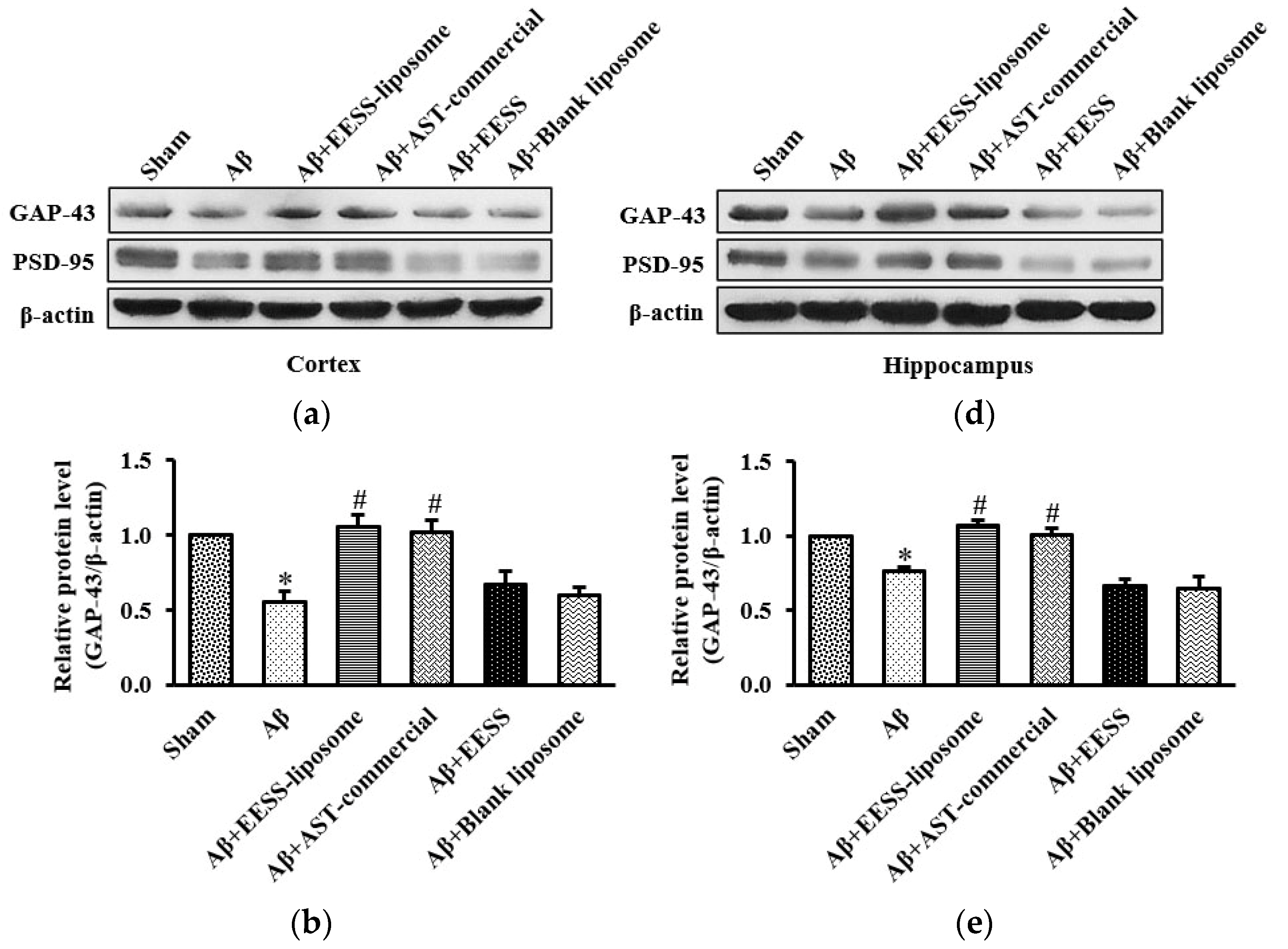
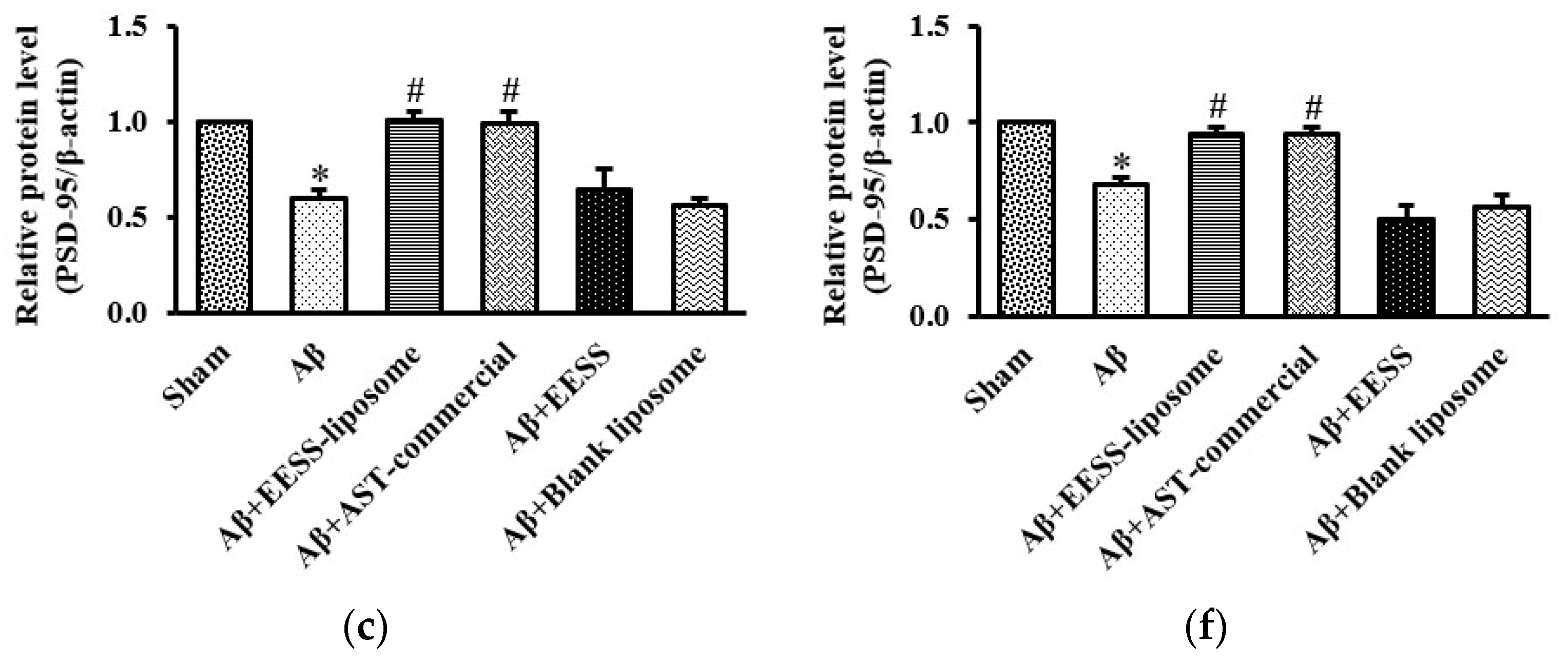
3.5. EESS-Loaded Liposome Ameliorated Aβ-Suppressed GAP-43 and PSD-95 Synaptic Plasticity Proteins
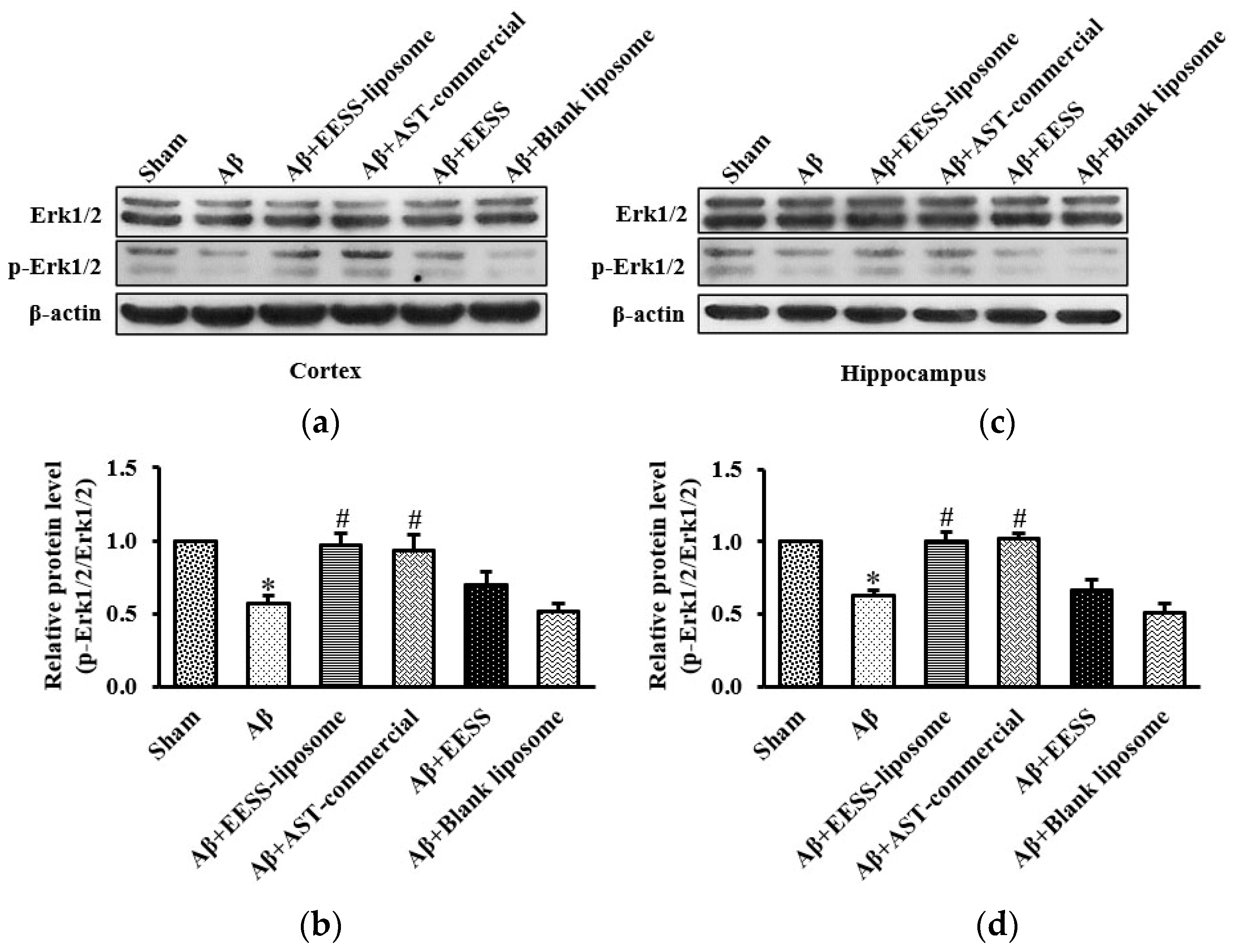
3.6. EESS-Loaded Liposome Suppressed Aβ-Induced Cognition and Synaptic Plasticity Deficits via Erk Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s Disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Sciaccaluga, M.; Megaro, A.; Bellomo, G.; Ruffolo, G.; Romoli, M.; Palma, E.; Costa, C. An Unbalanced Synaptic Transmission: Cause or Consequence of the Amyloid Oligomers Neurotoxicity? Int. J. Mol. Sci. 2021, 22, 5991. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Huang, Z.; Du, Y.; Fu, M.; Han, H.; Wang, Y.; Dong, Z. Capsaicin Attenuates Amyloid-β-Induced Synapse Loss and Cognitive Impairments in Mice. J. Alzheimer’s Dis. 2017, 59, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Numakawa, T.; Odaka, H. Brain-Derived Neurotrophic Factor Signaling in the Pathophysiology of Alzheimer’s Disease: Beneficial Effects of Flavonoids for Neuroprotection. Int. J. Mol. Sci. 2021, 22, 5719. [Google Scholar] [CrossRef]
- Pang, J.; Hou, J.; Zhou, Z.; Ren, M.; Mo, Y.; Yang, G.; Qu, Z.; Hu, Y. Safflower Yellow Improves Synaptic Plasticity in APP/PS1 Mice by Regulating Microglia Activation Phenotypes and BDNF/TrkB/ERK Signaling Pathway. NeuroMolecular Med. 2020, 22, 341–358. [Google Scholar] [CrossRef]
- Xiyang, Y.B.; Liu, R.; Wang, X.Y.; Li, S.; Zhao, Y.; Lu, B.T.; Xiao, Z.C.; Zhang, L.F.; Wang, T.H.; Zhang, J. COX5A Plays a Vital Role in Memory Impairment Associated With Brain Aging via the BDNF/ERK1/2 Signaling Pathway. Front. Aging Neurosci. 2020, 12, 215. [Google Scholar] [CrossRef]
- Holahan, M.R. A Shift from a Pivotal to Supporting Role for the Growth-Associated Protein (GAP-43) in the Coordination of Axonal Structural and Functional Plasticity. Front. Cell. Neurosci. 2017, 11, 266. [Google Scholar] [CrossRef]
- Coley, A.A.; Gao, W.J. PSD95: A Synaptic Protein Implicated in Schizophrenia or Autism? Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 82, 187–194. [Google Scholar] [CrossRef]
- Jerónimo-Santos, A.; Vaz, S.H.; Parreira, S.; Rapaz-Lérias, S.; Caetano, A.P.; Scherrer, V.B.; Castrén, E.; Valente, C.A.; Blum, D.; Sebastião, A.M.; et al. Dysregulation of TrkB Receptors and BDNF Function by Amyloid-β Peptide Is Mediated by Calpain. Cereb. Cortex 2015, 25, 3107–3121. [Google Scholar] [CrossRef]
- Zhang, L.; Fang, Y.; Xu, Y.; Lian, Y.; Xie, N.; Wu, T.; Zhang, H.; Sun, L.; Zhang, R.; Wang, Z. Curcumin Improves Amyloid β-Peptide (1-42) Induced Spatial Memory Deficits through BDNF-ERK Signaling Pathway. PLoS ONE 2015, 10, e0131525. [Google Scholar] [CrossRef] [Green Version]
- Ayyalasomayajula, N.; Bandaru, M.; Dixit, P.K.; Ajumeera, R.; Chetty, C.S.; Challa, S. Inactivation of GAP-43 Due to the Depletion of Cellular Calcium by the Pb and Amyloid Peptide Induced Toxicity: An in Vitro Approach. Chem. -Biol. Interact. 2020, 316, 108927. [Google Scholar] [CrossRef] [PubMed]
- Sheng, C.; Xu, P.; Zhou, K.; Deng, D.; Zhang, C.; Wang, Z. Icariin Attenuates Synaptic and Cognitive Deficits in an Aβ1-42-Induced Rat Model of Alzheimer’s Disease. BioMed Res. Int. 2017, 2017, 7464872. [Google Scholar] [CrossRef] [PubMed]
- Gulzar, S.; Raju, N.; Nagarajarao, R.C.; Benjakul, S. Oil and Pigments from Shrimp Processing By-Products: Extraction, Composition, Bioactivities and Its Application—A Review. Trends Food Sci. Technol. 2020, 100, 307–319. [Google Scholar] [CrossRef]
- Gul, K.; Tak, A.; Singh, A.K.; Singh, P.; Yousuf, B.; Wani, A.A. Chemistry, Encapsulation, and Health Benefits of β-Carotene—A Review. Cogent Food Agric. 2015, 1, 1018696. [Google Scholar] [CrossRef]
- Grimmig, B.; Hudson, C.; Moss, L.; Peters, M.; Subbarayan, M.; Weeber, E.J.; Bickford, P.C. Astaxanthin Supplementation Modulates Cognitive Function and Synaptic Plasticity in Young and Aged Mice. GeroScience 2019, 41, 77–87. [Google Scholar] [CrossRef]
- Jannel, S.; Caro, Y.; Bermudes, M.; Petit, T. Novel Insights into the Biotechnological Production of Haematococcus Pluvialis-Derived Astaxanthin: Advances and Key Challenges to Allow Its Industrial Use as Novel Food Ingredient. J. Mar. Sci. Eng. 2020, 8, 789. [Google Scholar] [CrossRef]
- Soukoulis, C.; Bohn, T. A Comprehensive Overview on the Micro- and Nano-Technological Encapsulation Advances for Enhancing the Chemical Stability and Bioavailability of Carotenoids. Crit. Rev. Food Sci. Nutr. 2018, 58, 1–36. [Google Scholar] [CrossRef]
- Gomez, A.G.; Syed, S.; Marshall, K.; Hosseinidoust, Z. Liposomal Nanovesicles for Efficient Encapsulation of Staphylococcal Antibiotics. ACS Omega 2019, 4, 10866–10876. [Google Scholar] [CrossRef]
- Zylberberg, C.; Matosevic, S. Pharmaceutical Liposomal Drug Delivery: A Review of New Delivery Systems and a Look at the Regulatory Landscape. Drug Deliv. 2016, 23, 3319–3329. [Google Scholar] [CrossRef]
- Pintea, A.; Diehl, H.A.; Momeu, C.; Aberle, L.; Socaciu, C. Incorporation of Carotenoid Esters into Liposomes. Biophys. Chem. 2005, 118, 7–14. [Google Scholar] [CrossRef]
- Gulzar, S.; Benjakul, S. Nanoliposome Powder Containing Shrimp Oil Increases Free Flowing Behavior and Storage Stability. Eur. J. Lipid Sci. Technol. 2020, 122, 2000049. [Google Scholar] [CrossRef]
- Chotphruethipong, L.; Battino, M.; Benjakul, S. Effect of Stabilizing Agents on Characteristics, Antioxidant Activities and Stability of Liposome Loaded with Hydrolyzed Collagen from Defatted Asian Sea Bass Skin. Food Chem. 2020, 328, 127127. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Paulson, A.T.; Gill, T.A. Encapsulation of Bioactive Salmon Protein Hydrolysates with Chitosan-Coated Liposomes. J. Funct. Foods 2015, 19, 733–743. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Elsevier: Amsterdam, The Netherlands, 1986. [Google Scholar]
- Morris, R. Developments of a Water-Maze Procedure for Studying Spatial Learning in the Rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Barker, G.R.I.; Warburton, E.C. Object-in-Place Associative Recognition Memory Depends on Glutamate Receptor Neurotransmission within Two Defined Hippocampal-Cortical Circuits: A Critical Role for AMPA and NMDA Receptors in the Hippocampus, Perirhinal, and Prefrontal Cortices. Cereb. Cortex 2015, 25, 472–481. [Google Scholar] [CrossRef]
- Taksima, T.; Chonpathompikunlert, P.; Sroyraya, M.; Hutamekalin, P.; Limpawattana, M.; Klaypradit, W. Effects of Astaxanthin from Shrimp Shell on Oxidative Stress and Behavior in Animal Model of Alzheimer’s Disease. Mar. Drugs 2019, 17, 628. [Google Scholar] [CrossRef]
- Dong, Y.; Hong, W.; Tang, Z.; Gao, Y.; Wu, X.; Liu, H. Sevoflurane Leads to Learning and Memory Dysfunction via Breaking the Balance of TPA/PAI. Neurochem. Int. 2020, 139, 104789. [Google Scholar] [CrossRef]
- Holahan, M. GAP-43 in Synaptic Plasticity: Molecular Perspectives. Res. Rep. Biochem. 2015, 5, 137–146. [Google Scholar] [CrossRef]
- Kabir, M.T.; Uddin, M.S.; Jeandet, P.; Emran, T.B.; Mitra, S.; Albadrani, G.M.; Sayed, A.A.; Abdel-Daim, M.M.; Simal-Gandara, J. Anti-Alzheimer’s Molecules Derived from Marine Life: Understanding Molecular Mechanisms and Therapeutic Potential. Mar. Drugs 2021, 19, 251. [Google Scholar] [CrossRef]
- Su, F.; Huang, B.; Liu, J. The Carotenoids of Shrimps (Decapoda: Caridea and Dendrobranchiata) Cultured in China. J. Crustac. Biol. 2018, 38, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Andar, A.U.; Hood, R.R.; Vreeland, W.N.; Devoe, D.L.; Swaan, P.W. Microfluidic Preparation of Liposomes to Determine Particle Size Influence on Cellular Uptake Mechanisms. Pharm. Res. 2014, 31, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.M.; Hyun, S.S.; Yun, P.; Lee, C.H.; Byun, S.Y. Identification of an Emulsifier and Conditions for Preparing Stable Nanoemulsions Containing the Antioxidant Astaxanthin. Int. J. Cosmet. Sci. 2012, 34, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Chotphruethipong, L.; Hutamekalin, P.; Sukketsiri, W.; Benjakul, S. Effects of Sonication and Ultrasound on Properties and Bioactivities of Liposomes Loaded with Hydrolyzed Collagen from Defatted Sea Bass Skin Conjugated with Epigallocatechin Gallate. J. Food Biochem. 2021, 45, e13809. [Google Scholar] [CrossRef] [PubMed]
- Sangsuriyawong, A.; Limpawattana, M.; Siriwan, D.; Klaypradit, W. Properties and Bioavailability Assessment of Shrimp Astaxanthin Loaded Liposomes. Food Sci. Biotechnol. 2019, 28, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Kinney, J.W.; Bemiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a Central Mechanism in Alzheimer’s Disease. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2018, 4, 575–590. [Google Scholar] [CrossRef]
- Reshetnikov, V.V.; Kisaretova, P.E.; Ershov, N.I.; Shulyupova, A.S.; Oshchepkov, D.Y.; Klimova, N.V.; Ivanchihina, A.V.; Merkulova, T.I.; Bondar, N.P. Genes Associated with Cognitive Performance in the Morris Water Maze: An RNA-Seq Study. Sci. Rep. 2020, 10, 22078. [Google Scholar] [CrossRef]
- Tanimizu, T.; Kono, K.; Kida, S. Brain Networks Activated to Form Object Recognition Memory. Brain Res. Bull. 2018, 141, 27–34. [Google Scholar] [CrossRef]
- Cunha, C.; Brambilla, R.; Thomas, K.L. A Simple Role for BDNF in Learning and Memory? Front. Mol. Neurosci. 2010, 3, 1. [Google Scholar] [CrossRef]
- Nakai, T.; Nagai, T.; Tanaka, M.; Itoh, N.; Asai, N.; Enomoto, A.; Asai, M.; Yamada, S.; Bin Saifullah, A.; Sokabe, M.; et al. Girdin Phosphorylation Is Crucial for Synaptic Plasticity and Memory: A Potential Role in the Interaction of BDNF/TrkB/Akt Signaling with NMDA Receptor. J. Neurosci. 2014, 34, 14995–15008. [Google Scholar] [CrossRef]
- Cao, C.; Rioult-Pedotti, M.S.; Migani, P.; Yu, C.J.; Tiwari, R.; Parang, K.; Spaller, M.R.; Goebel, D.J.; Marshall, J. Impairment of TrkB-PSD-95 Signaling in Angelman Syndrome. PLoS Biol. 2013, 11, e1001478. [Google Scholar] [CrossRef]
- Agrawal, R.; Noble, E.; Tyagi, E.; Zhuang, Y.; Ying, Z.; Gomez-Pinilla, F. Flavonoid Derivative 7,8-DHF Attenuates TBI Pathology via TrkB Activation. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2015, 1852, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Geng, X.; Huber, C.; Stone, C.; Ding, Y. In Search of a Dose: The Functional and Molecular Effects of Exercise on Post-Stroke Rehabilitation in Rats. Front. Cell. Neurosci. 2020, 14, 186. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Wang, X.; Xiang, Q.; Meng, X.; Peng, Y.; Du, N.; Liu, Z.; Sun, Q.; Wang, C.; Liu, X. Astaxanthin Alleviates Brain Aging in Rats by Attenuating Oxidative Stress and Increasing BDNF Levels. Food Funct. 2014, 5, 158–166. [Google Scholar] [CrossRef]
- Gulzar, S.; Benjakul, S. Characteristics and Storage Stability of Nanoliposomes Loaded with Shrimp Oil as Affected by Ultrasonication and Microfluidization. Food Chem. 2020, 310, 125916. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhang, S.; Gu, K.; Zhang, N. Preparation Of Astaxanthin-Loaded Liposomes: Characterization, Storage Stability and Antioxidant Activity. CYTA-J. Food 2018, 16, 607–618. [Google Scholar] [CrossRef]
- Ahn, H.; Park, J.H. Liposomal Delivery Systems for Intestinal Lymphatic Drug Transport. Biomater. Res. 2016, 20, 36. [Google Scholar] [CrossRef]
- Angelova, P.R.; Abramov, A.Y. Role of mitochondrial ROS in the brain: From physiology to neurodegeneration. FEBS Lett. 2018, 592, 692–702. [Google Scholar] [CrossRef]
- Galasso, C.; Orefice, I.; Pellone, P.; Cirino, P.; Miele, R.; Ianora, A.; Brunet, C.; Sansone, C. On the neuroprotective role of astaxanthin: New perspectives? Mar. Drugs 2018, 16, 247. [Google Scholar] [CrossRef]
- Wang, F.; Teng, Z.; Liu, D.; Wang, Y.; Lou, J.; Dong, Z. β-Caryophyllene Liposomes Attenuate Neurovascular unit Damage after Subarachnoid Hemorrhage in Rats. Neurochem. Res. 2020, 45, 1758–1768. [Google Scholar] [CrossRef]
- Hernandez, C.; Shukla, S. Liposome Based Drug Delivery as a Potential Treatment Option for Alzheimer’s Disease. Neural Regen. Res. 2022, 17, 1190. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | Treatment |
|---|---|
| 1. Control | Propylene glycol (PG; a vehicle of EESS) |
| 2. Sham | PG + artificial cerebrospinal fluid (aCSF) a |
| 3. Amyloid-beta | PG + Aβ a |
| 4. EESS-liposome | EESS-loaded liposome 100 mg/kg BW + Aβ a |
| 5. AST-commercial | AST-commercial 10 mg/kg BW + Aβ a |
| 6. EESS | EESS 100 mg/kg BW + Aβ a |
| 7. Blank liposome | Blank liposome + Aβ a |
| Sample | Entrapment Efficiency (%) | Particle Size (nm) | Polydispersity Index | Zeta Potential (mV) |
|---|---|---|---|---|
| Blank liposome | - | 587.2 ± 12.9 | 0.40 ± 0.00 | −55.12 ± 0.61 |
| EESS-loaded liposome | 55.34 ± 0.75 | 1009.40 ± 38.8 | 0.40 ± 0.01 | −61.27 ± 0.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuedo, Z.; Chotphruethipong, L.; Raju, N.; Reudhabibadh, R.; Benjakul, S.; Chonpathompikunlert, P.; Klaypradit, W.; Hutamekalin, P. Oral Administration of Ethanolic Extract of Shrimp Shells-Loaded Liposome Protects against Aβ-Induced Memory Impairment in Rats. Foods 2022, 11, 2673. https://doi.org/10.3390/foods11172673
Kuedo Z, Chotphruethipong L, Raju N, Reudhabibadh R, Benjakul S, Chonpathompikunlert P, Klaypradit W, Hutamekalin P. Oral Administration of Ethanolic Extract of Shrimp Shells-Loaded Liposome Protects against Aβ-Induced Memory Impairment in Rats. Foods. 2022; 11(17):2673. https://doi.org/10.3390/foods11172673
Chicago/Turabian StyleKuedo, Zulkiflee, Lalita Chotphruethipong, Navaneethan Raju, Ratchaneekorn Reudhabibadh, Soottawat Benjakul, Pennapa Chonpathompikunlert, Wanwimol Klaypradit, and Pilaiwanwadee Hutamekalin. 2022. "Oral Administration of Ethanolic Extract of Shrimp Shells-Loaded Liposome Protects against Aβ-Induced Memory Impairment in Rats" Foods 11, no. 17: 2673. https://doi.org/10.3390/foods11172673
APA StyleKuedo, Z., Chotphruethipong, L., Raju, N., Reudhabibadh, R., Benjakul, S., Chonpathompikunlert, P., Klaypradit, W., & Hutamekalin, P. (2022). Oral Administration of Ethanolic Extract of Shrimp Shells-Loaded Liposome Protects against Aβ-Induced Memory Impairment in Rats. Foods, 11(17), 2673. https://doi.org/10.3390/foods11172673