
Effects on Cell Membrane Integrity of Pichia anomala by the Accumulating Excessive Reactive Oxygen Species under Ethanol Stress
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain
2.2. Culture Conditions
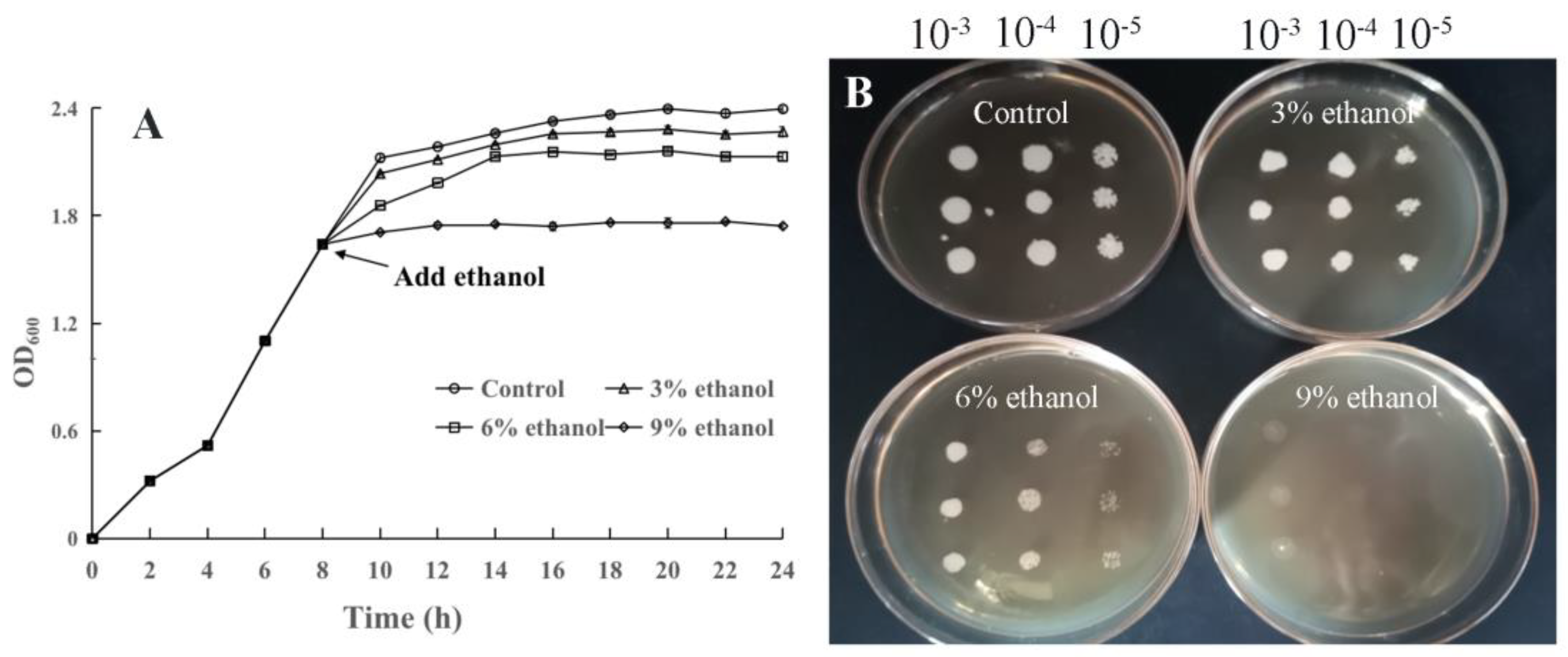
2.3. Determination of Growth of P. anomala NCU003 under Ethanol Stress
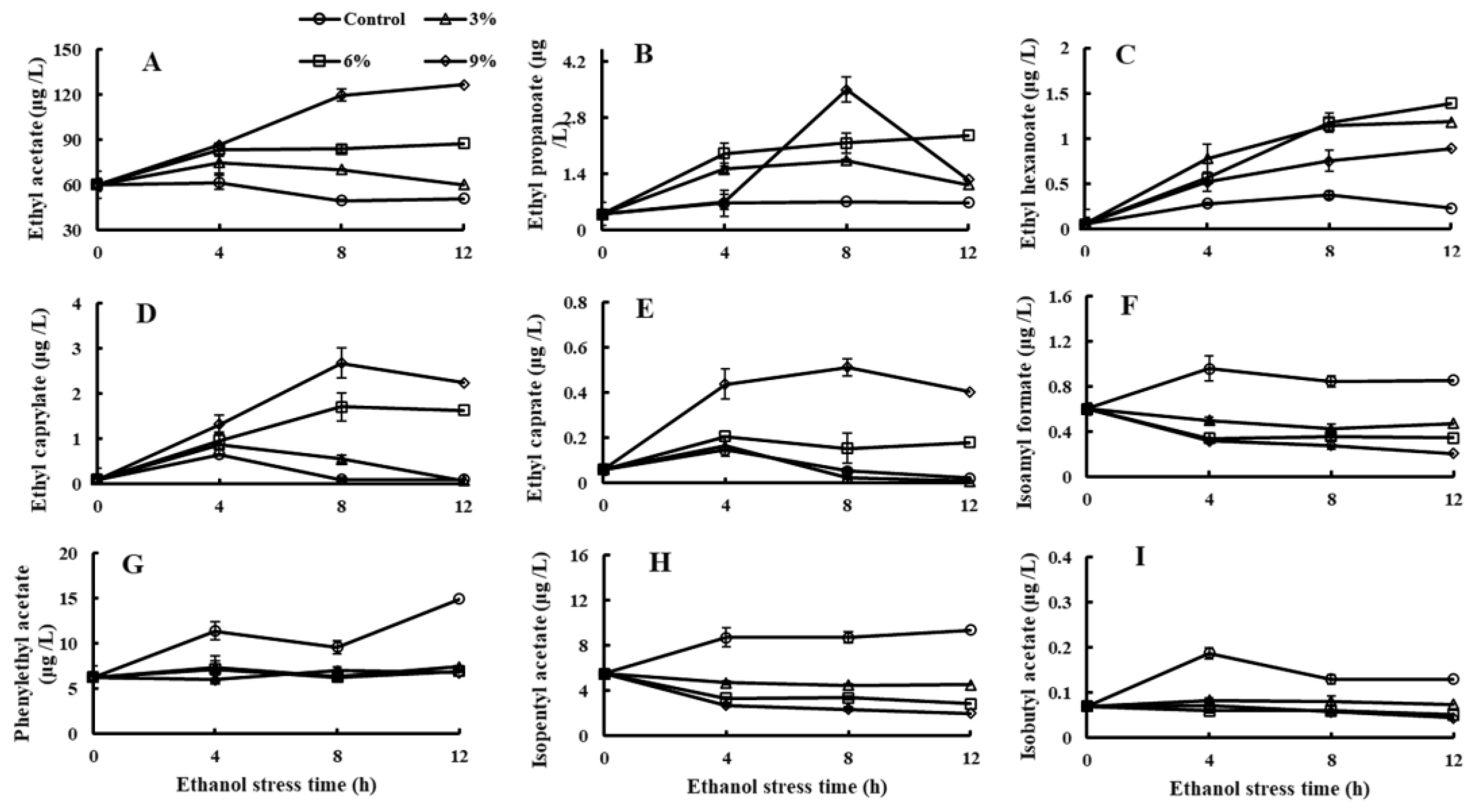
2.4. Determination of Esters Production Ability of P. anomala NCU003 by HS-SPME-GC-MS
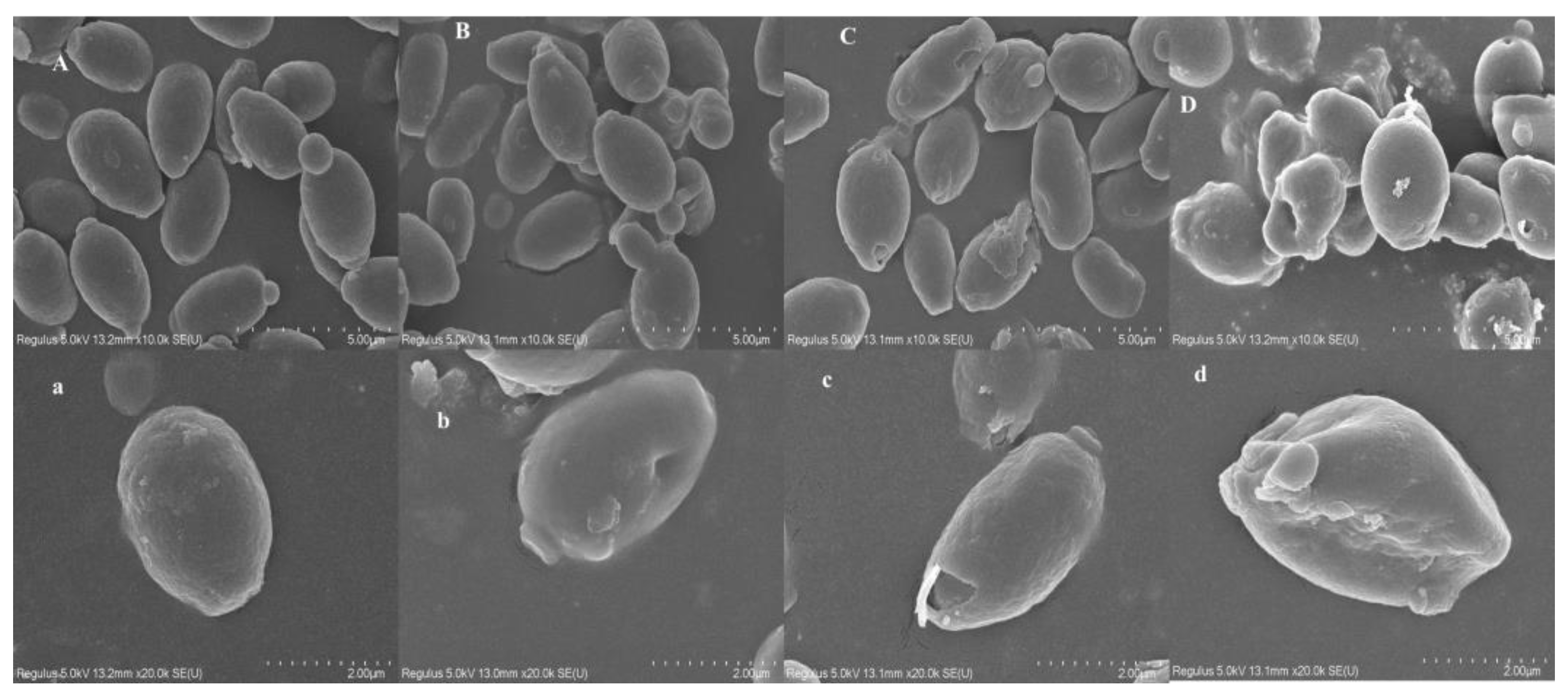
2.5. Determination of Cell Surface Morphology of P. anomala NCU003 with Scanning Electron Microscopy (SEM)
2.6. Determination of Cell Membrane Integrity of P. anomala NCU003 with Propidium Iodide (PI) Staining
2.7. Determination of Intracellular Malondialdehyde (MDA) and Extracellular Leakage Substance Content of P. anomala NCU003
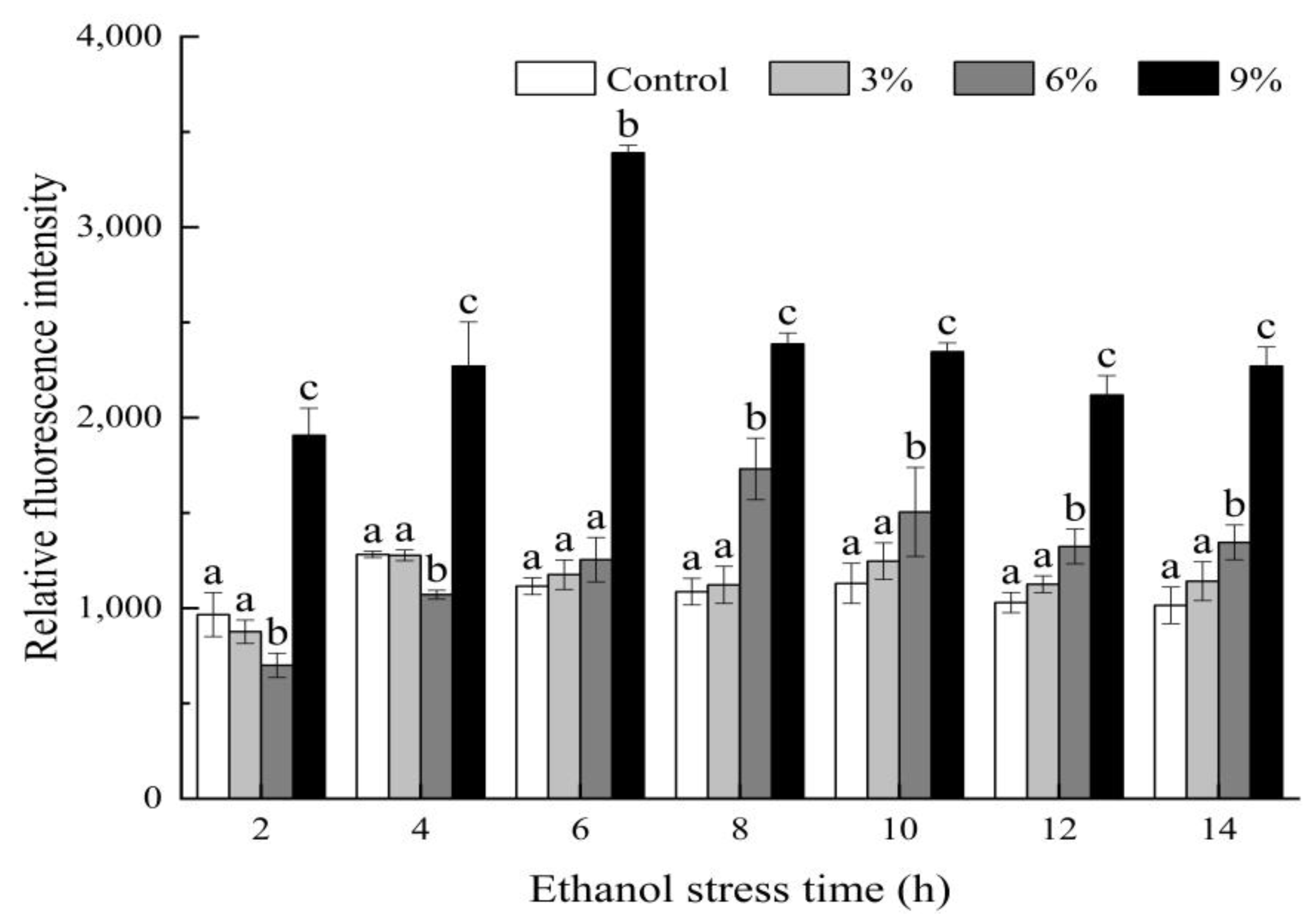
2.8. Determination of ROS Content of P. anomala NCU003
2.9. Determination of Antioxidant Enzyme Activity of P. anomala NCU003
2.10. Data Analysis
3. Results
3.1. Effect of Ethanol Stress on the Growth of P. anomala NCU003
3.2. Effect of Ethanol Stress on the Esters Production Ability of P. anomala NCU003
3.3. Effect of Ethanol Stress on the Cell Morphology of P. anomala NCU003
3.4. Effect of Ethanol Stress on the Cell Membrane Integrity of P. anomala NCU003
3.5. Effect of Ethanol Stress on the MDA and Extracellular Leakage Substance Content of P. anomala NCU003
3.6. Effect of Ethanol Stress on the ROS Content of P. anomala NCU003
3.7. Effect of Ethanol Stress on the Antioxidant Enzymes Activity of P. anomala NCU003
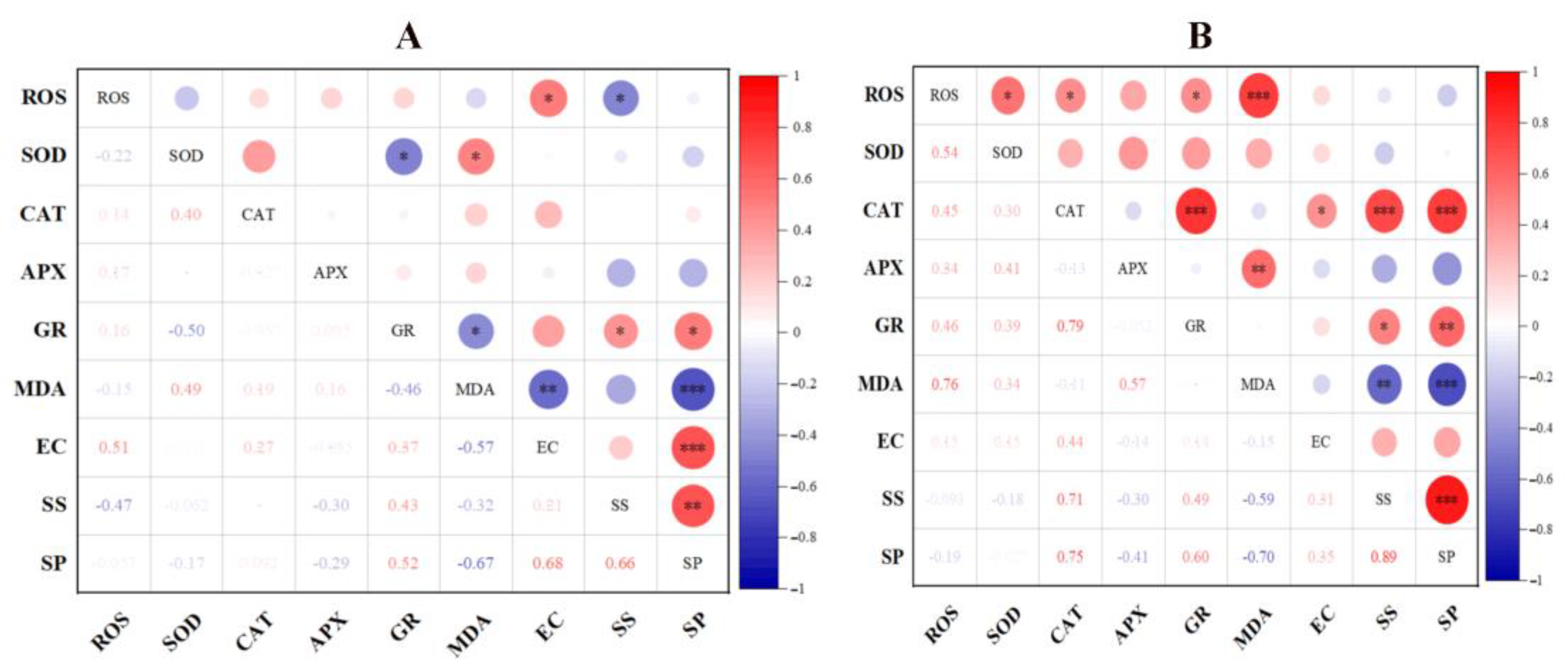
3.8. Correlation Analysis of Cell Membrane Integrity and ROS Metabolism
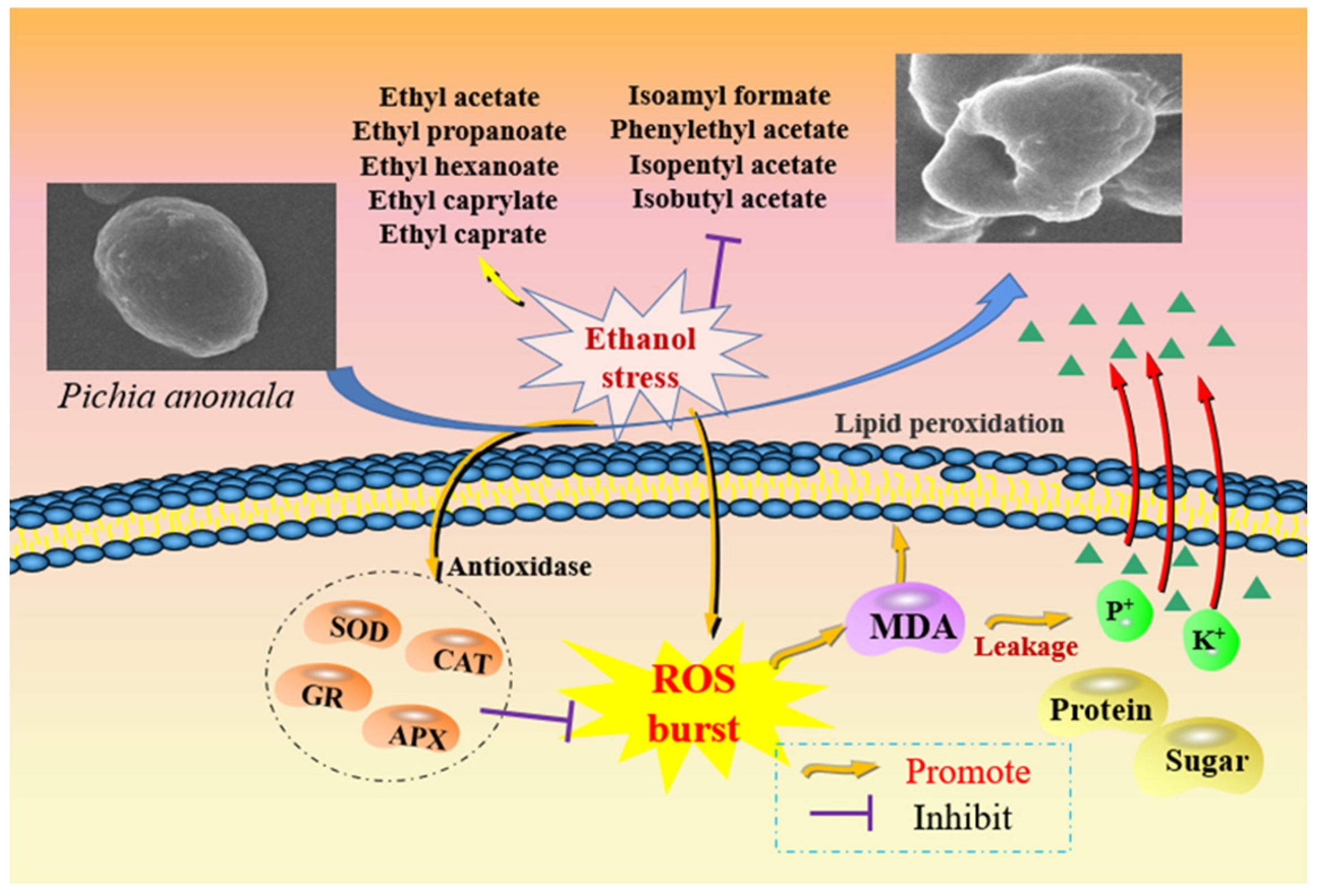
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Li, R.Y.; Miao, Y.J.; Yuan, S.K.; Li, Y.D.; Wu, Z.F.; Weng, P.F. Integrated transcriptomic and proteomic analysis of the ethanol stress response in Saccharomyces cerevisiae Sc131. J. Proteom. 2019, 203, 103377. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Du, H.; Xu, Y. Cooperative response of Pichia kudriavzevii and Saccharomyces cerevisiae to lactic acid stress in Baijiu fermentation. J. Agric. Food Chem. 2020, 68, 4903–4911. [Google Scholar] [CrossRef]
- Liu, C.F.; Li, M.X.; Ren, T.; Wang, J.J.; Niu, C.T.; Zheng, F.Y.; Li, Q. Effect of Saccharomyces cerevisiae and non-Saccharomyces strains on alcoholic fermentation behavior and aroma profile of yellow-fleshed peach wine. LWT-Food Sci. Technol. 2022, 155, 112993. [Google Scholar] [CrossRef]
- Vazquez, J.; Grillitsch, K.; Daum, G.; Mas, A.; Beltran, G.; Torija, M.J. The role of the membrane lipid composition in the oxidative stress tolerance of different wine yeasts. Food Microbiol. 2019, 78, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.C.; Jiang, Y.J.; Liu, J.; Guo, W.L.; Liu, Z.B.; Zhang, W.; Rao, P.F.; Ni, L. Evaluation of different PCR primers for denaturing gradient gel electrophoresis (DGGE) analysis of fungal community structure in traditional fermentation starters used for Hong Qu glutinous rice wine. Int. J. Food Microbiol. 2017, 255, 58–65. [Google Scholar] [CrossRef]
- Larroque, M.N.; Carrau, F.; Faria, L.; Boido, E.; Dellacassa, E.; Medina, K. Effect of Saccharomyces and non-Saccharomyces native yeasts on beer aroma compounds. Int. J. Food Microbiol. 2021, 337, 108953. [Google Scholar] [CrossRef]
- Tang, J.; Wang, H.Y.; Xu, Y. Effect of mixed culture of Saccharomyces cerevisiae and Pichia anomala on fermentation efficiency and flavor compounds in Chinese liquor. Microbiol. China 2012, 39, 921–930, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Challenges of the non-conventional yeast Wickerhamomyces anomalus in winemaking. Fermentation 2018, 4, 68. [Google Scholar] [CrossRef] [Green Version]
- Ianieva, O.; Pidgorskyi, V.S. Enological potential of non-Saccharomyces yeast strains of enological and brewery origin from Ukrainian collection of microorganisms. Mycology 2020, 12, 203–215. [Google Scholar] [CrossRef]
- Li, K.M.; Fu, G.M.; Wu, C.F.; Liu, C.M.; Wan, Y.; Pan, F.; Zheng, F.P. Dynamics of eukaryotic microbial community succession during the traditional fermentation of special-flavor liquor. Food Sci. 2017, 38, 131–136, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Chen, Y.R.; Li, K.M.; Liu, T.; Li, R.Y.; Fu, G.M.; Wan, Y.; Zheng, F.P. Analysis of difference in microbial community and physicochemical indices between surface and central parts of Chinese special-flavor Baijiu Daqu. Front. Microbiol. 2021, 11, 592421. [Google Scholar] [CrossRef]
- Cunha, J.T.; Romaní, A.; Costa, C.E.; Sa-Correia, I.; Domingues, L. Molecular and physiological basis of Saccharomyces cerevisiae tolerance to adverse lignocellulose-based process conditions. Appl. Microbiol. Biotechnol. 2019, 103, 159–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.F.; Yang, H.R.; Coldea, T.E.; Xu, Y.C.; Zhao, H.F. Metabonomic analysis reveals enhanced growth and ethanol production of brewer’s yeast by wheat gluten hydrolysates and potassium supplementation. LWT-Food Sci. Technol. 2021, 145, 111387. [Google Scholar] [CrossRef]
- Duan, L.L.; Shi, Y.; Jiang, R.; Yang, Q.; Wang, Y.Q.; Liu, P.T.; Duan, C.Q.; Yan, G.L. Effects of adding unsaturated fatty acids composition of Saccharomyces cerevisiae and compounds in wine on fatty acid major volatile. S. Afr. J. Enol. Vitic. 2015, 36, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Jing, H.; Liu, H.; Lu, Z.; Gao, J.; Song, H.R.; Tan, X.R. Ethanol induces autophagy regulated by mitochondrial ROS in Saccharomyces cerevisiae. J. Microbiol. Biotechnol. 2018, 28, 1982–1991. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.W.; Wu, Q.; Xu, Y. Specific physiological characteristic of Saccharomyces cerevisiae in Chinese Maotai-flavor liquor making. Microbiol. China 2015, 42, 2098–2107, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Archana, K.M.; Ravi, R.; Anu-Appaiah, K.A. Correlation between ethanol stress and cellular fatty acid composition of alcohol producing non-Saccharomyces in comparison with Saccharomyces cerevisiae by multivariate techniques. J. Food. Sci. Technol. Mys. 2015, 52, 6770–6776. [Google Scholar] [CrossRef] [Green Version]
- Soares, J.R.; de Melo, E.J.T.; da Cunha, M.; Fernandes, K.V.S.; Taveira, G.B.; Pereira, L.D.; Pimenta, S.; Trindade, F.G.; Regente, M.; Pinedo, M. Interaction between the plant ApDef1 defensin and Saccharomyces cerevisiae results in yeast death through a cell cycle-and caspase-dependent process occurring via uncontrolled oxidative stress. BBA-Gen. Subj. 2017, 1861, 3429–3443. [Google Scholar] [CrossRef]
- Mahmud, S.A.; Hirasawa, T.; Shimizu, H. Differential importance of trehalose accumulation in Saccharomyces cerevisiae in response to various environmental stresses. J. Biosci. Bioeng. 2010, 109, 262–266. [Google Scholar] [CrossRef]
- Landolfo, S.; Politi, H.; Angelozzi, D.; Mannazzu, I. ROS accumulation and oxidative damage to cell structures in Saccharomyces cerevisiae wine strains during fermentation of high-sugar-containing medium. BBA-Gen. Subj. 2008, 1780, 892–898. [Google Scholar] [CrossRef]
- Galant, L.S.; Braga, M.M.; de Souza, D.; de Bem, A.F.; Sancineto, L.; Santi, C.; da Rocha, J.B.T. Induction of reactive oxygen species by diphenyl diselenide is preceded by changes in cell morphology and permeability in Saccharomyces cerevisiae. Free Radic. Res. 2017, 51, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, R.; Yamada, R.; Matsumoto, T.; Yoshihara, S.; Tokumoto, H.; Ogino, H. Construction of lactic acid-tolerant Saccharomyces cerevisiae by using CRISPR-Cas-mediated genome evolution for efficient-lactic acid production. Appl. Microbiol. Biotechnol. 2020, 104, 9147–9158. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.M.; Deng, M.F.; Chen, Y.; Chen, Y.R.; Wu, S.W.; Lin, P.; Huang, B.J.; Liu, C.M.; Wan, Y. Analysis of microbial community, physiochemical indices, and volatile compounds of Chinese te-flavor Baijiu Daqu produced in different seasons. J. Sci. Food Agric. 2021, 101, 6525–6532. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, D.; Hoberg, E.; Neugebauer, W.; Tiemann, H.; Darsow, U. Investigation of the boiled potato flavor by human sensory and instrumental methods. Am. J. Potato Res. 2000, 77, 111–117. [Google Scholar] [CrossRef]
- Miao, Y.J.; Xiong, G.T.; Li, R.Y.; Wu, Z.F.; Zhang, X.; Weng, P.F. Transcriptome profiling of Issatchenkia orientalis under ethanol stress. AMB Express 2018, 8, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, Q.R.; Liu, J.H.; Lu, Y.H.; Wang, Y.; Wang, Y.H. Astaxanthin overproduction and proteomic analysis of Phaffia rhodozyma under the oxidative stress induced by TiO2. Bioresour. Technol. 2020, 311, 123525. [Google Scholar] [CrossRef]
- Wei, M.L.; Ge, Y.H.; Li, C.Y.; Chen, Y.R.; Wang, W.H.; Duan, B. Antifungal activity of ε-poly-L-lysine on Trichothecium roseum in vitro and its mechanisms. Physiol. Mol. Plant Pathol. 2018, 103, 23–27. [Google Scholar] [CrossRef]
- Liu, K.W.; Zhou, X.J.; Fu, M.R. Inhibiting effects of epsilon-poly-lysine (ε-PL) on Pencillium digitatum and its involved mechanism. Postharvest Biol. Technol. 2017, 123, 94–101. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Morris, D.L. Quantitative determination of carbohydrates with dreywood’s anthrone reagent. Science 1948, 107, 254–255. [Google Scholar] [CrossRef]
- Ge, Y.H.; Chen, Y.R.; Li, C.Y.; Wei, M.L.; Li, X.; Tang, Q.; Duan, B. Effect of trisodium phosphate treatment on black spot of apple fruit and the roles of anti-oxidative enzymes. Physiol. Mol. Plant Pathol. 2019, 106, 226–231. [Google Scholar] [CrossRef]
- Watanabe, M.; Tamura, K.; Magbanua, J.P.; Takano, K.; Kitamoto, K.; Kitagaki, H.; Akao, T.; Shimoi, H. Elevated expression of genes under the control of stress response element (STRE) and Msn2p in an ethanol-tolerance sake yeast Kyokai no. 11. J. Biosci. Bioeng. 2007, 104, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.J.; Wang, M.Z.; Zhan, R.R.; Yu, Y.; He, Y.G.; Lu, H. Kluyveromyces marxianus developing ethanol tolerance during adaptive evolution with significant improvements of multiple pathways. Biotechnol. Biofuels 2019, 12, 63–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.Q.; Xu, D.D.; Duan, C.Q.; Yan, G.L. Synergistic effect enhances 2-phenylethyl acetate production in the mixed fermentation of Hanseniaspora vineae and Saccharomyces cerevisiae. Process Biochem. 2020, 90, 44–49. [Google Scholar] [CrossRef]
- Arellano-Plaza, M.; Noriega-Cisneros, R.; Clemente-Guerrero, M.; González-Hernández, J.C.; Robles-Herrera, P.D.; Manzo-Ávalos, S.; Saavedra-Molina, A.; Gschaedler-Mathis, A. Fermentative capacity of Kluyveromyces marxianus and Saccharomyces cerevisiae after oxidative stress. J. Inst. Brew. 2017, 123, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Xiao, M.; Zhang, H.; Peng, B. Effects of chlorogenic acid stress on the physiological characteristics of Saccharomyces cerevisiae during the fermentation of cider. J. Chin. Inst. Food Sci. Technol. 2022, 22, 116–124, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Shi, W.Q.; Li, J.; Chen, Y.F.; Liu, X.H.; Chen, Y.F.; Guo, X.W.; Xiao, D.G. Metabolic engineering of Saccharomyces cerevisiae for ethyl acetate biosynthesis. ACS Synth. Biol. 2021, 10, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Cheng, L.; Zhang, X.; Cao, J.X.; Wu, Z.F.; Zheng, X.Z. Transcriptomic and proteomic effects of (-)-epigallocatechin 3-O-(3-O-methyl) gallate (EGCG3″Me) treatment on ethanol-stressed Saccharomyces cerevisiae cells. Food Res. Int. 2019, 119, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Y.R.; Wu, X.J.; Deng, M.F.; Wu, S.W.; Wan, Y.; Liu, C.M.; Fu, G.M. Breeding of excellent ethanol-tolerant Millerozyma farinosa by adaptive evolution. Food Ferment. Ind. 2021, 47, 1–6, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Liu, G.D.; Chen, Y.; Færgeman, N.J.; Nielsen, J. Elimination of the last reactions in ergosterol biosynthesis alters the resistance of Saccharomyces cerevisiae to multiple stresses. FEMS Yeast Res. 2017, 17, 6–13. [Google Scholar] [CrossRef]
- Wang, P.M.; Zheng, D.Q.; Chi, X.Q.; Ou, L.; Wu, X.C. Relationship of trehalose accumulation with ethanol fermentation in industrial Saccharomyces cerevisiae yeast strains. Bioresour. Technol. 2014, 152, 371–376. [Google Scholar] [CrossRef]
- Moravvej, H.; Memariani, H.; Memariani, M.; Kabir-Salmani, M.; Shoae-Hassani, A.; Abdollahimajd, F. Evaluation of fibroblast viability seeded on acellular human amniotic membrane. BioMed Res. Int. 2021, 21, 5597758. [Google Scholar] [CrossRef]
- Talbi, W.; Ghazouani, T.; Braconi, D.; Ben Abdallah, R.; Raboudi, F.; Santucci, A.; Fattouch, S. Effects of selenium on oxidative damage and antioxidant enzymes of eukaryotic cells: Wine Saccharomyces cerevisiae. J. Appl. Microbiol. 2019, 126, 555–566. [Google Scholar] [CrossRef]
- Chen, Y.R.; Ge, Y.H.; Wei, M.L.; Li, C.Y.; Li, X.; Tang, Q.; Duan, B. Inhibition effects of sodium nitroprusside on the Trichothecium roseum in vitro. Food Sci. Technol. 2018, 43, 19–24, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Cheng, Y.F.; Du, Z.L.; Zhu, H.; Guo, X.N.; He, X.P. Protective effects of arginine on Saccharomyces cerevisiae against ethanol stress. Sci. Rep. 2016, 6, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Câmara, A.A., Jr.; Pierre-André Maréchal, P.A.; Tourdot-Maréchal, R.; Husson, F. Oxidative stress resistance during dehydration of three non-Saccharomyces wine yeast strains. Food Res. Int. 2019, 123, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yi, L.H.; Ruan, C.Q.; Yao, S.X.; Deng, L.L.; Zeng, K.F. Proline increases pigment production to improve oxidative stress tolerance and biocontrol ability of Metschnikowia citriensis. Front. Microbiol. 2019, 10, 1273–1282. [Google Scholar] [CrossRef]
- Qiu, Z.; Deng, Z.; Tan, H.; Zhou, S.; Cao, L. Engineering the robustness of Saccharomyces cerevisiae by introducing bifunctional glutathione synthase gene. J. Ind. Microbiol. Biotechnol. 2015, 42, 537–542. [Google Scholar] [CrossRef]
- Kontogianni, V.G.; Tsiafoulis, C.G.; Roussis, I.G.; Gerothanassis, I.P. Selective 1D TOCSY NMR method for the determination of glutathione in white wine. Anal. Methods 2017, 9, 4464–4470. [Google Scholar] [CrossRef]
- Wang, G.Y.; Zhang, T.; Sun, W.; Wang, H.S.; Yin, F.; Wang, Z.Y.; Zuo, D.Q.; Sun, M.X.; Zhou, Z.F.; Lin, B.H.; et al. Arsenic sulfide induces apoptosis and autophagy through the activation of ROS/JNK and suppression of Akt/mTOR signaling pathways in osteosarcoma. Free Radic. Biol. Med. 2017, 106, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, J.Z.; Lu, A.X.; Zhang, K.F.; Li, B.J. Anticancer effect of salidroside on A549 lung cancer cells through inhibition of oxidative stress and phospho-p38 expression. Oncol. Lett. 2014, 7, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Gokce, Z. The protective effect of pistacia vera L. (Pistachio) against to carbon tetrachloride (CCl4)-induced damage in Saccharomyces cerevisiae. Prog. Nutr. 2020, 22, e2020077. [Google Scholar] [CrossRef]
- Fan, G.; Lu, W.; Yao, X.; Zhang, Y.; Wang, K.; Pan, S. Effect of fermentation on free and bound volatile compounds of orange juice. Flavour Fragr. J. 2009, 24, 219–229. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, X.L.; Zhou, G.H. Comparative study of volatile compounds in traditional Chinese Nanjing marinated duck by different extraction techniques. Int. J. Food Sci. Technol. 2007, 42, 543–550. [Google Scholar] [CrossRef]
- Jalali-Heravi, M.; Zekavat, B.; Sereshti, H. Characterization of essential oil components of Iranian geranium oil using gas chromatography-mass spectrometry combined with chemometric resolution techniques. J. Chromatogr. A 2006, 1114, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Pino, J.A.; Mesa, J.; Muñoz, Y.; Martí, M.P.; Marbot, R. Volatile components from mango (Mangifera indica L.) cultivars. J. Agric. Food Chem. 2005, 53, 2213–2223. [Google Scholar] [CrossRef]
- Raffo, A.; Kelderer, M.; Paoletti, F.; Zanella, A. Impact of innovative controlled atmosphere storage technologies and postharvest treatment on volatile compound production in Cv. Pinova apples. J. Agric. Food Chem. 2009, 57, 915–923. [Google Scholar] [CrossRef]
- Pérez, R.A.; Navarro, T.; de Lorenzo, C. HS-SPME analysis of the volatile compounds from spices as a source of flavour in ‘Campo Real’ table olive preparations. Flavour Fragr. J. 2007, 22, 265–273. [Google Scholar] [CrossRef]
- Alissandrakis, E.; Tarantilis, P.A.; Harizanis, P.C.; Polissiou, M. Comparison of the volatile composition in thyme honeys from several origins in Greece. J. Agric. Food Chem. 2007, 55, 8152–8157. [Google Scholar] [CrossRef] [PubMed]
- Javidnia, K.; Miri, R.; Sadeghpour, H. Composition of the volatile oil of Achillea wilhelmsii C. koch from Iran. DARU 2004, 12, 63–66. [Google Scholar]
- Kim, M.R.; Abd El-Aty, A.M.; Kim, I.S.; Shim, J.H. Determination of volatile flavor components in danggui cultivars by solvent free injection and hydrodistillation followed by gas chromatographic-mass spectrometric analysis. J. Chromatogr. A 2006, 1116, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Harzallah-Skhiri, F.; Ben, J.H.; Hammami, S.; Mighri, Z. Variation of volatile compounds in two Prosopis farcta (Banks et Sol.) Eig. (Fabales, Fabaceae = Leguminosae) populations. Flavour Fragr. J. 2006, 21, 484–487. [Google Scholar] [CrossRef]
- Asuming, W.A.; Beauchamp, P.S.; Descalzo, J.T.; Dev, B.C.; Dev, V.; Frost, S.; Ma, C.W. Essential oil composition of four Lomatium Raf. species and their chemotaxonomy. Biochem. Syst. Ecol. 2005, 33, 17–26. [Google Scholar] [CrossRef]
- Mevy, J.P.; Bousquet-Melou, A.; Greff, S.; Millogo, J.; Fernandez, C. Chemical composition of the volatile oil of Laggera aurita Schulz from Burkina-Faso. Biochem. Syst. Ecol. 2006, 34, 815–818. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Stress Time (h) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | ||
| MDA content (μmol/g) | Control | 2.19 ± 0.59 a | 2.97 ± 1.05 a | 3.84 ± 1.66 a | 5.03 ± 0.47 a | 5.17 ± 0.69 a | 4.54 ± 2.05 a | 5.59 ± 0.40 a | 6.73 ± 1.68 a |
| 3% | 2.19 ± 0.59 a | 4.72 ± 0.96 b | 5.63 ± 0.28 b | 5.32 ± 0.91 a | 5.76 ± 0.98 a | 5.65 ± 0.48 a | 7.56 ± 1.26 b | 8.15 ± 0.53 a | |
| 6% | 2.19 ± 0.59 a | 5.02 ± 1.14 b | 9.94 ± 2.10 c | 10.41 ± 3.03 b | 9.43 ± 0.90 b | 7.93 ± 1.94 b | 11.34 ± 2.66 c | 12.84 ± 1.54 b | |
| 9% | 2.19 ± 0.59 a | 21.71 ± 3.07 c | 24.34 ± 4.18 d | 29.16 ± 1.58 c | 28.19 ± 2.64 c | 26.04 ± 2.11 c | 26.16 ± 1.99 d | 26.81 ± 2.12 c | |
| Electrical conductivity (mS/cm) | Control | 25.63 ± 0.86 a | 26.75 ± 0.56 a | 28.23 ± 0.70 a | 28.09 ± 0.47 a | 24.78 ± 0.18 a | 24.58 ± 0.23 a | 24.24 ± 0.27 a | 23.37 ± 0.89 a |
| 3% | 25.63 ± 0.86 a | 28.09 ± 0.24 a | 32.66 ± 1.50 a | 29.72 ± 0.44 a | 28.07 ± 0.65 b | 25.92 ± 0.34 a | 26.25 ± 1.35 a | 27.37 ± 0.74 b | |
| 6% | 25.63 ± 0.86 a | 35.58 ± 1.01 b | 36.08 ± 0.44 a | 33.77 ± 0.43 b | 34.78 ± 0.63 c | 32.49 ± 0.62 b | 35.25 ± 1.55 b | 36.40 ± 1.07 c | |
| 9% | 25.63 ± 0.86 a | 85.95 ± 3.69 c | 83.85 ± 8.29 b | 85.29 ± 1.88 c | 81.66 ± 2.18 d | 79.87 ± 1.17 c | 79.12 ± 2.28 c | 83.92 ± 3.87 d | |
| Soluble protein content (μg/g) | Control | 20.14 ± 0.15 a | 10.10 ± 0.48 a | 8.57 ± 0.08 a | 8.40 ± 0.07 a | 7.34 ± 0.08 a | 7.70 ± 0.13 a | 6.96 ± 0.08 a | 7.38 ± 0.60 a |
| 3% | 20.14 ± 0.15 a | 11.06 ± 0.21 b | 10.28 ± 0.09 b | 9.54 ± 0.16 b | 8.54 ± 0.27 b | 8.01 ± 0.15 a | 8.09 ± 0.21 b | 8.16 ± 0.27 b | |
| 6% | 20.14 ± 0.15 a | 14.05 ± 0.61 c | 13.44 ± 0.15 c | 12.06 ± 0.34 c | 11.35 ± 0.19 c | 10.76 ± 0.05 b | 11.20 ± 0.09 c | 11.28 ± 0.26 c | |
| 9% | 20.14 ± 0.15 a | 35.61 ± 0.24 d | 32.63 ± 0.71 d | 29.00 ± 0.23 d | 27.62 ± 0.78 d | 26.42 ± 0.16 c | 25.68 ± 0.16 d | 24.75 ± 0.62 d | |
| Soluble sugar content (mg/g) | Control | 25.16 ± 0.93 a | 3.68 ± 1.55 a | 0.92 ± 0.17 a | 1.19 ± 0.45 a | 1.31 ± 0.19 a | 1.18 ± 0.60 a | 1.13 ± 0.12 a | 0.88 ± 0.10 a |
| 3% | 25.16 ± 0.93 a | 6.46 ± 0.09 b | 2.04 ± 0.53 a | 2.79 ± 0.86 a | 1.97 ± 0.24 a | 1.30 ± 0.12 a | 2.55 ± 0.21 a | 1.46 ± 0.21 a | |
| 6% | 25.16 ± 0.93 a | 11.00 ± 1.48 c | 10.21 ± 1.32 b | 4.06 ± 0.62 b | 2.57 ± 0.27 b | 3.47 ± 1.65 b | 4.52 ± 0.97 b | 1.97 ± 0.39 a | |
| 9% | 25.16 ± 0.93 a | 35.42 ± 2.87 d | 22.98 ± 1.04 c | 23.90 ± 1.13 c | 20.08 ± 2.52 c | 17.78 ± 2.90 c | 17.96 ± 1.21 c | 10.15 ± 1.62 b | |
| Samples | Stress Time (h) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | ||
| CAT (U/g) | Control | 5.62 ± 0.52 a | 5.55 ± 0.08 a | 5.85 ± 0.40 a | 5.94 ± 0.87 a | 5.89 ± 0.17 a | 4.63 ± 0.23 a | 5.24 ± 0.13 a | 5.42 ± 1.11 a |
| 3% | 5.62 ± 0.52 a | 6.66 ± 0.29 b | 7.27 ± 0.86 b | 7.50 ± 0.48 b | 6.33 ± 0.51 a | 5.48 ± 0.91 a | 5.68 ± 0.19 a | 6.22 ± 1.00 a | |
| 6% | 5.62 ± 0.52 a | 9.30 ± 0.46 c | 8.88 ± 0.29 c | 10.02 ± 0.62 c | 9.64 ± 0.20 b | 9.81 ± 0.47 b | 7.93 ± 0.33 b | 7.76 ± 0.15 b | |
| 9% | 5.62 ± 0.52 a | 25.14 ± 0.16 d | 25.02 ± 0.59 d | 26.71 ± 0.89 d | 22.63 ± 0.61 c | 20.40 ± 0.73 c | 19.37 ± 1.52 c | 19.14 ± 1.07 c | |
| SOD (U/g) | Control | 12.64 ± 0.84 a | 13.24 ± 0.51 a | 12.50 ± 0.09 a | 16.58 ± 0.07 a | 13.46 ± 0.09 a | 12.32 ± 0.16 a | 14.96 ± 0.10 a | 14.26 ± 0.73 a |
| 3% | 12.64 ± 0.84 a | 14.66 ± 0.23 a | 17.37 ± 0.09 b | 18.62 ± 0.16 b | 13.54 ± 0.29 a | 14.76 ± 0.23 ab | 15.47 ± 0.29 a | 16.56 ± 0.35 ab | |
| 6% | 12.64 ± 0.84 a | 16.96 ± 0.68 b | 18.82 ± 0.19 b | 22.82 ± 0.35 c | 16.13 ± 0.23 b | 18.82 ± 0.06 b | 19.78 ± 0.10 b | 19.69 ± 0.40 b | |
| 9% | 12.64 ± 0.84 a | 25.05 ± 0.14 c | 29.68 ± 0.64 c | 33.26 ± 0.20 d | 26.90 ± 0.70 c | 27.22 ± 0.16 c | 24.58 ± 0.16 c | 28.58 ± 0.54 c | |
| APX (U/g) | Control | 0.58 ± 0.14 a | 0.54 ± 0.07 a | 0.61 ± 0.06 a | 0.62 ± 0.15 a | 0.69 ± 0.06 a | 0.61 ± 0.11 a | 0.62 ± 0.09 a | 0.61 ± 0.09 a |
| 3% | 0.58 ± 0.14 a | 0.63 ± 0.10 a | 0.85 ± 0.25 b | 0.83 ± 0.26 b | 0.75 ± 0.08 a | 0.79 ± 0.07 b | 0.63 ± 0.18 a | 0.60 ± 0.06 a | |
| 6% | 0.58 ± 0.14 a | 0.67 ± 0.11 a | 0.96 ± 0.10 b | 1.13 ± 0.23 c | 1.35 ± 0.09 b | 1.19 ± 0.09 c | 0.83 ± 0.24 b | 0.76 ± 0.22 b | |
| 9% | 0.58 ± 0.14 a | 1.73 ± 0.35 b | 1.85 ± 0.03 c | 2.23 ± 0.28 d | 2.43 ± 0.22 c | 2.11 ± 0.27 d | 2.06 ± 0.30 c | 1.99 ± 0.26 c | |
| GR (U/g) | Control | 0.37 ± 0.06 a | 0.40 ± 0.07 a | 0.39 ± 0.06 a | 0.29 ± 0.12 a | 0.37 ± 0.05 a | 0.31 ± 0.03 a | 0.27 ± 0.03 a | 0.33 ± 0.07 a |
| 3% | 0.37 ± 0.06 a | 0.47 ± 0.09 ab | 0.49 ± 0.09 ab | 0.45 ± 0.06 b | 0.35 ± 0.03 a | 0.26 ± 0.02 a | 0.25 ± 0.02 a | 0.32 ± 0.03 a | |
| 6% | 0.37 ± 0.06 a | 0.61 ± 0.09 b | 0.63 ± 0.04 b | 0.48 ± 0.05 bc | 0.44 ± 0.11 b | 0.39 ± 0.04 b | 0.35 ± 0.04 b | 0.37 ± 0.09 a | |
| 9% | 0.37 ± 0.06 a | 0.86 ± 0.11 c | 0.93 ± 0.19 c | 1.01 ± 0.17 c | 0.77 ± 0.18 c | 0.67 ± 0.09 c | 0.60 ± 0.17 c | 0.63 ± 0.08 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Wan, Y.; Cai, W.; Liu, N.; Zeng, J.; Liu, C.; Peng, H.; Fu, G. Effects on Cell Membrane Integrity of Pichia anomala by the Accumulating Excessive Reactive Oxygen Species under Ethanol Stress. Foods 2022, 11, 3744. https://doi.org/10.3390/foods11223744
Chen Y, Wan Y, Cai W, Liu N, Zeng J, Liu C, Peng H, Fu G. Effects on Cell Membrane Integrity of Pichia anomala by the Accumulating Excessive Reactive Oxygen Species under Ethanol Stress. Foods. 2022; 11(22):3744. https://doi.org/10.3390/foods11223744
Chicago/Turabian StyleChen, Yanru, Yin Wan, Wenqin Cai, Na Liu, Jiali Zeng, Chengmei Liu, Hong Peng, and Guiming Fu. 2022. "Effects on Cell Membrane Integrity of Pichia anomala by the Accumulating Excessive Reactive Oxygen Species under Ethanol Stress" Foods 11, no. 22: 3744. https://doi.org/10.3390/foods11223744
APA StyleChen, Y., Wan, Y., Cai, W., Liu, N., Zeng, J., Liu, C., Peng, H., & Fu, G. (2022). Effects on Cell Membrane Integrity of Pichia anomala by the Accumulating Excessive Reactive Oxygen Species under Ethanol Stress. Foods, 11(22), 3744. https://doi.org/10.3390/foods11223744