
Evidence for Quercetin as a Dietary Supplement for the Treatment of Cardio-Metabolic Diseases in Pregnancy: A Review in Rodent Models
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Quercetin
3. Quercetin Intervention in Rodents during Pregnancy and Lactation
4. Quercetin Intervention in Gestational Diabetes Mellitus
4.1. GDM: A Pregnant Disorder Linked to Oxidative Stress and Inflammation
4.2. Quercetin Supplementation in Rodent Models of GDM
5. Quercetin Intervention in Gestational Undernutrition
5.1. Gestational Undernutrition: A Pregnant Disorder Linked to Oxidative Stress and Inflammation
5.2. Quercetin Supplementation in Rodent Models of Gestational Undernutrition
6. Quercetin Supplementation in High-Fat Diet Rodent Models Inducing Maternal Overweight and Dyslipidemia
7. Quercetin Supplementation in Rodent Models of Pregnancy Hypertension Syndromes
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Plagemann, A.; Harder, T.; Schellong, K.; Schulz, S.; Stupin, J.H. Early postnatal life as a critical time window for determination of long-term metabolic health. Best Pract. Res. Clin. Endocrinol. Metab. 2012, 26, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.D.; Hanson, M.A. The developmental origins of the metabolic syndrome. Trends Endocrinol. Metab. 2004, 15, 183–187. [Google Scholar] [CrossRef]
- Agarwal, P.; Morriseau, T.S.; Kereliuk, S.M.; Doucette, C.A.; Wicklow, B.A.; Dolinsky, V.W. Maternal obesity, diabetes during pregnancy and epigenetic mechanisms that influence the developmental origins of cardiometabolic disease in the offspring. Crit. Rev. Clin. Lab. Sci. 2018, 55, 71–101. [Google Scholar] [CrossRef] [PubMed]
- Fall, C.H.D.; Kumaran, K. Metabolic programming in early life in humans. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180123. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomas-Barberan, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Pham, N.M.; Do, V.V.; Lee, A.H. Polyphenol-rich foods and risk of gestational diabetes: A systematic review and meta-analysis. Eur. J. Clin. Nutr. 2019, 73, 647–656. [Google Scholar] [CrossRef]
- Tenorio, M.B.; Ferreira, R.C.; Moura, F.A.; Bueno, N.B.; Goulart, M.O.F.; Oliveira, A.C.M. Oral antioxidant therapy for prevention and treatment of preeclampsia: Meta-analysis of randomized controlled trials. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 865–876. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Vanhees, K.; van Schooten, F.J.; van Waalwijk van Doorn-Khosrovani, S.B.; van Helden, S.; Munnia, A.; Peluso, M.; Briede, J.J.; Haenen, G.R.; Godschalk, R.W. Intrauterine exposure to flavonoids modifies antioxidant status at adulthood and decreases oxidative stress-induced DNA damage. Free Radic. Biol. Med. 2013, 57, 154–161. [Google Scholar] [CrossRef]
- Anachuna, K.K.; Ekhoye, E.I.; Iyare, C.; Katchy, N.; Ben-Azu, B.; Adeniyi, D.B.; Daubry, T.M.E.; Iyare, E. Altered gestational outcomes and delayed pubertal onset in prenatally and early postnatally food restricted male and female rats: Mitigation by quercetin and kaempferol. Int. J. Physiol. Pathophysiol. Pharmacol. 2020, 12, 115–127. [Google Scholar]
- Takashima, M.; Tanaka, W.; Matsuyama, H.; Tajiri, H.; Sakakibara, H. Maternal Quercetin Consumption during Pregnancy May Help Regulate Total Cholesterol/HDL-Cholesterol Ratio without Effect on Cholesterol Levels in Male Progeny Consuming High-Fat Diet. Nutrients 2021, 13, 1242. [Google Scholar] [CrossRef]
- Li, Q.; Yin, L.; Si, Y.; Zhang, C.; Meng, Y.; Yang, W. The bioflavonoid quercetin improves pathophysiology in a rat model of preeclampsia. Biomed. Pharmacother. 2020, 127, 110122. [Google Scholar] [CrossRef]
- Ozarowski, M.; Mikolajczak, P.L.; Kujawski, R.; Wielgus, K.; Klejewski, A.; Wolski, H.; Seremak-Mrozikiewicz, A. Pharmacological Effect of Quercetin in Hypertension and Its Potential Application in Pregnancy-Induced Hypertension: Review of In Vitro, In Vivo, and Clinical Studies. Evid. Based Complement. Altern. Med. 2018, 2018, 7421489. [Google Scholar] [CrossRef]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef]
- Kondratyuk, T.P.; Pezzuto, J.M. Natural product polyphenols of relevance to human health. Pharm. Biol. 2004, 42, 46–63. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Zhang, L.; Han, Z.; Granato, D. Polyphenols in foods: Classification, methods of identification, and nutritional aspects in human health. Adv. Food Nutr. Res. 2021, 98, 1–33. [Google Scholar] [CrossRef]
- Kawabata, K.; Mukai, R.; Ishisaka, A. Quercetin and related polyphenols: New insights and implications for their bioactivity and bioavailability. Food Funct. 2015, 6, 1399–1417. [Google Scholar] [CrossRef]
- Ulusoy, H.G.; Sanlier, N. A minireview of quercetin: From its metabolism to possible mechanisms of its biological activities. Crit. Rev. Food Sci. Nutr. 2020, 60, 3290–3303. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintac, D.; Majkic, T.; Bekvalac, K.; Orcic, D.; Mimica-Dukic, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Andres, S.; Pevny, S.; Ziegenhagen, R.; Bakhiya, N.; Schafer, B.; Hirsch-Ernst, K.I.; Lampen, A. Safety Aspects of the Use of Quercetin as a Dietary Supplement. Mol. Nutr. Food Res. 2018, 62, 1700447. [Google Scholar] [CrossRef] [PubMed]
- Mirsafaei, L.; Reiner, Z.; Shafabakhsh, R.; Asemi, Z. Molecular and Biological Functions of Quercetin as a Natural Solution for Cardiovascular Disease Prevention and Treatment. Plant Foods Hum. Nutr. 2020, 75, 307–315. [Google Scholar] [CrossRef]
- Tanir, H.M.; Sener, T.; Inal, M.; Akyuz, F.; Uzuner, K.; Sivri, E. Effect of quercetine and glutathione on the level of superoxide dismutase, catalase, malonyldialdehyde, blood pressure and neonatal outcome in a rat model of pre-eclampsia induced by NG-nitro-L-arginine-methyl ester. Eur. J. Obstet. Gynecol. Reprod Biol. 2005, 118, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Tan, C.; Meng, F.; Liu, P.; Reece, E.A.; Zhao, Z. Amelioration of intracellular stress and reduction of neural tube defects in embryos of diabetic mice by phytochemical quercetin. Sci. Rep. 2016, 6, 21491. [Google Scholar] [CrossRef]
- Vanhees, K.; Godschalk, R.W.; Sanders, A.; van Waalwijk van Doorn-Khosrovani, S.B.; van Schooten, F.J. Maternal quercetin intake during pregnancy results in an adapted iron homeostasis at adulthood. Toxicology 2011, 290, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Sun, L.; Lu, L.; Zhong, X.; Ma, Y.; Qin, J. Effects of quercetin on CYP450 and cytokines in Aroclor 1254 injured endometrial cells of the pregnant rats. Biomed Res. Int. 2014, 2014, 497508. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhou, Y.; Qin, Y.; Yu, L.; Li, R.; Chen, Y.; Xu, Y. Quercetin Intervention Alleviates Offspring’s Oxidative Stress, Inflammation, and Tight Junction Damage in the Colon Induced by Maternal Fine Particulate Matter (PM2.5) Exposure through the Reduction of Bacteroides. Nutrients 2020, 12, 3095. [Google Scholar] [CrossRef]
- Sistani Karampour, N.; Arzi, A.; Najafzadeh Varzi, H.; Mohammadian, B.; Rezaei, M. Quercetin preventive effects on theophylline-induced anomalies in rat embryo. Jundishapur J. Nat. Pharm. Prod. 2014, 9, e17834. [Google Scholar] [CrossRef]
- Lin, X.; Peng, Q.; Zhang, J.; Li, X.; Huang, J.; Duan, S.; Zhang, W. Quercetin Prevents Lipopolysaccharide-Induced Experimental Preterm Labor in Mice and Increases Offspring Survival Rate. Reprod. Sci. 2020, 27, 1047–1057. [Google Scholar] [CrossRef]
- Lin, M.; Wang, N.; Yao, B.; Zhong, Y.; Lin, Y.; You, T. Quercetin improves postpartum hypogalactia in milk-deficient mice via stimulating prolactin production in pituitary gland. Phytother. Res. 2018, 32, 1511–1520. [Google Scholar] [CrossRef]
- Plows, J.F.; Stanley, J.L.; Baker, P.N.; Reynolds, C.M.; Vickers, M.H. The Pathophysiology of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2018, 19, 3342. [Google Scholar] [CrossRef] [Green Version]
- Johns, E.C.; Denison, F.C.; Norman, J.E.; Reynolds, R.M. Gestational Diabetes Mellitus: Mechanisms, Treatment, and Complications. Trends Endocrinol. Metab. 2018, 29, 743–754. [Google Scholar] [CrossRef]
- Lappas, M.; Hiden, U.; Desoye, G.; Froehlich, J.; Hauguel-de Mouzon, S.; Jawerbaum, A. The role of oxidative stress in the pathophysiology of gestational diabetes mellitus. Antioxid. Redox. Signal. 2011, 15, 3061–3100. [Google Scholar] [CrossRef]
- Kolyada, A.Y.; Savikovsky, N.; Madias, N.E. Transcriptional regulation of the human iNOS gene in vascular-smooth-muscle cells and macrophages: Evidence for tissue specificity. Biochem. Biophys. Res. Commun. 1996, 220, 600–605. [Google Scholar] [CrossRef]
- Nunokawa, Y.; Oikawa, S.; Tanaka, S. Human inducible nitric oxide synthase gene is transcriptionally regulated by nuclear factor-kappaB dependent mechanism. Biochem. Biophys. Res. Commun. 1996, 223, 347–352. [Google Scholar] [CrossRef]
- Jawerbaum, A.; Gonzalez, E. The role of alterations in arachidonic acid metabolism and nitric oxide homeostasis in rat models of diabetes during early pregnancy. Curr. Pharm. Des. 2005, 11, 1327–1342. [Google Scholar] [CrossRef]
- Sugimura, Y.; Murase, T.; Oyama, K.; Uchida, A.; Sato, N.; Hayasaka, S.; Kano, Y.; Takagishi, Y.; Hayashi, Y.; Oiso, Y.; et al. Prevention of neural tube defects by loss of function of inducible nitric oxide synthase in fetuses of a mouse model of streptozotocin-induced diabetes. Diabetologia 2009, 52, 962–971. [Google Scholar] [CrossRef]
- Jawerbaum, A.; Sinner, D.; White, V.; Pustovrh, C.; Capobianco, E.; Gonzalez, E. Modulation of nitric oxide concentration and lipid metabolism by 15-deoxy Delta12,14prostaglandin J2 in embryos from control and diabetic rats during early organogenesis. Reproduction 2002, 124, 625–631. [Google Scholar] [CrossRef]
- Tan, C.; Meng, F.; Reece, E.A.; Zhao, Z. Modulation of nuclear factor-kappaB signaling and reduction of neural tube defects by quercetin-3-glucoside in embryos of diabetic mice. Am. J. Obstet. Gynecol. 2018, 219, 197.e1–197.e8. [Google Scholar] [CrossRef]
- Bolouki, A.; Zal, F.; Alaee, S. Ameliorative effects of quercetin on the preimplantation embryos development in diabetic pregnant mice. J. Obstet. Gynaecol. Res. 2020, 46, 736–744. [Google Scholar] [CrossRef]
- Mahabady, M.K.; Shamsi, M.M.; Ranjbar, R.; Tabandeh, M.R.; Khazaeel, K. Quercetin improved histological structure and upregulated adiponectin and adiponectin receptors in the placenta of rats with gestational diabetes mellitus. Placenta 2021, 106, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Lillycrop, K.A.; Burdge, G.C. Epigenetic mechanisms linking early nutrition to long term health. Best Pract. Res. Clin. Endocrinol. Metab. 2012, 26, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, D.C.; Ferraz-Pereira, K.N.; Visco, D.B.; Pontes, P.B.; Chaves, W.F.; Guzman-Quevedo, O.; Manhaes-de-Castro, R.; Toscano, A.E. Perinatal undernutrition associated to experimental model of cerebral palsy increases adverse effects on chewing in young rats. Physiol. Behav. 2017, 173, 69–78. [Google Scholar] [CrossRef]
- Silva, K.O.; Pereira Sda, C.; Portovedo, M.; Milanski, M.; Galindo, L.C.; Guzman-Quevedo, O.; Manhaes-de-Castro, R.; Toscano, A.E. Effects of maternal low-protein diet on parameters of locomotor activity in a rat model of cerebral palsy. Int. J. Dev. Neurosci. 2016, 52, 38–45. [Google Scholar] [CrossRef]
- De Brito Alves, J.L.; Toscano, A.E.; da Costa-Silva, J.H.; Vidal, H.; Leandro, C.G.; Pirola, L. Transcriptional response of skeletal muscle to a low protein perinatal diet in rat offspring at different ages: The role of key enzymes of glucose-fatty acid oxidation. J. Nutr. Biochem. 2017, 41, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Ferraz-Pereira, K.N.; da Silva Aragao, R.; Verdier, D.; Toscano, A.E.; Lacerda, D.C.; Manhaes-de-Castro, R.; Kolta, A. Neonatal low-protein diet reduces the masticatory efficiency in rats. Br. J. Nutr. 2015, 114, 1515–1530. [Google Scholar] [CrossRef] [PubMed]
- Chotechuang, N.; Azzout-Marniche, D.; Bos, C.; Chaumontet, C.; Gausseres, N.; Steiler, T.; Gaudichon, C.; Tome, D. mTOR, AMPK, and GCN2 coordinate the adaptation of hepatic energy metabolic pathways in response to protein intake in the rat. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E1313–E1323. [Google Scholar] [CrossRef] [PubMed]
- De Brito Alves, J.L.; Nogueira, V.O.; de Oliveira, G.B.; da Silva, G.S.; Wanderley, A.G.; Leandro, C.G.; Costa-Silva, J.H. Short- and long-term effects of a maternal low-protein diet on ventilation, O(2)/CO(2) chemoreception and arterial blood pressure in male rat offspring. Br. J. Nutr. 2014, 111, 606–615. [Google Scholar] [CrossRef]
- Barros, M.A.; De Brito Alves, J.L.; Nogueira, V.O.; Wanderley, A.G.; Costa-Silva, J.H. Maternal low-protein diet induces changes in the cardiovascular autonomic modulation in male rat offspring. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 123–130. [Google Scholar] [CrossRef]
- Barros, M.A.V.; de Brito Alves, J.L.; Barros, R.G.N.; Santana, E.S.M.; Nasser, S.; Tourneur, Y.; Leandro, C.V.G.; Vidal, H.; Pirola, L.; Costa-Silva, J.H. Effects of maternal protein restriction on central and peripheral renin-angiotensin systems in male rat offspring. Life Sci. 2020, 263, 118574. [Google Scholar] [CrossRef]
- Inadera, H. Developmental origins of obesity and type 2 diabetes: Molecular aspects and role of chemicals. Environ. Health Prev. Med. 2013, 18, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Li, X.; Niu, W.; Ma, G.; Sun, Q.; Bi, Y.; Guo, Z.; Ren, D.; Hu, J.; Yuan, F.; et al. Metabolomic profiling on rat brain of prenatal malnutrition: Implicated for oxidative stress and schizophrenia. Metab. Brain Dis. 2019, 34, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- De Brito Alves, J.L.; de Oliveira, J.M.; Ferreira, D.J.; Barros, M.A.; Nogueira, V.O.; Alves, D.S.; Vidal, H.; Leandro, C.G.; Lagranha, C.J.; Pirola, L.; et al. Maternal protein restriction induced-hypertension is associated to oxidative disruption at transcriptional and functional levels in the medulla oblongata. Clin. Exp. Pharmacol. Physiol. 2016, 43, 1177–1184. [Google Scholar] [CrossRef]
- Anachuna, K.K.; Moke, G.E.; Iyare, C.; Katchy, N.; Ben-Azu, B.; Adeniyi, B.; Nwogueze, B.C.; Iyare, E. Prenatal and early postnatal food restrictions cause changes in brain oxidative status and orexigenic/anorexigenic hormones in the offspring of rats: Prevention by quercetin and kaempferol. Curr. Res. Pharmacol. Drug Discov. 2020, 1, 39–52. [Google Scholar] [CrossRef]
- Sato, S.; Mukai, Y.; Saito, T. Quercetin intake during lactation modulates the AMP-activated protein kinase pathway in the livers of adult male rat offspring programmed by maternal protein restriction. J. Nutr. Biochem. 2013, 24, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Claycombe, K.J.; Brissette, C.A.; Ghribi, O. Epigenetics of inflammation, maternal infection, and nutrition. J. Nutr. 2015, 145, 1109S–1115S. [Google Scholar] [CrossRef] [PubMed]
- Poston, L.; Harthoorn, L.F.; Van Der Beek, E.M. Obesity in pregnancy: Implications for the mother and lifelong health of the child. A consensus statement. Pediatr. Res. 2011, 69, 175–180. [Google Scholar] [CrossRef]
- Riley, L.; Wertz, M.; McDowell, I. Obesity in Pregnancy: Risks and Management. Am. Fam. Physician 2018, 97, 559–561. [Google Scholar]
- Wu, Z.; Zhao, J.; Xu, H.; Lyv, Y.; Feng, X.; Fang, Y.; Xu, Y. Maternal quercetin administration during gestation and lactation decrease endoplasmic reticulum stress and related inflammation in the adult offspring of obese female rats. Eur. J. Nutr. 2014, 53, 1669–1683. [Google Scholar] [CrossRef]
- Adeyemi, T.E.; Ajonijebu, D.C.; Channa, M.L.; Nadar, A. Postnatally induced metabolic and oxidative changes associated with maternal high-fat consumption were mildly affected by Quercetin-3-O-rutinoside treatment in rats. Heliyon 2021, 7, e06847. [Google Scholar] [CrossRef]
- Adeyemi, T.E.; Channa, M.L.; Nadar, A. Evaluation of maternal high-fat diet and Quercetin-3-O-rutinoside treatment on the reproductive profile of diet naive male offspring. Life Sci. 2021, 271, 119179. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Oest, M.E.; Prater, M.R. Intrauterine exposure to high saturated fat diet elevates risk of adult-onset chronic diseases in C57BL/6 mice. Birth Defects Res. B Dev. Reprod. Toxicol. 2009, 86, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, D.C.; Urquiza-Martinez, M.V.; Manhaes-de-Castro, R.; Visco, D.B.; Derosier, C.; Mercado-Camargo, R.; Torner, L.; Toscano, A.E.; Guzman-Quevedo, O. Metabolic and neurological consequences of the treatment with polyphenols: A systematic review in rodent models of noncommunicable diseases. Nutr. Neurosci. 2021, 25, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.; Muro, T.; Kobori, M.; Nishihira, J. Effect of Daily Ingestion of Quercetin-Rich Onion Powder for 12 Weeks on Visceral Fat: A Randomised, Double-Blind, Placebo-Controlled, Parallel-Group Study. Nutrients 2019, 12, 91. [Google Scholar] [CrossRef]
- Khorshidi, M.; Moini, A.; Alipoor, E.; Rezvan, N.; Gorgani-Firuzjaee, S.; Yaseri, M.; Hosseinzadeh-Attar, M.J. The effects of quercetin supplementation on metabolic and hormonal parameters as well as plasma concentration and gene expression of resistin in overweight or obese women with polycystic ovary syndrome. Phytother. Res. 2018, 32, 2282–2289. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, X.; Li, X.; Gaman, M.A.; Kord-Varkaneh, H.; Rahmani, J.; Salehi-Sahlabadi, A.; Day, A.S.; Xu, Y. Serum Vitamin D Levels and Risk of Liver Cancer: A Systematic Review and Dose-Response Meta-Analysis of Cohort Studies. Nutr. Cancer 2020, 73, 1–9. [Google Scholar] [CrossRef]
- Yang, S.; Song, L.; Shi, X.; Zhao, N.; Ma, Y. Ameliorative effects of pre-eclampsia by quercetin supplement to aspirin in a rat model induced by L-NAME. Biomed. Pharmacother. 2019, 116, 108969. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, S.; Song, H. Quercetin attenuates reduced uterine perfusion pressure -induced hypertension in pregnant rats through regulation of endothelin-1 and endothelin-1 type A receptor. Lipids Health Dis. 2020, 19, 180. [Google Scholar] [CrossRef]
- Biesinger, S.; Michaels, H.A.; Quadros, A.S.; Qian, Y.; Rabovsky, A.B.; Badger, R.S.; Jalili, T. A combination of isolated phytochemicals and botanical extracts lowers diastolic blood pressure in a randomized controlled trial of hypertensive subjects. Eur. J. Clin. Nutr. 2016, 70, 10–16. [Google Scholar] [CrossRef]
- Edwards, R.L.; Lyon, T.; Litwin, S.E.; Rabovsky, A.; Symons, J.D.; Jalili, T. Quercetin reduces blood pressure in hypertensive subjects. J. Nutr. 2007, 137, 2405–2411. [Google Scholar] [CrossRef]
- Larson, A.; Witman, M.A.; Guo, Y.; Ives, S.; Richardson, R.S.; Bruno, R.S.; Jalili, T.; Symons, J.D. Acute, quercetin-induced reductions in blood pressure in hypertensive individuals are not secondary to lower plasma angiotensin-converting enzyme activity or endothelin-1: Nitric oxide. Nutr. Res. 2012, 32, 557–564. [Google Scholar] [CrossRef]
- Gongol, B.; Sari, I.; Bryant, T.; Rosete, G.; Marin, T. AMPK: An Epigenetic Landscape Modulator. Int. J. Mol. Sci. 2018, 19, 3238. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Mukai, Y. Modulation of Chronic Inflammation by Quercetin: The Beneficial Effects on Obesity. J. Inflamm. Res. 2020, 13, 421–431. [Google Scholar] [CrossRef]
- Huang, H.; Liao, D.; Dong, Y.; Pu, R. Effect of quercetin supplementation on plasma lipid profiles, blood pressure, and glucose levels: A systematic review and meta-analysis. Nutr. Rev. 2020, 78, 615–626. [Google Scholar] [CrossRef]
- Tabrizi, R.; Tamtaji, O.R.; Mirhosseini, N.; Lankarani, K.B.; Akbari, M.; Heydari, S.T.; Dadgostar, E.; Asemi, Z. The effects of quercetin supplementation on lipid profiles and inflammatory markers among patients with metabolic syndrome and related disorders: A systematic review and meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2020, 60, 1855–1868. [Google Scholar] [CrossRef]
- Han, M.K.; Barreto, T.A.; Martinez, F.J.; Comstock, A.T.; Sajjan, U.S. Randomised clinical trial to determine the safety of quercetin supplementation in patients with chronic obstructive pulmonary disease. BMJ Open Respir. Res. 2020, 7, e000392. [Google Scholar] [CrossRef] [PubMed]
- Di Pierro, F.; Derosa, G.; Maffioli, P.; Bertuccioli, A.; Togni, S.; Riva, A.; Allegrini, P.; Khan, A.; Khan, S.; Khan, B.A.; et al. Possible Therapeutic Effects of Adjuvant Quercetin Supplementation Against Early-Stage COVID-19 Infection: A Prospective, Randomized, Controlled, and Open-Label Study. Int. J. Gen. Med. 2021, 14, 2359–2366. [Google Scholar] [CrossRef]
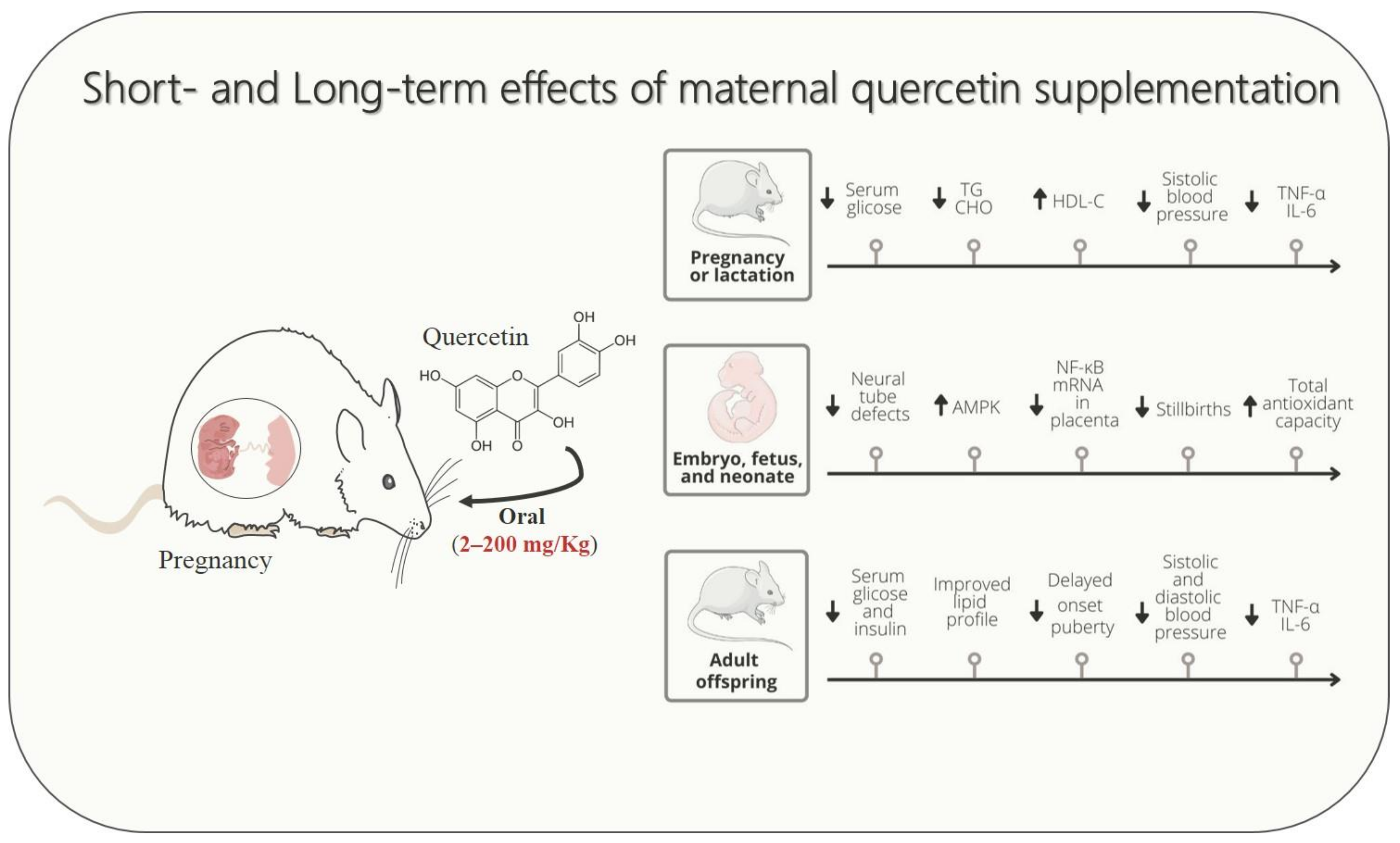
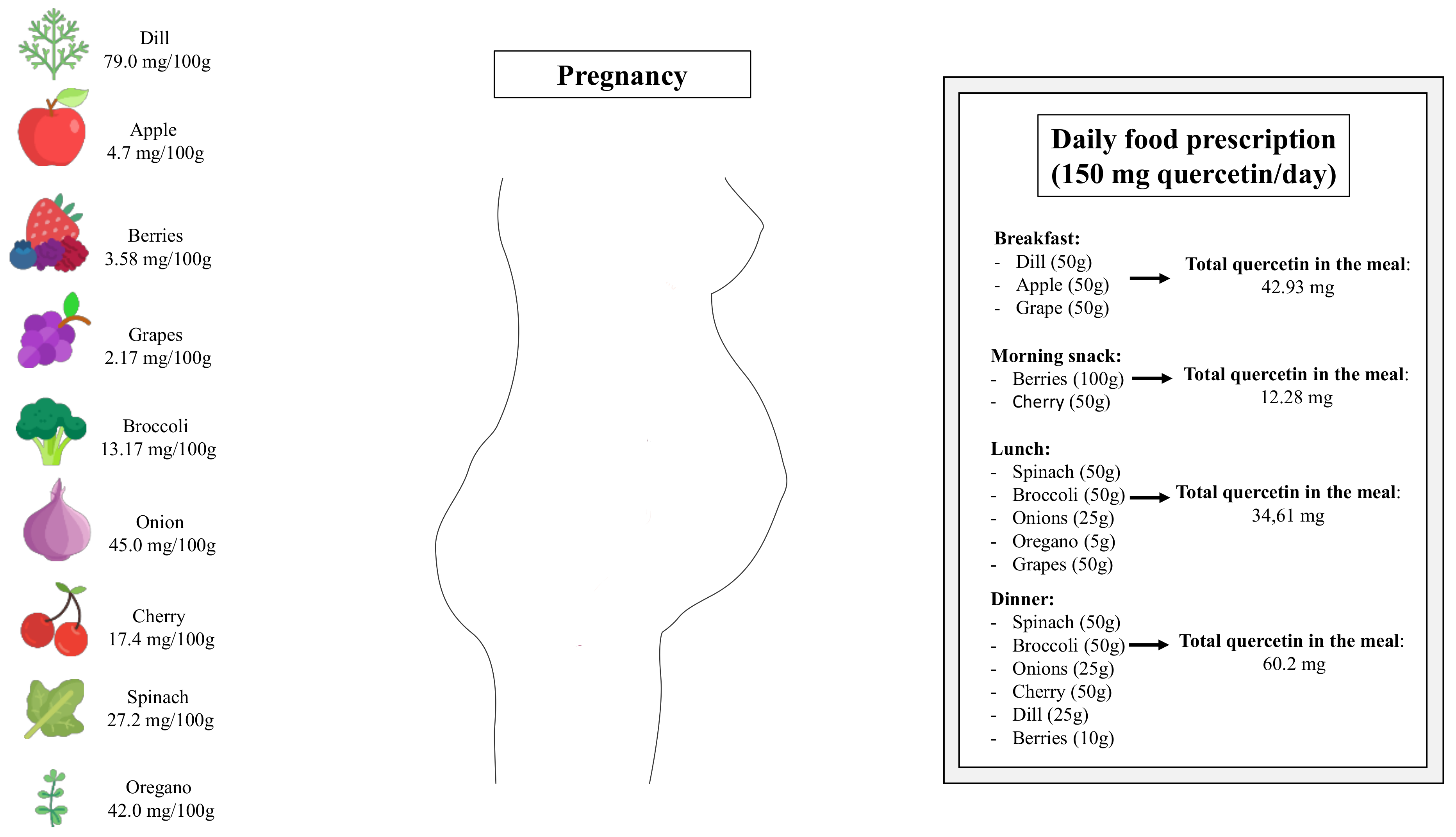

{kind=link}
{kind=link}
| Reference | Type of Study | Dose | Duration | Maternal Outcomes | Embryonic, Fetal, and Neonatal Outcomes |
|---|---|---|---|---|---|
| Cao et al., 2016 [24] | Experimental (mice) | 100 mg/kg | Embryonic period: 7.5 to 10.5. | No difference in maternal glucose. | ↓ Decreased the neural tube defects rate; ↓ Apoptosis in the dorsal neural tube of the embryos and levels of Caspase-3; ↓ Nos2 level in the neural tissues of the embryos; ↓ Levels of protein S nitrosylation in the neural tube; ↑ Enzyme levels of SOD1 and SOD2; ↓ Levels of 4-HNE and MDA; ↑ Expression of redox regulating and DNA damage; There was the presence of quercetin metabolites in the embryo. |
| Tan et al., 2018 [39] | Experimental (mice) | 100 mg/kg | Embryonic period: 6.5 to 9.5. | The authors did not evaluate maternal outcomes. | ↓ Neural tube defects; ↓ The apoptotic signals were lower in the neural tube regions; ↓ Nos2 expression in the embryos. ↑ The levels of Sod1 in the embryos; ↓ Nitrosative and oxidative stresses in the endoplasmic reticulum (ER); ↓ The expression of p65. |
| Bolouki et al., 2020 [40] | Experimental (mice) | 30 mg/kg | 4 weeks before conception. | ↓ Levels of blood glucose; ↑ The number of embryos per mouse; ↑ Levels of serum 17β-estradiol; ↑ Estradiol/progesterone ratio. | ↑ The embryo morphological distribution to the well-developed stages; ↑ IGF1r, integrin αvβ3, and Cox2 mRNA express these genes in the blastocyst; ↓ Expression of the Caspase3 gene; Can activate the nuclear Wnt-β-catenin signaling pathway. |
| Mahabady et al., 2021 [41] | Experimental (rats) | 75 mg/kg | On 0, 7, 14, and 20 days of gestation. | The authors did not evaluate maternal outcomes. | ↓ LIM thickness; ↓ Mean the number of glycogen cells; ↑ Increased placental adiponectin expression; ↓ Placental expression of AdipoR1 and AdipoR2. |
| Reference | Type of Study | Dose | Duration | Maternal Outcomes | Embryonic, Fetal, and Neonatal Outcomes | Adulthood Outcomes |
|---|---|---|---|---|---|---|
| Anachuna et al., 2020 [10] | Experimental (rats) | 50, 100 and 200 mg/kg | 21–22 days—during pregnancy e/or 22 days during weaning. | The authors did not evaluate maternal outcomes. | ↓ Stillbirths; ↑ Nose-tail lenghts at P1 and P22. | ↓ Onset puberty. |
| Anachuna et al., 2020 [54] | Experimental (rats) | 50, 100 and 200 mg/kg | 21–22 days—during pregnancy e/or 22 days during weaning. | ↑ Maternal weight. | ↑ Body weight undernourished rats until weaning | ↓ Body weight; ↑ Leptin levels; ↓ Ghrelin levels; ↓ Brain oxidative stress. |
| Sato et al., 2013 [55] | Experimental (rats) | 80–110 mg/day | 10 to 22 postnatal days. | The authors did not evaluate maternal outcomes. | ↓ TG blood level; ↑ AMPK. | ↑ Body weight; ↑ Adiponectin; ↑ AMPK. |
| Reference | Type of Study | Dose | Duration | Maternal Outcomes | Embryonic, Fetal and Neonatal Outcomes | Adulthood Outcomes |
|---|---|---|---|---|---|---|
| Adeyemi et al., 2021 [60] | Experimental (rats subjected to HFD contained 45% of fat) | 150 mg/kg | From the beginning of gestation to gestational day 19. | The authors did not evaluate maternal outcomes. | ↓ MDA and NO concentration in the placenta and liver tissues in neonate male rats; ↓ SOD concentration in the placenta and liver tissues in neonatal rats; ↑ Total antioxidant capacity of the liver in neonates. | The authors did not evaluate adulthood outcomes. |
| Adeyemi et al., 2021 [61] | Experimental (rats subjected to HFD contained 45% of fat) | 150 mg/kg | From the beginning of gestation to gestational day 19. | The authors did not evaluate maternal outcomes. | ↓ Expression of mRNA of TNF-α and IL-1β in placenta and hypothalamus of neonatal rats; ↑ NF-κB mRNA level in placenta. | The authors did not evaluate adulthood outcomes. |
| Liang et al., 2009 [62] | Experimental (mice subjected to HFD contained 60% of fat) | 66 mg/kg | For 4 weeks before breeding. | The authors did not evaluate maternal outcomes. | The authors did not evaluate embryonic, fetal, and neonatal outcomes. | ↓ Blood glucose and plasma insulin levels of 6- and 12-month offspring; ↓ Systolic and diastolic blood pressure of 6- and 12-month offspring. |
| Takashima et al., 2021 [11] | Experimental (mice subjected to HFD contained 30% of fat) | 1.0% | Before breeding, throughout gestation, lactation until 13 weeks of postnatal days. | ↑ Body weight; ↑ Liver weight. | ↑ Total CHO, non-HDL-C, and HDL-C levels of pups; ↓ Gastric inhibitory polypeptide levels of pups. | The authors did not evaluate adulthood outcomes. |
| Wu et al., 2014 [59] | Experimental (rats subjected to HFD contained about 42% of fat) | 50, 100 or 200 mg/kg | Throughout gestation and lactation. | Did not change body weight and blood glucose levels; Improved lipid profile (↓ TG, CHO, and ↑ HDL-C levels). | The authors did not evaluate embryonic, fetal, and neonatal outcomes. | ↓ Blood glucose and insulin levels in adult rats; Improved lipid profile (↓ TG, CHO) in adult rats; ↓ Expression of TNF-α, IL-6 in adult rats; ↓ Endoplasmic reticulum stress in liver and adipose tissues; ↓ In p-JNK and NF-κB protein expression. |
| Reference | Type of Study | Dose | Duration | Maternal Outcomes | Embryonic, Fetal and Neonatal Outcomes |
|---|---|---|---|---|---|
| Li et al., 2020 [12] | Experimental (rats subjected to preeclampsia model) | The authors did not inform the dose of treatment. | The authors did not inform the duration of treatment. | ↓ Systolic blood pressure; ↓ Proteinuria. | Reversed imbalance of angiogenic factors production in the placenta; ↓ Placental growth factor; ↓ Placental expression of TNF-α, IL-6, and MCP-1 levels; ↓ Placental MDA; ↑ Placenta weight; ↓ Reabsorbed fetuses %; ↑ Pups weight; ↑ Placenta weight. |
| Sun et al., 2020 [68] | Experimental (rats subjected to gestational hypertension) | 10 mg/kg, 20 mg/kg, or 50 mg/kg. | From gestational day 14 to gestational day 21. | ↓ Systolic and diastolic blood pressure; ↓ Plasma level of ET-1, sFLT1; ↑ Plasma level of VEGF; ↓ Plasma levels of TNF-α and IL-6, and ↑ IL-10. | ↓ ET-1, ETAR expression in placenta tissue; ↑ Fetal weight; Did not change placental weight; ↑ Fetal weight/ Placenta weight; ↓ Reabsorbed fetuses %. |
| Tanir et al., 2005 [23] | Experimental (rats subjected to preeclampsia model) | 10 mg/kg | Single administration at gestational day 17. | Did not change blood pressure;↓ Plasma MDA levels; ↓ Erythrocyte CAT and SOD levels; ↓ Proteinuria. | Did not change the birth weight of pups; ↑ Neonatal survival (higher percentages of liveborn pups and lower rates of dead pups). |
| Yang et al., 2019 [67] | Experimental (rats subjected to preeclampsia model) | 2 mg/kg | From gestational day 4 to gestational day 19. | ↓ Proteinuria; ↓ Plasma MDA levels; Did not change TNF-α; ↓ Plasma levels of IL-6. | Did not change placental MDA; Did not change placental expression of TNF-α and IL-6; Did not change relative mRNA expression of uterus VEGF and sFlt-1. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, P.C.T.d.; de Souza, E.L.; Lacerda, D.C.; Cruz Neto, J.P.R.; Sales, L.C.S.d.; Silva Luis, C.C.; Pontes, P.B.; Cavalcanti Neto, M.P.; de Brito Alves, J.L. Evidence for Quercetin as a Dietary Supplement for the Treatment of Cardio-Metabolic Diseases in Pregnancy: A Review in Rodent Models. Foods 2022, 11, 2772. https://doi.org/10.3390/foods11182772
Costa PCTd, de Souza EL, Lacerda DC, Cruz Neto JPR, Sales LCSd, Silva Luis CC, Pontes PB, Cavalcanti Neto MP, de Brito Alves JL. Evidence for Quercetin as a Dietary Supplement for the Treatment of Cardio-Metabolic Diseases in Pregnancy: A Review in Rodent Models. Foods. 2022; 11(18):2772. https://doi.org/10.3390/foods11182772
Chicago/Turabian StyleCosta, Paulo César Trindade da, Evandro Leite de Souza, Diego Cabral Lacerda, José Patrocínio Ribeiro Cruz Neto, Ludmilla Christine Silva de Sales, Cristiane Cosmo Silva Luis, Paula Brielle Pontes, Marinaldo Pacífico Cavalcanti Neto, and José Luiz de Brito Alves. 2022. "Evidence for Quercetin as a Dietary Supplement for the Treatment of Cardio-Metabolic Diseases in Pregnancy: A Review in Rodent Models" Foods 11, no. 18: 2772. https://doi.org/10.3390/foods11182772
APA StyleCosta, P. C. T. d., de Souza, E. L., Lacerda, D. C., Cruz Neto, J. P. R., Sales, L. C. S. d., Silva Luis, C. C., Pontes, P. B., Cavalcanti Neto, M. P., & de Brito Alves, J. L. (2022). Evidence for Quercetin as a Dietary Supplement for the Treatment of Cardio-Metabolic Diseases in Pregnancy: A Review in Rodent Models. Foods, 11(18), 2772. https://doi.org/10.3390/foods11182772