
Effect of Combined Treatment with Cinnamon Oil and petit-High Pressure CO2 against Saccharomyces cerevisiae


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials and Yeast Strain
2.2. The Minimum Inhibitory Concentration of CIN
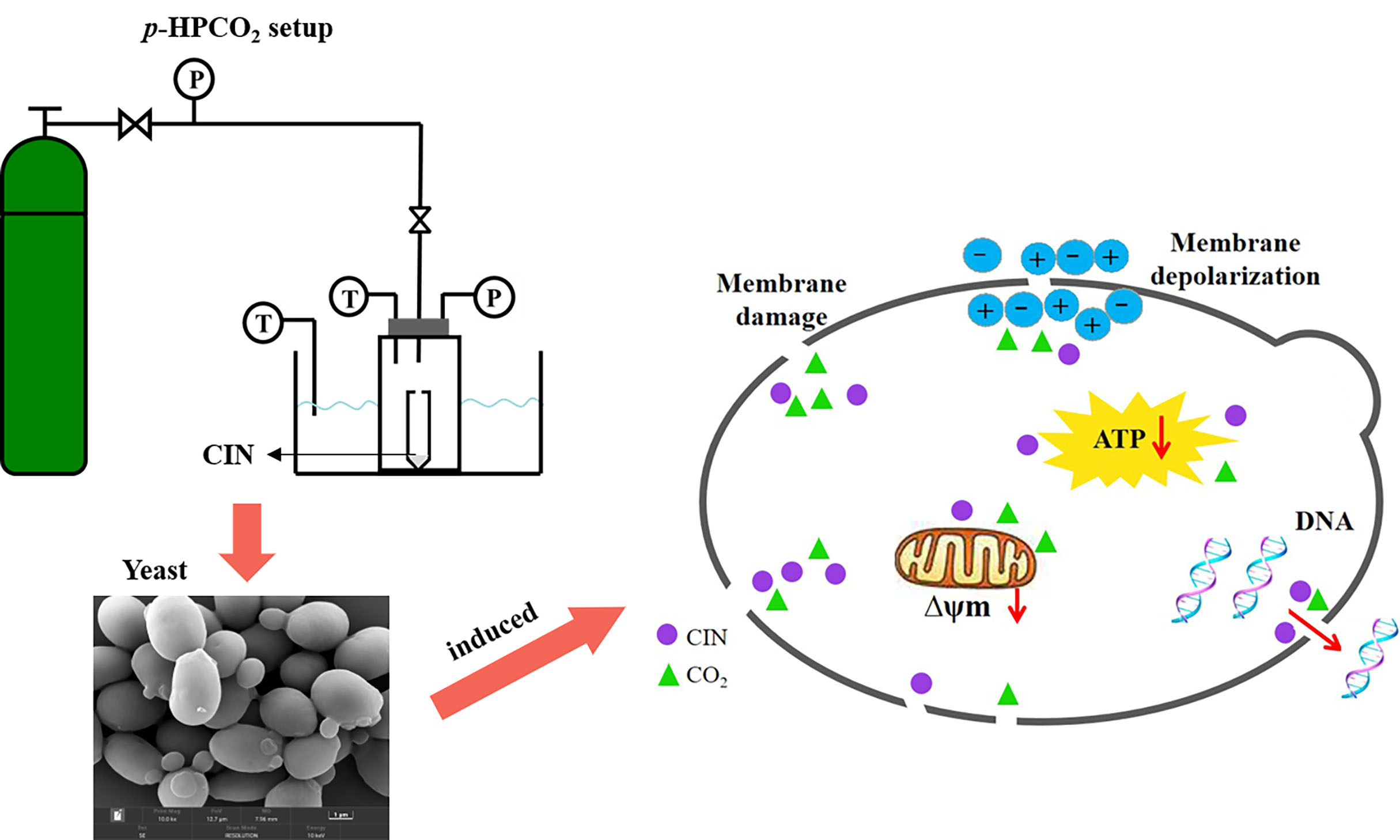
2.3. Treatment with CIN Combined with p-HPCO2
2.4. Synergism Assessment
2.5. Field Emission-Scanning Electron Microscopy
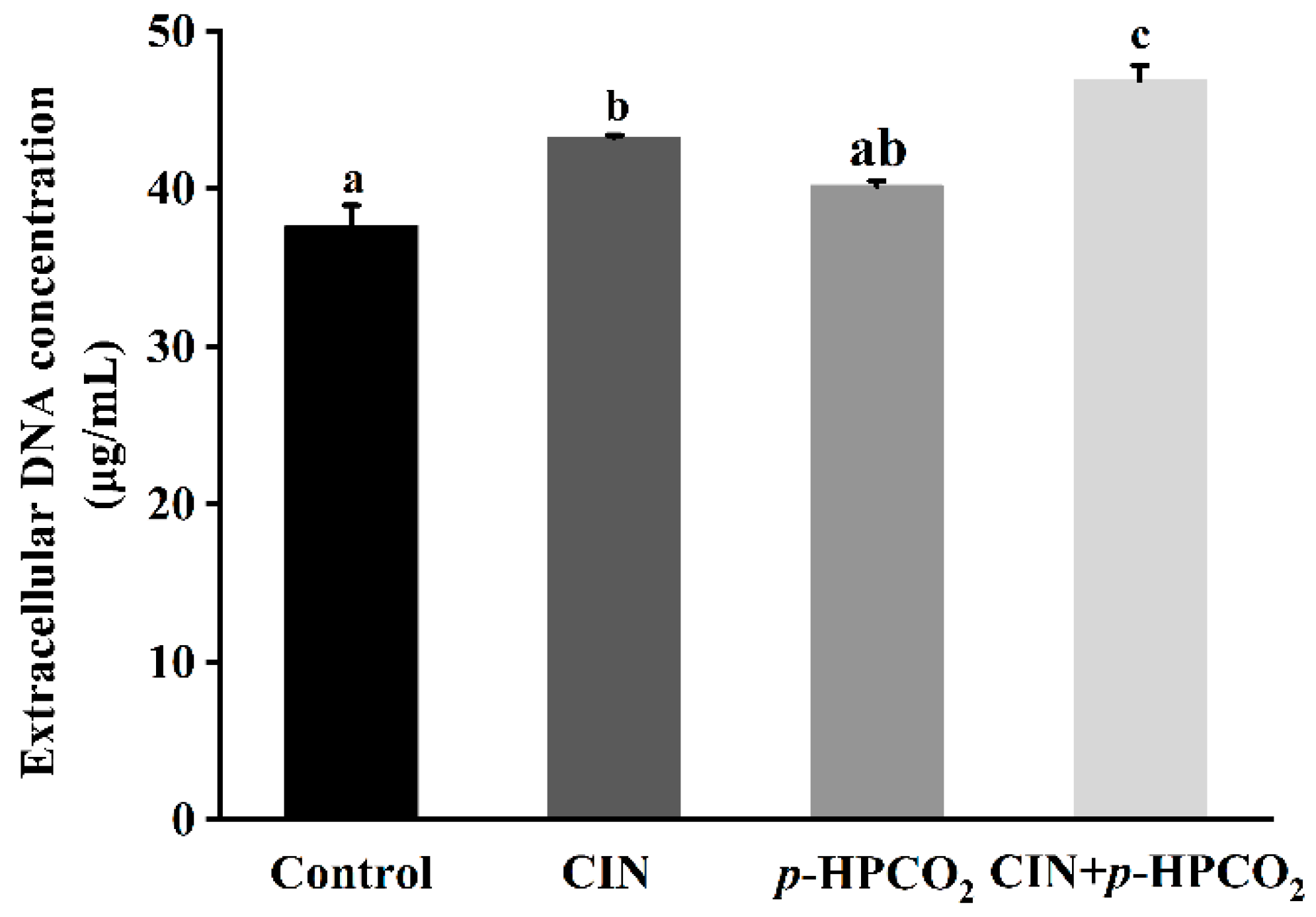
2.6. Leakage of Intracellular DNA
2.7. Cellular Membrane Integrity
2.8. Cellular Membrane Potential
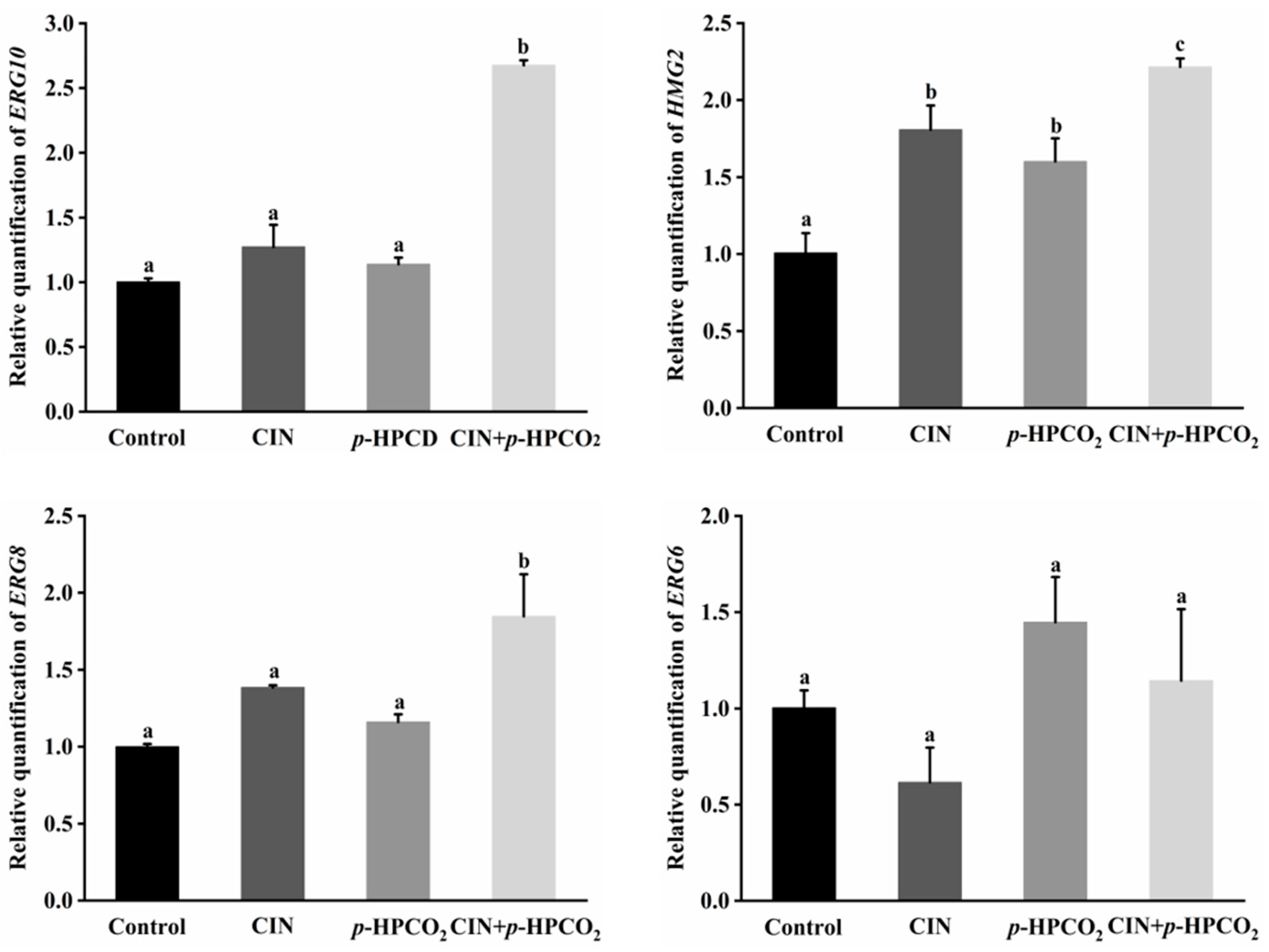
2.9. Expression of the Genes Related to Ergosterol Biosynthesis
2.10. Mitochondrial Membrane Potential
2.11. Intracellular ATP Content
2.12. Statistical Analysis
3. Results and Discussion
3.1. The MIC of CIN
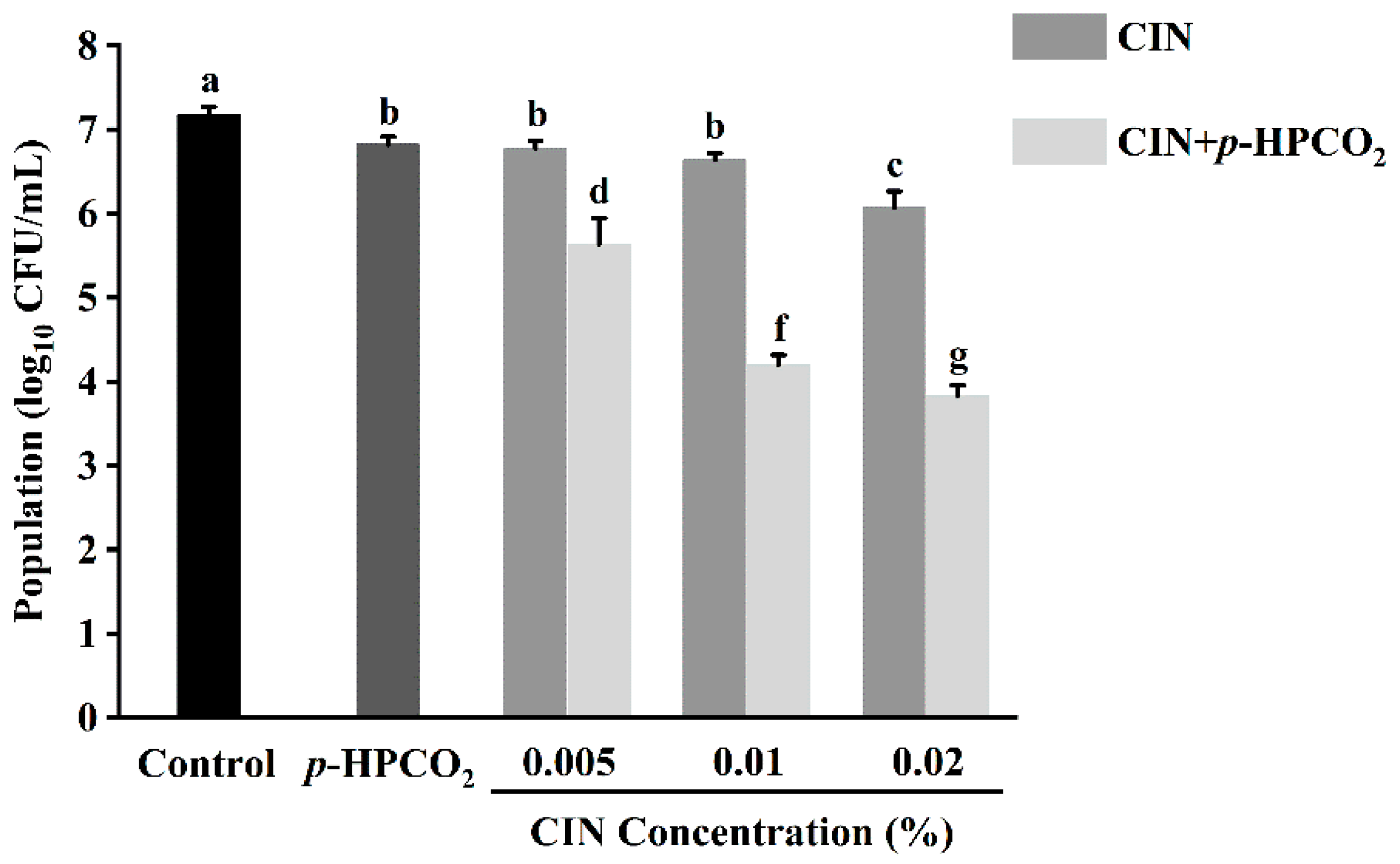
3.2. Antifungal Analysis of CIN + p-HPCO2
3.3. Cell Morphological Characterization
3.4. Cell Membrane Permeability
3.5. Plasma Membrane Integrity
3.6. Cell Membrane Potential
3.7. Expression of Genes Related to Ergosterol Biosynthesis
3.8. Mitochondrial Membrane Potential
3.9. Intracellular ATP
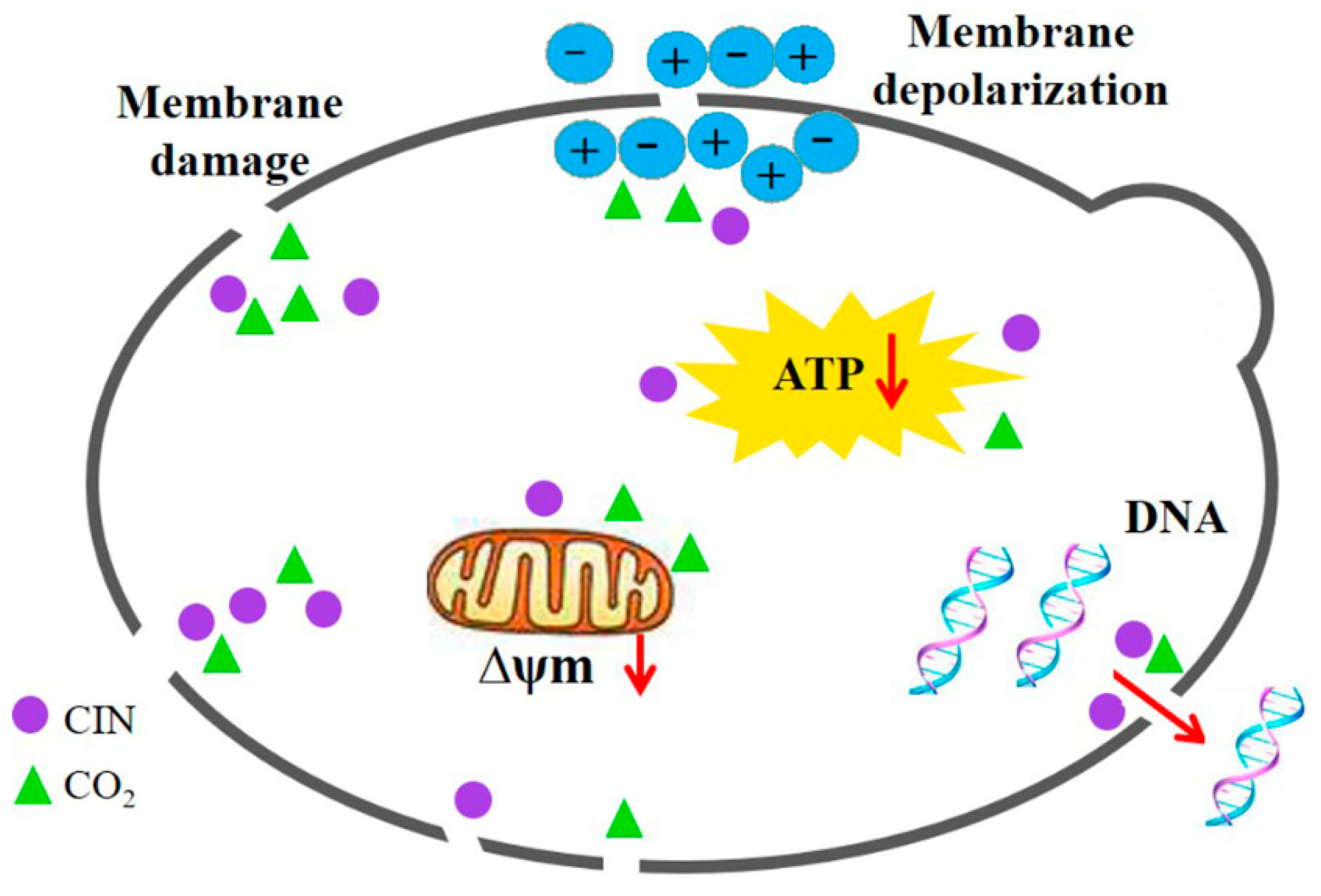
3.10. Synergistic Antifungal Mechanism of CIN + p-HPCO2
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davidson, P.M.; Taylor, T.M.; Schmidt, S.E. Chemical preservatives and natural antimicrobial compounds. In Food Microbiology; Doyle, M.P., Beuchat, L.R., Montville, T.J., Eds.; American Society of Microbiology: Washington, DC, USA, 2013; pp. 765–801. [Google Scholar]
- Mirza, S.K.; Asema, U.; Kasim, S.S. To study the harmful effects of food preservatives on human health. J. Med. Chem. Drug Discovery 2017, 2, 610–616. [Google Scholar]
- Batiha, G.E.; Hussein, D.E.; Algammal, A.M.; George, T.T.; Jeandet, P.; Al-Snafi, A.E.; Tiwari, A.; Pagnossa, J.P.; Lima, C.M.; Thorat, N.D.; et al. Application of natural antimicrobials in food preservation: Recent views. Food Control 2021, 126, 108066. [Google Scholar] [CrossRef]
- Corrales, M.; Fernández, A.; Han, J.H. Antimicrobial Packaging Systems. In Innovations in Food Packaging; Han, J.H., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 133–170. [Google Scholar]
- Ni, Z.J.; Wang, X.; Shen, Y.; Thakur, K.; Han, J.; Zhang, J.G.; Hu, F.; Wei, Z.J. Recent updates on the chemistry, bioactivities, mode of action, and industrial applications of plant essential oils. Trends Food Sci. Tech. 2021, 110, 78–89. [Google Scholar] [CrossRef]
- Chaudhari, A.K.; Singh, A.; Singh, V.K.; Dwivedy, A.K.; Das, S.; Ramsdam, M.G.; Dkhar, M.S.; Kayang, H.; Dubey, N.K. Assessment of chitosan biopolymer encapsulated α-Terpineol against fungal, aflatoxin B1 (AFB1) and free radicals mediated deterioration of stored maize and possible mode of action. Food Chem. 2020, 311, 126010. [Google Scholar] [CrossRef]
- Han, Y.J.; Sun, Z.C.; Chen, W.W. Antimicrobial susceptibility and antibacterial mechanism of limonene against Listeria monocytogenes. Molecules 2020, 25, 33. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Li, C.Z.; Dai, J.M.; Cui, H.Y.; Lin, L. Antibacterial activity and mechanism of Litsea cubeba essential oil against methicillin-resistant Staphylococcus aureus (MRSA). Ind. Crop. Prod. 2019, 130, 34–41. [Google Scholar] [CrossRef]
- Boskovica, M.; Djordjevica, J.; Ivanovica, J.; Janjica, J.; Zdravkovica, N.; Glisica, M.; Glamoclijaa, N.; Balticb, B.; Djordjevicb, V.; Baltic, M. Inhibition of Salmonella by thyme essential oil and its effect on microbiological and sensory properties of minced pork meat packaged under vacuum and modified atmosphere. Int. J. Food Microbiol. 2017, 258, 58–67. [Google Scholar] [CrossRef]
- Pedrós-Garrido, S.; Clemente, I.; Calanche, J.B.; Condón-Abanto, S.; Beltrán, J.A.; Lyng, J.G.; Brunton, N.; Bolton, D.; Whyte, P. Antimicrobial activity of natural compounds against listeria spp. and their effects on sensory attributes in salmon (Salmo salar) and cod (Gadus morhua). Food Control 2020, 107, 106768. [Google Scholar] [CrossRef]
- Sánchez-Rubio, M.; Taboada-Rodríguez, A.; Cava-Roda, R.; López-Molina, D.; Marín-Iniesta, F. Combined use of thermo-ultrasound and cinnamon leaf essential oil to inactivate Saccharomyces cerevisiae in culture broth and natural orange juice. J. Food Sci. Technol. 2018, 55, 4623–4633. [Google Scholar] [CrossRef]
- Martín-Vertedor, D.; Schaide, T.; Boselli, E.; Martínez, M.; García-Parra, J.; Pérez-Nevado, F. Effect of high hydrostatic pressure in the storage of Spanish-style table olive fermented with olive leaf extract and Saccharomyces cerevisiae. Molecules 2022, 27, 2028. [Google Scholar] [CrossRef]
- Lin, L.; Wang, X.L.; Cui, H.Y. Synergistic efficacy of pulsed magnetic fields and Litseacubeba essential oil treatment against Escherichia coli O157:H7 in vegetable juices. Food Control 2019, 106, 106686. [Google Scholar] [CrossRef]
- Paudel, S.K.; Bhargava, K.; Kotturi, H. Antimicrobial activity of cinnamon oil nanoemulsion against Listeria monocytogenes and Salmonella spp. on melons. LWT- Food Sci. Technol. 2019, 111, 682–687. [Google Scholar] [CrossRef]
- Gogoi, R.; Loying, R.; Sarma, N.; Munda, S.; Pandey, S.K.; Lal, M. A comparative study on antioxidant, anti-inflammatory, genotoxicity, anti-microbial activities and chemical composition of fruit and leaf essential oils of Litsea cubeba Pers from North-east India. Ind. Crop. Prod. 2018, 125, 131–139. [Google Scholar] [CrossRef]
- Niu, L.Y.; Nomura, K.; Iwahashi, H.; Matsuoka, H.; Kawachi, S.; Suzuki, Y.; Tamura, K. petit-high Pressure Carbon Dioxide stress increases synthesis of S-Adenosylmethionine and phosphatidylcholine in yeast Saccharomyces cerevisiae. Biophy. Chem. 2017, 231, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.Y.; Zhang, Y.L.; Jie, M.S.; Cheng, Y.X.; Xiang, Q.S.; Zhang, Z.J.; Bai, Y.H. Synergetic effect of petit-high pressure carbon dioxide combined with cinnamon (Cinnamomum cassia) essential oil against Salmonella typhimurium. Int. J. Food Sci. Technol. 2022, 57, 2954–2967. [Google Scholar] [CrossRef]
- Ibrahim, S.Y. Essential oils antagonism against three hygiene significant yeasts and juice spoilage by Saccharomyces cerevisiae. J. Hum. Health Res. 2017, 1, 2576–9383. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, N.; Aliakbarlu, J. Antibacterial effects of ultrasound, cinnamon essential oil, and their combination against Listeria monocytogenes and Salmonella Typhimurium in milk. J. Food Sci. 2019, 84, 3700–3706. [Google Scholar] [CrossRef]
- Niu, L.Y.; Wu, Z.H.; Yang, L.R.; Wang, Y.Q.; Xiang, Q.S.; Bai, Y.H. Antimicrobial Effect of UVC light-emitting diodes against Saccharomyces cerevisiae and their application in orange juice decontamination. J. Food Prot. 2021, 84, 139–146. [Google Scholar] [CrossRef]
- Liu, X.; Li, Y.F.; Zhang, R.; Huangfu, L.L.; Du, G.H.; Xiang, Q.S. Inactivation effects and mechanisms of plasma-activated water combined with sodium laureth sulfate (SLES) against Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2021, 105, 2855–2865. [Google Scholar] [CrossRef]
- Le, V.D.; Tran, V.V.T.; Dang, S.; Nguyen, D.T.; Dang, C.H.; Nguyen, T.D. Physicochemical characterizations, antimicrobial activity and non-isothermal decomposition kinetics of Cinnamomum cassia essential oils. J. Essent. Oil Res. 2020, 32, 158–168. [Google Scholar] [CrossRef]
- Denkova-Kostova, R.; Teneva, D.; Tomova, T.; Goranov, B.; Denkova, Z.; Shopska, V.; Slavchev, A.; Hristova-Ivanova, Y. Chemical composition, antioxidant and antimicrobial activity of essential oils from tangerine (Citrus reticulata L.), grapefruit (Citrus paradisi L.), lemon (Citrus lemon L.) and cinnamon (Cinnamomum zeylanicum Blume). Z. Naturforsch. 2021, 76, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Bang, K.H.; Lee, D.W.; Park, H.M.; Rhee, Y.H. Inhibition of fungal cell wall synthesizing enzymes by trans-cinnamaldehyde. Biosci. Biotechnol. Biochem. 2000, 64, 1061–1063. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gonzalez, L.; Geeraer, A.H.; Spilimbergo, S.; Elst, K.; Ginneken, L.V.; Debevere, J.; Impe, J.F.V.; Devlieghere, F. High pressure carbon dioxide inactivation of microorganisms in foods: The past, the present and the future. Int. J. Food Microbiol. 2007, 117, 1–28. [Google Scholar] [CrossRef]
- Leite, C.; Sousa, J.; Medeiros, J.A.D.C.; Conceicao, M.L.D.; Falcao-Silva, V.D.S.; Souza, E. Inactivation of Escherichia coli, Listeria monocytogenes, and Salmonella Enteritidis by Cymbopogon citratus D.C. Stapf. essential oil in pineapple juice. J. Food Prot. 2016, 79, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Smid, E.J.; Koeken, J.G.P.; Gorris, L.G.M. Fungicidal and fungistatic action of the secondary plant metabolites cinnamaldehyde and carvone. In Modern Fungicides and Antifungal Compounds; Lyr, H., Russell, P.E., Sisler, H.D., Eds.; Intercept Ltd.: Bambous, Mauritius, 1996; pp. 173–180. [Google Scholar]
- Yang, L.; Zhang, C.; Su, Z.; Zhao, L.; Wu, J.; Sun, X.; Zhang, X.; Hu, X. Inactivation of Salmonella typhimurium SL1344 by Chlorogenic Acid and the impairment of cellular integrity. Front. Microbiol. 2022, 13, 887950. [Google Scholar] [CrossRef]
- Ramahi, A.A.; Ruff, R.L. Membrane potential. In Encyclopedia of the Neurological Sciences; Aminoff, M.J., Daroff, R.B., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 1034–1035. [Google Scholar]
- Liu, J.; Zhu, Y.; Du, G.; Zhou, J.; Chen, J. Exogenous ergosterol protects Saccharomyces cerevisiae from d-limonene stress. J. Appl. Microbiol. 2012, 114, 482–491. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, W.; Fan, J.; Jin, C.; Lu, L.; Fang, W. Aspergillus fumigatus mitochondrial acetyl coenzyme A acetyltransferase as an antifungal target. Appl. Environ. Microbiol. 2020, 18, e02986-19. [Google Scholar] [CrossRef] [Green Version]
- Buhaescu, I.; Izzedine, H. Mevalonate pathway: A review of clinical and therapeutical implications. Clin. Biochem. 2007, 40, 575–584. [Google Scholar] [CrossRef]
- Niu, M.; Xiong, Y.; Yan, H.; Zhang, X.; Yuan, L.; Jaime, T.S.; Ma, G. Cloning and expression analysis of mevalonate kinase and phosphomevalonate kinase genes associated with the MVA pathway in Santalum album. Sci. Rep. 2021, 11, 16913. [Google Scholar] [CrossRef]
- Parveen1, M.; Hasan, M.K.; Takahashi, J.; Murata, Y.; Kitagawa, E.; Kodama, O.; Iwahashi, H. Response of Saccharomyces cerevisiae to a monoterpene: Evaluation of antifungal potential by DNA microarray analysis. J. Antimicrob. Chemother. 2004, 54, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Carraro, M.; Bernardi, P. Calcium and reactive oxygen species in regulation of the mitochondrial permeability transition and of programmed cell death in yeast. Cell Calcium 2016, 60, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanovac, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Kim, Y.S.; Shin, D.H. Antimicrobial synergistic effect of linoleic acid and monoglyceride against Bacillus cereus and Staphylococcus aureus. J. Agric. Food Chem. 2002, 50, 2193–2199. [Google Scholar] [CrossRef] [PubMed]
- Amalaradjou, M.A.R.; Venkitanarayanan, K. Effect of trans-cinnamaldehyde on inhibition and inactivation of Cronobacter sakazakii biofilm on abiotic surfaces. J. Food Prot. 2011, 74, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Clément, M.V.; Pervaiz, S. Intracellular superoxide and hydrogen peroxide concentrations: A critical balance that determines survival or death. Redox Rep. 2001, 6, 211–214. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) | Amplicon Size (bp) | |
|---|---|---|---|
| ACT1 | F | ACCGCTGCTCAATCTTCTTC | 164 |
| R | ATGATGGAGTTGTAAGTAGTTTGG | ||
| HMG2 | F | GGTTGGGAAGATATGGAAGTTG | 131 |
| R | ACGACATCACCAGGAATAGTAG | ||
| ERG6 | F | AAGACCTGGCGGACAATGATG | 199 |
| R | AGAGCAGCAGTAACTTCCTTGG | ||
| ERG8 | F | AGTGGCTTCATTCCTGTTTCG | 177 |
| R | TTCGGTAACGCTATCCTCCTG | ||
| ERG10 | F | TCCGCTATGAAGGCAATC | 163 |
| R | CGACACCATCAACAAGAAC | ||
| Observed Log Reduction | Expected Log Reduction | Ratio | Synergism | |
|---|---|---|---|---|
| 0.005% | ||||
| p-HPCO2 | 0.38 | |||
| CIN | 0.40 | |||
| p-HPCO2 + CIN | 1.54 | 0.78 | 1.97 | Yes |
| 0.01% | ||||
| p-HPCO2 | 0.38 | |||
| CIN | 0.54 | |||
| p-HPCO2 + CIN | 2.97 | 0.92 | 3.24 | Yes |
| 0.02% | ||||
| p-HPCO2 | 0.31 | |||
| CIN | 1.11 | |||
| p-HPCO2 + CIN | 3.34 | 1.42 | 2.36 | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, L.; Liu, J.; Wang, X.; Wu, Z.; Xiang, Q.; Bai, Y. Effect of Combined Treatment with Cinnamon Oil and petit-High Pressure CO2 against Saccharomyces cerevisiae. Foods 2022, 11, 3474. https://doi.org/10.3390/foods11213474
Niu L, Liu J, Wang X, Wu Z, Xiang Q, Bai Y. Effect of Combined Treatment with Cinnamon Oil and petit-High Pressure CO2 against Saccharomyces cerevisiae. Foods. 2022; 11(21):3474. https://doi.org/10.3390/foods11213474
Chicago/Turabian StyleNiu, Liyuan, Jingfei Liu, Xinpei Wang, Zihao Wu, Qisen Xiang, and Yanhong Bai. 2022. "Effect of Combined Treatment with Cinnamon Oil and petit-High Pressure CO2 against Saccharomyces cerevisiae" Foods 11, no. 21: 3474. https://doi.org/10.3390/foods11213474
APA StyleNiu, L., Liu, J., Wang, X., Wu, Z., Xiang, Q., & Bai, Y. (2022). Effect of Combined Treatment with Cinnamon Oil and petit-High Pressure CO2 against Saccharomyces cerevisiae. Foods, 11(21), 3474. https://doi.org/10.3390/foods11213474