

Digestive Constraints of Arthrospira platensis in Poultry and Swine Feeding


Abstract
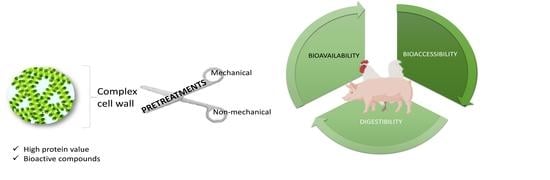
:
1. Introduction
2. Nutritional Composition of Arthrospira platensis
3. Digestibility, Bio-Accessibility and Bioavailability of Nutrients
4. Methods to Evaluate Bio-accessibility and Bioavailability of Nutrients and Protein Quality
5. Bioavailability of Arthrospira platensis Nutrients for Poultry
6. Bioavailability of Arthrospira platensis Nutrients for Swine
7. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Altmann, B.A.; Neumann, C.; Velten, S.; Liebert, F.; Mörlein, D. Meat quality derived from high inclusion of a micro-alga or insect meal as an alternative protein source in poultry diets: A pilot study. Foods 2018, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Chaves, A.A.M.; Martins, C.F.; Carvalho, D.F.P.; Ribeiro, D.M.; Lordelo, M.; Freire, J.P.B.; de Almeida, A.M. A viewpoint on the use of microalgae as an alternative feedstuff in the context of pig and poultry feeding—A special emphasis on tropical regions. Trop. Anim. Health Prod. 2021, 53, 396. [Google Scholar] [CrossRef]
- Martins, C.F.; Ribeiro, D.M.; Costa, M.; Coelho, D.; Alfaia, C.M.; Lordelo, M.; Almeida, A.M.; Freire, J.P.B.; Prates, J.A.M. Using microalgae as a sustainable feed resource to enhance quality and nutritional value of pork and poultry meat. Foods 2021, 10, 2933. [Google Scholar] [CrossRef]
- Ljubic, A.; Safafar, H.; Holdt, S.L.; Jacobsen, C. Biomass composition of Arthrospira platensis during cultivation on industrial process water and harvesting. J. Appl. Phycol. 2018, 30, 943–954. [Google Scholar] [CrossRef]
- Neumann, U.; Derwenskus, F.; Gille, A.; Louis, S.; Schmid-Staiger, U.; Briviba, K.; Bischoff, S.C. Bioavailability and safety of nutrients from the microalgae Chlorella vulgaris, Nannochloropsis oceanica and Phaeodactylum tricornutum in c57bl/6 mice. Nutrients 2018, 10, 965. [Google Scholar] [CrossRef]
- Tavernari, F.C.; Roza, L.F.; Surek, D.; Sordi, C.; Silva, M.; Albino, L.F.T.; Migliorini, M.J.; Paiano, D.; Boiago, M.M. Apparent metabolisable energy and amino acid digestibility of microalgae Spirulina platensis as an ingredient in broiler chicken diets. Br. Poult. Sci. 2018, 59, 562–567. [Google Scholar] [CrossRef]
- Seghiri, R.; Kharbach, M.; Essamri, A. Functional composition, nutritional properties, and biological activities of Moroccan Spirulina microalga. J. Food Qual. 2019, 3, 11. [Google Scholar]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef]
- Wu, H.; Li, T.; Lv, J.; Chen, Z.; Wu, J.; Wang, N.; Wu, H.; Xiang, W. Growth and biochemical composition characteristics of Arthrospira platensis induced by simultaneous nitrogen deficiency and seawater-supplemented medium in an outdoor raceway pond in winter. Foods 2021, 10, 2974. [Google Scholar] [CrossRef]
- Cai, J.; Lovatelli, A.; Garrido Gamarro, E.; Geehan, J.; Lucente, D.; Mair, G.; Weimin, M.; Bondad-Reantaso, M.; Roubach, R. Seaweeds and Microalgae: An Overview for Unlocking Their Potential in Global Aquaculture Development; FAO Fisheries and Aquaculture Circular No. 122; FAO: Rome, Italy, 2021. [Google Scholar]
- Reboleira, J.; Freitas, R.; Pinteus, S.; Silva, J.; Alves, C.; Pedrosa, R.; Bernardino, S. Spirulina. In Nonvitamin and Nonmineral Nutritional Supplements; Nabavi, S.M., Silva, A.S., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 409–413. [Google Scholar]
- DiNicolantonio, J.J.; Bhat, A.G.; OKeefe, J. Effects of spirulina on weight loss and blood lipids: A review. Open Heart 2020, 7, e001003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, M.; Skomedal, H.; Skjånes, K.; Mazur-Marzec, H.; Toruńska-Sitarz, A.; Catala, M.; Isleten Hosoglu, M.; García-Vaquero, M. Microalgal proteins for feed, food and health. In Microalgae-Based Biofuels and Bioproducts; Gonzalez-Fernandez, C., Muñoz, R., Eds.; Woodhead Publishing: Sawston, UK, 2017; pp. 347–368. [Google Scholar]
- Toyomizu, M.; Sato, K.; Taroda, H.; Kato, T.; Akiba, Y. Effects of dietary Spirulina on meat colour in muscle of broiler chickens. Br. Poult. Sci. 2001, 42, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Omri, B.; Amraoui, M.; Tarek, A.; Lucarini, M.; Durazzo, A.; Cicero, N.; Santini, A.; Kamoun, M. Arthrospira platensis (Spirulina) supplementation on laying hens’ performance: Eggs physical, chemical, and sensorial qualities. Foods 2019, 8, 386. [Google Scholar] [CrossRef] [PubMed]
- Demarco, M.; Oliveira de Moraes, J.; Matos, Â.P.; Derner, R.B.; de Farias Neves, F.; Tribuzi, G. Digestibility, bioaccessibility and bioactivity of compounds from algae. Trends Food Sci. Technol. 2022, 121, 114–128. [Google Scholar] [CrossRef]
- Acién, F.G.; Fernández, J.M.; Magán, J.J.; Molina, E. Production cost of a real microalgae production plant and strategies to reduce it. Biotechnol. Adv. 2012, 30, 1344–1353. [Google Scholar] [CrossRef] [PubMed]
- Enzing, C.; Ploeg, M.; Barbosa, M.; Sijtsma, L. Microalgae-based products for the food and feed sector: An outlook for Europe. JRC Sci. Policy Rep. 2014, 19–37. [Google Scholar]
- Zittelli, G.C.; Mangini, S.; Unamunzaga, C. Algae as Novel Food in Europe. European Algae Biomass Association 2021. [Google Scholar]
- European Commission. Regulation (EC) No 258/97 Of The European Parliament and of the Council concerning novel foods and novel food ingredients. Off. J. Eur. Communities 1997, 43, 1–7. [Google Scholar]
- Martín-Girela, I.; Albero, B.; Tiwari, B.K.; Miguel, E.; Aznar, R. Screening of Contaminants of Emerging Concern in Microalgae Food Supplements. Separations 2020, 7, 28. [Google Scholar] [CrossRef]
- Manali, K.M.; Arunraj, R.; Kumar, T.; Ramya, M. Detection of microcystin producing cyanobacteria in Spirulina dietary supplements using multiplex HRM quantitative PCR. J. Appl. Phycol. 2017, 29, 1279–1286. [Google Scholar] [CrossRef]
- Scoglio, S. Microcystins in water and in microalgae: Do microcystins as microalgae contaminants warrant the current public alarm? Toxicol. Rep. 2018, 5, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Holman, B.W.B.; Malau-Aduli, A.E.O. Spirulina as a livestock supplement and animal feed. J. Anim. Physiol. Anim. Nutr. 2013, 97, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Seyidoglu, N.; Inan, S.; Aydin, C. A Prominent Superfood: Spirulina platensis. In Superfood and Functional Food—The Development of Superfoods and Their Roles as Medicine; TechOpen: London, UK, 2017; Volume 22, pp. 1–27. [Google Scholar]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Parsons, C.M. Unresolved issues for amino acid digestibility in poultry nutrition. J. Appl. Poult. Res. 2020, 29, 1–10. [Google Scholar] [CrossRef]
- van Milgen, J.; Dourmad, J.-Y. Concept and application of ideal protein for pigs. J. Anim. Sci. Biotechnol. 2015, 6, 15. [Google Scholar] [CrossRef]
- van Eykelenburg, C. On the morphology and ultrastructure of the cell wall of Spirulina platensis. Antonie Leeuwenhoek 1977, 43, 89–99. [Google Scholar] [CrossRef]
- Park, J.; Lee, S.; Kim, I.-S. Effect of dietary Spirulina (Arthrospira) platensis on the growth performance, antioxidant enzyme activity, nutrient digestibility, cecal microflora, excreta noxious gas emission, and breast meat quality of broiler chickens. Poult. Sci. 2018, 97, 2451–2459. [Google Scholar] [CrossRef]
- Takashi, S. Effect of administration of Spirulina on egg quality and egg components. Anim. Husb. 2003, 57, 191–195. [Google Scholar]
- Coelho, D.; Lopes, P.A.; Cardoso, V.; Ponte, P.; Brás, J.; Madeira, M.S.; Alfaia, C.M.; Bandarra, N.M.; Fontes, C.M.G.A.; Prates, J.A.M. A two-enzyme constituted mixture to improve the degradation of Arthrospira platensis microalga cell wall for monogastric diets. J. Anim. Physiol. Anim. Nutr. 2020, 104, 310–321. [Google Scholar] [CrossRef]
- Batista, A.P.; Niccolai, A.; Fradinho, P.; Fragoso, S.; Bursic, I.; Rodolfi, L.; Biondi, N.; Tredici, M.R.; Sousa, I.; Raymundo, A. Microalgae biomass as an alternative ingredient in cookies: Sensory, physical and chemical properties, antioxidant activity and in vitro digestibility. Algal Res. 2017, 26, 161–171. [Google Scholar] [CrossRef]
- Gamboa-Delgado, J.; Navarro, Y.; Nieto-López, M.; Villarreal-Cavazos, D.A.; Cruz-Suarez, L. Assimilation of dietary nitrogen supplied by fish meal and microalgal biomass from Spirulina (Arthrospira platensis) and Nannochloropsis oculata in shrimp Litopenaeus vannamei fed compound diets. J. Appl. Phycol. 2019, 31, 2379–2389. [Google Scholar] [CrossRef]
- Mišurcová, L.; Kracmar, S.; Klejdus, B.; Vacek, J. Nitrogen content, dietary Fiber, and digestibility in algal food products. Czech J. Food Sci. 2010, 28, 27–35. [Google Scholar] [CrossRef]
- Mohammadi, M.; Soltanzadeh, M.; Ebrahimi, A.R.; Hamishehkar, H. Spirulina platensis protein hydrolysates: Techno-functional, nutritional and antioxidant properties. Algal Res. 2022, 65, 102739. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J. Appl. Phycol. 2015, 27, 1109–1119. [Google Scholar] [CrossRef]
- Wild, K.J.; Steingaß, H.; Rodehutscord, M. Variability in nutrient composition and in vitro crude protein digestibility of 16 microalgae products. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1306–1319. [Google Scholar] [CrossRef]
- Altmann, B.A.; Neumann, C.; Rothstein, S.; Liebert, F.; Mörlein, D. Do dietary soy alternatives lead to pork quality improvements or drawbacks? A look into micro-alga and insect protein in swine diets. Meat Sci. 2019, 153, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Holman, B.W.B.; Kashani, A.; Malau-Aduli, A.E.O. Effects of Spirulina (Arthrospira platensis) supplementation level and basal diet on liveweight, body conformation and growth traits in genetically divergent Australian dual-purpose lambs during simulated drought and typical pasture grazing. Small Rumin. Res. 2014, 120, 6–14. [Google Scholar] [CrossRef]
- Macias-Sancho, J.; Poersch, L.H.; Bauer, W.; Romano, L.A.; Wasielesky, W.; Tesser, M.B. Fishmeal substitution with Arthrospira (Spirulina platensis) in a practical diet for Litopenaeus vannamei: Effects on growth and immunological parameters. Aquaculture 2014, 426-427, 120–125. [Google Scholar] [CrossRef]
- Grinstead, G.S.; Tokach, M.D.; Dritz, S.S.; Goodband, R.D.; Nelssen, J.L. Effects of Spirulina platensis on growth performance of weanling pigs. Anim. Feed Sci. Technol. 2000, 83, 237–247. [Google Scholar] [CrossRef]
- Radhakrishnan, S.; Belal, I.E.H.; Seenivasan, C.; Muralisankar, T.; Bhavan, P.S. Impact of fishmeal replacement with Arthrospira platensis on growth performance, body composition and digestive enzyme activities of the freshwater prawn, Macrobrachium rosenbergii. Aquac. Rep. 2016, 3, 35–44. [Google Scholar] [CrossRef]
- Dalle Zotte, A.; Cullere, M.; Sartori, A.; Szendrő, Z.; Kovàcs, M.; Giaccone, V.; Dal Bosco, A. Dietary Spirulina (Arthrospira platensis) and Thyme (Thymus vulgaris) supplementation to growing rabbits: Effects on raw and cooked meat quality, nutrient true retention and oxidative stability. Meat Sci. 2014, 98, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Shabana, E.F.; Gabr, M.A.; Moussa, H.R.; El-Shaer, E.A.; Ismaiel, M.M.S. Biochemical composition and antioxidant activities of Arthrospira (Spirulina) platensis in response to gamma irradiation. Food Chem 2017, 214, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Aouir, A.; Amiali, M.; Bitam, A.; Benchabane, A.; Raghavan, V.G. Comparison of the biochemical composition of different Arthrospira platensis strains from Algeria, Chad and the USA. J. Food Meas. Charact. 2017, 11, 913–923. [Google Scholar] [CrossRef]
- Tokuşoglu, Ö.; Üunal, M.K. Biomass nutrient profiles of three microalgae: Spirulina platensis, Chlorella vulgaris, and Isochrisis galbana. J. Food Sci. 2003, 68, 1144–1148. [Google Scholar] [CrossRef]
- Alghonaim, A.A.; Alqahtani, M.F.; Al-Garadi, M.A.; Suliman, G.M.; Al-Baadani, H.H.; Al-Badwi, M.A.; Abdelrahman, M.M.; Alowaimer, A.N.; Khan, R.U.; Alhidary, I.A. Effects of different levels of spirulina (Arthrospira platensis) supplementation on productive performance, nutrient digestibility, blood metabolites, and meat quality of growing Najdi lambs. Trop. Anim. Health Prod. 2022, 54, 124. [Google Scholar] [CrossRef]
- Bennamoun, L.; Afzal, M.T.; Léonard, A. Drying of alga as a source of bioenergy feedstock and food supplement—A review. Renew. Sustain. Energy Rev. 2015, 50, 1203–1212. [Google Scholar] [CrossRef]
- Bensehaila, S.; Doumandji, A.; Boutekrabt, L.; Manafikhi, H.; Peluso, I.; Bensehaila, K.; Kouache, A.; Bensehaila, A. The nutritional quality of Spirulina platensis of Tamenrasset, Algeria. Afr. J. Biotechnol. 2015, 14, 1649–1654. [Google Scholar]
- Carbonell-Capella, J.M.; Buniowska, M.; Barba, F.J.; Esteve, M.J.; Frígola, A. Analytical methods for determining bioavailability and bioaccessibility of bioactive compounds from fruits and vegetables: A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 155–171. [Google Scholar] [CrossRef]
- Cherian, G. A Guide to the Principles of Animal Nutrition; Oregon State University: Corvallis, OR, USA, 2020. [Google Scholar]
- Niazi, S.K. Bioavailability and bioequivalence studies for orally administered drug products. In Handbook of Pharmaceutical Manufacturing Formulations; CRC Press: Boca Raton, FL, USA, 2004; pp. 27–40. [Google Scholar]
- Johnson, R.J. Principles, problems and application of amino acid digestibility in poultry. World’s Poult. Sci. J. 1992, 48, 232–246. [Google Scholar] [CrossRef]
- Sauer, W.C.; Ozimek, L. Digestibility of amino acids in swine: Results and their practical applications: A review. Livest. Prod. Sci. 1986, 15, 367–388. [Google Scholar] [CrossRef]
- Foltyn, M.; Lichovníková, M.; Rada, V.; Musilová, A. Apparent ileal digestibility of protein and amino acids in protein feedstuffs and trypsin activity in the small intestine in broiler chickens. Czech J. Anim. Sci. 2015, 60, 375–382. [Google Scholar] [CrossRef]
- Ravindran, V.; Hew, L.; Ravindran, G.; Bryden, W. A comparison of ileal digesta and excreta analysis for the determination of amino acid digestibility in food ingredients for poultry. Br. Poult. Sci. 1999, 40, 266–274. [Google Scholar] [CrossRef]
- Sibbald, I.R. Estimation of bioavailable amino acids in feedingstuffs for poultry and pigs: A review with emphasis on balance experiments. Can. J. Anim. Sci. 1987, 67, 221–300. [Google Scholar] [CrossRef]
- Parsons, C.M.; Potter, L.M.; Brown, R.D.; Wilkins, T.D.; Bliss, B.A. Microbial contribution to dry matter and amino acid content of poultry excreta. Poult. Sci. 1982, 61, 925–932. [Google Scholar] [CrossRef]
- Mason, V.C.; Just, A. Bacterial activity in the hind-gut of pigs 1. Its influence on the apparent digestibility of dietary energy and fat. Z. Für Tierphysiol. Tierernährung Futterm. 1976, 36, 301–310. [Google Scholar] [CrossRef]
- Jones, J.D.; Petersburg, S.J.; Burnett, P.C. The Mechanism of the Lysine-Arginine Antagonism in the Chick: Effect of lysine on digestion, kidney arginase, and liver transamidinase. J. Nutr. 1967, 93, 103–116. [Google Scholar] [CrossRef]
- Shioji, Y.; Kobayashi, T.; Yoshida, T.; Otagiri, T.; Onoda, K.; Yoshioka, Y.; Sasada, T.; Miyoshi, N. Nitrogen balance and bioavailability of amino acids in Spirulina diet-fed wistar rats. J. Agric. Food Chem. 2021, 69, 13780–13786. [Google Scholar] [CrossRef]
- Niccolai, A.; Chini Zittelli, G.; Rodolfi, L.; Biondi, N.; Tredici, M.R. Microalgae of interest as food source: Biochemical composition and digestibility. Algal Res. 2019, 42, 101617. [Google Scholar] [CrossRef]
- Kose, A.; Ozen, M.O.; Elibol, M.; Oncel, S.S. Investigation of in vitro digestibility of dietary microalga Chlorella vulgaris and cyanobacterium Spirulina platensis as a nutritional supplement. 3 Biotech 2017, 7, 170. [Google Scholar] [CrossRef]
- Lopes, P.A.; Coelho, D.; Prates, J.A.M. Testimony on a successful lab protocol to disrupt Chlorella vulgaris microalga cell wall. PLoS ONE 2022, 17, e0268565. [Google Scholar] [CrossRef]
- Ribeiro, D.M.; Martins, C.F.; Kuleš, J.; Horvatić, A.; Guillemin, N.; Freire, J.P.; Eckersall, P.D.; Almeida, A.M.; Prates, J.A. Influence of dietary Spirulina inclusion and lysozyme supplementation on the longissimus lumborum muscle proteome of newly weaned piglets. J. Proteomics 2021, 244, 104274. [Google Scholar] [CrossRef]
- Bryan, D.D.; Classen, H.L. In vitro Methods of Assessing Protein Quality for Poultry. Animals 2020, 10, 551. [Google Scholar] [CrossRef]
- Neumann, C.; Velten, S.; Liebert, F. N balance studies emphasize the superior protein quality of pig diets at high inclusion level of algae meal (Spirulina platensis) or insect meal (Hermetia illucens) when adequate amino acid supplementation is ensured. Animals 2018, 8, 172. [Google Scholar] [CrossRef]
- Lucakova, S.; Branyikova, I.; Hayes, M. Microalgal Proteins and Bioactives for Food, Feed, and Other Applications. Appl. Sci. 2022, 12, 4402. [Google Scholar] [CrossRef]
- Payne, W.L.; Kifer, R.R.; Snyder, D.G.; Combs, G.F. Studies of protein digestion in the chicken: 1. Investigation of apparent amino acid digestibility of fish meal protein using cecectomized, adult male chickens. Poult. Sci. 1971, 50, 143–150. [Google Scholar] [CrossRef]
- Evans, A.M.; Smith, D.L.; Moritz, J.S. Effects of algae incorporation into broiler starter diet formulations on nutrient digestibility and 3 to 21 d bird performance. J. Appl. Poult. Res. 2015, 24, 206–214. [Google Scholar] [CrossRef]
- Pestana, J.M.; Puerta, B.; Santos, H.; Madeira, M.S.; Alfaia, C.M.; Lopes, P.A.; Pinto, R.M.A.; Lemos, J.P.C.; Fontes, C.M.G.A.; Lordelo, M.M.; et al. Impact of dietary incorporation of Spirulina (Arthrospira platensis) and exogenous enzymes on broiler performance, carcass traits, and meat quality. Poult. Sci. 2020, 99, 2519–2532. [Google Scholar] [CrossRef]
- Chronakis, I.S. Gelation of edible blue-green algae protein isolate (Spirulina platensis Strain Pacifica): Thermal transitions, rheological properties, and molecular forces involved. J. Agric. Food Chem. 2001, 49, 888–898. [Google Scholar] [CrossRef]
- Furbeyre, H.; van Milgen, J.; Mener, T.; Gloaguen, M.; Labussière, E. Effects of dietary supplementation with freshwater microalgae on growth performance, nutrient digestibility and gut health in weaned piglets. Animal 2017, 11, 183–192. [Google Scholar] [CrossRef]
- Linden, J. Starter Pig Amino Acid Requirements to Gut Health Concerns. In Proceedings of the Kansas Swine Profitability Conference, Manhattan, KS, USA, 8 February 2022; p. 1. [Google Scholar]
- Gallo, J.; Pond, W. Tryptophan requirement of early-weaned pigs from three to seven weeks of age. J. Anim. Sci. 1966, 25, 774–778. [Google Scholar] [CrossRef]
- Council, N.R. Nutrient Requirements of Swine; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Altmann, B.A.; Rosenau, S. Spirulina as animal feed: Opportunities and challenges. Foods 2022, 11, 965. [Google Scholar] [CrossRef]
- Martins, C.F.; Pestana Assunção, J.; Ribeiro Santos, D.M.; Madeira, M.; Alfaia, C.; Lopes, P.; Coelho, D.F.M.; Cardoso Lemos, J.P.; de Almeida, A.M.; Mestre Prates, J.A.; et al. Effect of dietary inclusion of Spirulina on production performance, nutrient digestibility and meat quality traits in post-weaning piglets. J. Anim. Physiol. Anim. Nutr. 2021, 105, 247–259. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Nutritional Composition | Arthrospira platensis 1 |
|---|---|
| Crude protein (%) | 26.0–75.6 (61.3) |
| Amino acid profile (% total amino acids) | |
| Alanine | 7.8–10.2 (8.6) |
| Arginine | 7.0–14.9 (9.9) |
| Aspartic acid | 9.8–17.6 (12.4) |
| Cystine | 0.7–2.3 (1.3) |
| Glutamic acid | 14.1–22.5 (17.7) |
| Glycine | 4.9–6.9 (5.6) |
| Histidine | 1.6–3.3 (2.4) |
| Isoleucine | 5.3–12.3 (8.1) |
| Leucine | 9.0–22.4 (14.3) |
| Lysine | 4.4–11.3 (7.8) |
| Methionine | 2.4–6.3 (4.1) |
| Phenylalanine | 4.4–10.5 (7.6) |
| Proline | 3.7 |
| Serine | 5.2–7.1 (5.9) |
| Threonine | 5.0–10.5 (7.4) |
| Tryptophan | 0.85–2.0 (1.4) |
| Tyrosine | 4.0–9.4 (6.5) |
| Valine | 6.2–13.9 (9.1) |
| Ash (%) | 6.1–19.8 (9.0) |
| Macrominerals | |
| Calcium (g/kg) | 0.2–9.2 (4.0) |
| Magnesium (g/kg) | 1.6–4.1 (3.4) |
| Phosphorus (mg/kg) | 1.5–13.9 (8.4) |
| Potassium (g/kg) | 13.7–27.8 (17.6) |
| Sodium (g/kg) | 4.8–27.0 (14.3) |
| Microminerals | |
| Copper (mg/kg) | 1.2–5.1 (3.2) |
| Iron (g/kg) | 0.2–1.1 (0.8) |
| Manganese (mg/kg) | 39.2–54.0 (44.3) |
| Selenium (mg/kg) | 1.1–38.0 (13.5) |
| Zinc (mg/kg) | 25.3–31.1 (27.6) |
| Crude carbohydrates (%) | 4.0–44.8 (17.8) |
| Non-fibre carbohydrates | 7.9–20.9 (15.7) |
| Crude fibre (%) | 0.1–5.0 (2.6) |
| Acid detergent lignin | 0.1–3.2 (1.6) |
| Acid neutral fibre | 0.3–18.3 (6.4) |
| Neutral detergent fibre | 0.2–32.6 (11.0) |
| Crude fat (%) | 0.9–14.2 (5.8) |
| Fatty acid profile (% total fatty acids) | |
| 16:0 | 26.6–71.2 (40.1) |
| 16:1n-7 | 1.8–13.5 (5.3) |
| 18:0 | 0.7–8.8 (2.9) |
| 18:1n-9 | 1.5–35.7 (12.0) |
| 18:2n-6 | 7.9–28.2 (16.8) |
| 18:3n-3 | 0.6–3.0 (1.0) |
| 18:3n-6 | 2.7–28.6 (9.2) |
| 20:0 | 0.02–15.7 (8.1) |
| 20:4n-6 | 0.3–0.4 (0.4) |
| 20:5n-3 | 0.1–2.9 (1.3) |
| 22:6n-3 | 2.3–3.5 (3.0) |
| Pigments (mg/kg) | |
| Total carotenoids | 743–2230 (1450) |
| Total chlorophylls | 1324–3635 (2455) |
| β-carotene | 248–1497 (872) |
| Vitamins (mg/kg) | |
| A | 1.0 |
| B1 | 6.7–44.9 (31.2) |
| B12 | 1.9–17.5 (9.7) |
| B2 | 38.0–56.1 (47.1) |
| B3 | 130–202 (161) |
| B5 | 14.8 |
| B6 | 6.5–12.1 (8.5) |
| B8 | 0.1 |
| B9 | 0.5–500 (250) |
| K | 13.9 |
| α-Tocopherol | 26.2 |
| β-Tocopherol | 1.0 |
| γ-Tocopherol | 1.0 |
| Nutritional Composition | A. platensis Control | A. platensis with Enzyme Treatment |
|---|---|---|
| Total protein | 669 | 586 |
| Total carotenois | 3.04 | 2.73 |
| Total chlorophylls | 6.46 | 8.71 |
| Total fatty acids | 46.7 | 41.8 |
| 16:0 | 41.3 | 41.3 |
| 16:1n-7 | 1.51 | 1.51 |
| 18:0 | 3.10 | 2.74 |
| 18:1n-9 | 2.43 | 2.23 |
| 18:2n-6 | 18.4 | 18.8 |
| 18:3n-3 | 0.090 | 0.106 |
| 18:3n-6 | 24.7 | 24.6 |
| 20:0 | 0.202 | 0.224 |
| 22:2n-6 | 0.066 | 0.103 |
| Pre-Treatment of A. platensis | Main Effects | References |
|---|---|---|
| Combination of pepsin and pancreatin | Increase in organic matter, crude protein and carbohydrate digestibility | Niccolai, et al. [64] |
| Pepsin or pancreatin | Increase in dry weight digestibility (89.6% with pepsin, 97.5% with pancreatin and 94.3% with combination of the two enzymes) | Misurcova, et al. [36] |
| Pancreatin | 64% of hydrolysis yield of microalga biomass | Kose, et al. [65] |
| Enzyme mixture with lysozyme and α-amilase | Release of n-6 PUFA, monounsaturated fatty acids and chlorophyll a | Coelho, et al. [33] |
| Bead milling before in vitro digestion assays | Improvement of protein digestibility (74% for non- disrupted cells vs. 78% for disrupted cells) | Wild, et al. [39] |
| Animals (Age/Initial Body Weight) | Inclusion Level in Feed and Duration of Trial | Main Effects | References |
|---|---|---|---|
| 1-day-old male broilers | 20% with an indigestible marker for 10 days | Enhanced amino acid digestibility | Tavernari, et al. [7] |
| 1-day-old male broilers weighing 41.5 ± 0.5 kg | 1% for 35 days | Increased apparent total tract digestibility of dry matter and nitrogen | Park, et al. [31] |
| 1-day-old male broilers | 15% supplemented with a recombinant lysozyme for 14 days | Nutrient digestibility was not analysed | Pestana, et al. [73] |
| Animals (Age/Initial Body Weight) | Inclusion Level in Feed and Duration of Trial | Main Effects | References |
|---|---|---|---|
| Piglets weaned at 28 days weighing 9.1 ± 1.1 kg | 1% for 14 days | Increased gross energy digestibility coefficient Tended to enhance dry matter, organic matter and neutral detergent fibre digestibility | Furbeyre, et al. [75] |
| Piglets, weaned at 28 days weighing 12.0 ± 0.89kg | 10% for 28 days | Increased total tract apparent digestibility of crude fat and acid detergent fibre (lysozyme supplementation) Decreased total tract apparent digestibility of crude protein (with or without lysozyme) | Martins, et al. [80] |
| 9 week-old growing-finishing pigs weighing 22 ± 1.6 kg | 9.5% with amino acid supplementation, mostly lysine, for 35 to 49 days | Digestibility was not analysed | Altmann, et al. [40] |
| Growing-finishing pigs weighing 60 kg | 13% with amino acid suplementation, mostly lysine. for 10 days | No significant effect on apparent nitrogen digestibility | Neumann, et al. [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spínola, M.P.; Costa, M.M.; Prates, J.A.M. Digestive Constraints of Arthrospira platensis in Poultry and Swine Feeding. Foods 2022, 11, 2984. https://doi.org/10.3390/foods11192984
Spínola MP, Costa MM, Prates JAM. Digestive Constraints of Arthrospira platensis in Poultry and Swine Feeding. Foods. 2022; 11(19):2984. https://doi.org/10.3390/foods11192984
Chicago/Turabian StyleSpínola, Maria P., Mónica M. Costa, and José A. M. Prates. 2022. "Digestive Constraints of Arthrospira platensis in Poultry and Swine Feeding" Foods 11, no. 19: 2984. https://doi.org/10.3390/foods11192984