
Mechanisms of Mitochondrial Apoptosis-Mediated Meat Tenderization Based on Quantitative Phosphoproteomic Analysis
Abstract
:1. Introduction
2. Materials and Methods
2.1. Meat Samples
2.2. Apoptosis Related Factors and Tenderness
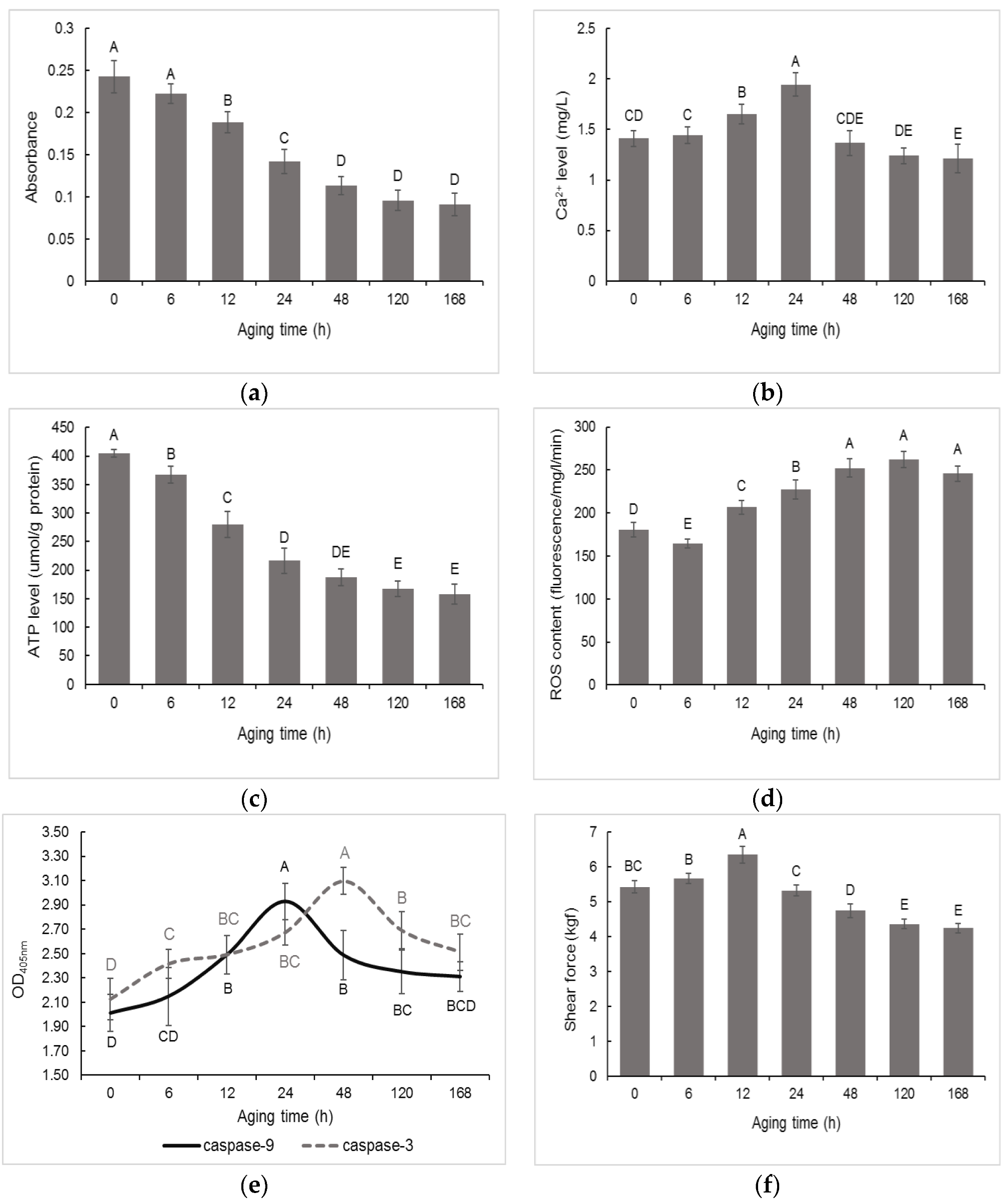
2.2.1. Mitochondrial Membrane Permeability (MMP) and Ca2+ Level
2.2.2. ATP Level
2.2.3. Reactive Oxygen Species (ROS) Content
2.2.4. Caspases Activity
2.2.5. Shear Force
2.3. Protein Extraction and Trypsin Digestion
2.4. Tandem Mass Tags (TMT) Labeling
2.5. High Performance Liquid Chromatography (HPLC) Fractionation
2.6. Phosphopeptide Enrichment and LC-MS/MS Analysis
2.7. Database Search
2.8. Statistical Analysis
3. Results
3.1. Changes in Mitochondrial Apoptosis Related Factors and Tenderness in Postmortem Muscle
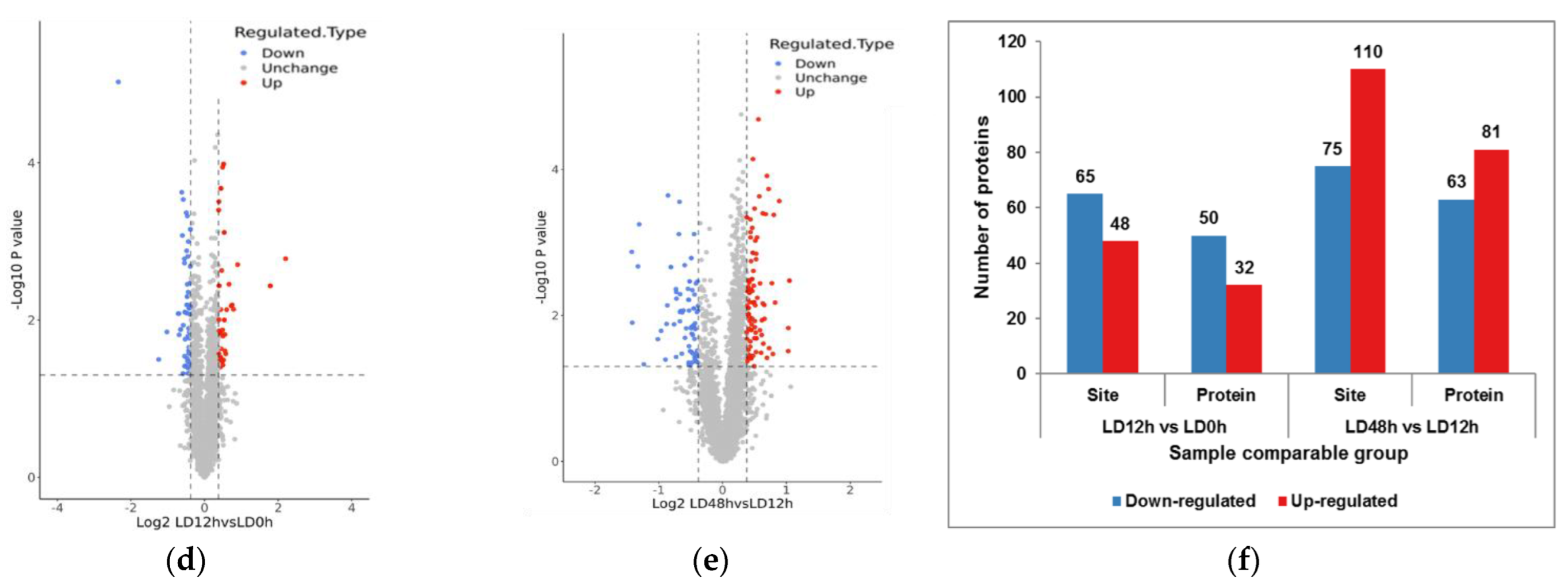
3.2. Identification of Phosphorylated Differentially Expressed Proteins in Postmortem Muscle
3.3. Functional Analysis of Differentially Expressed Proteins in Postmortem Muscle
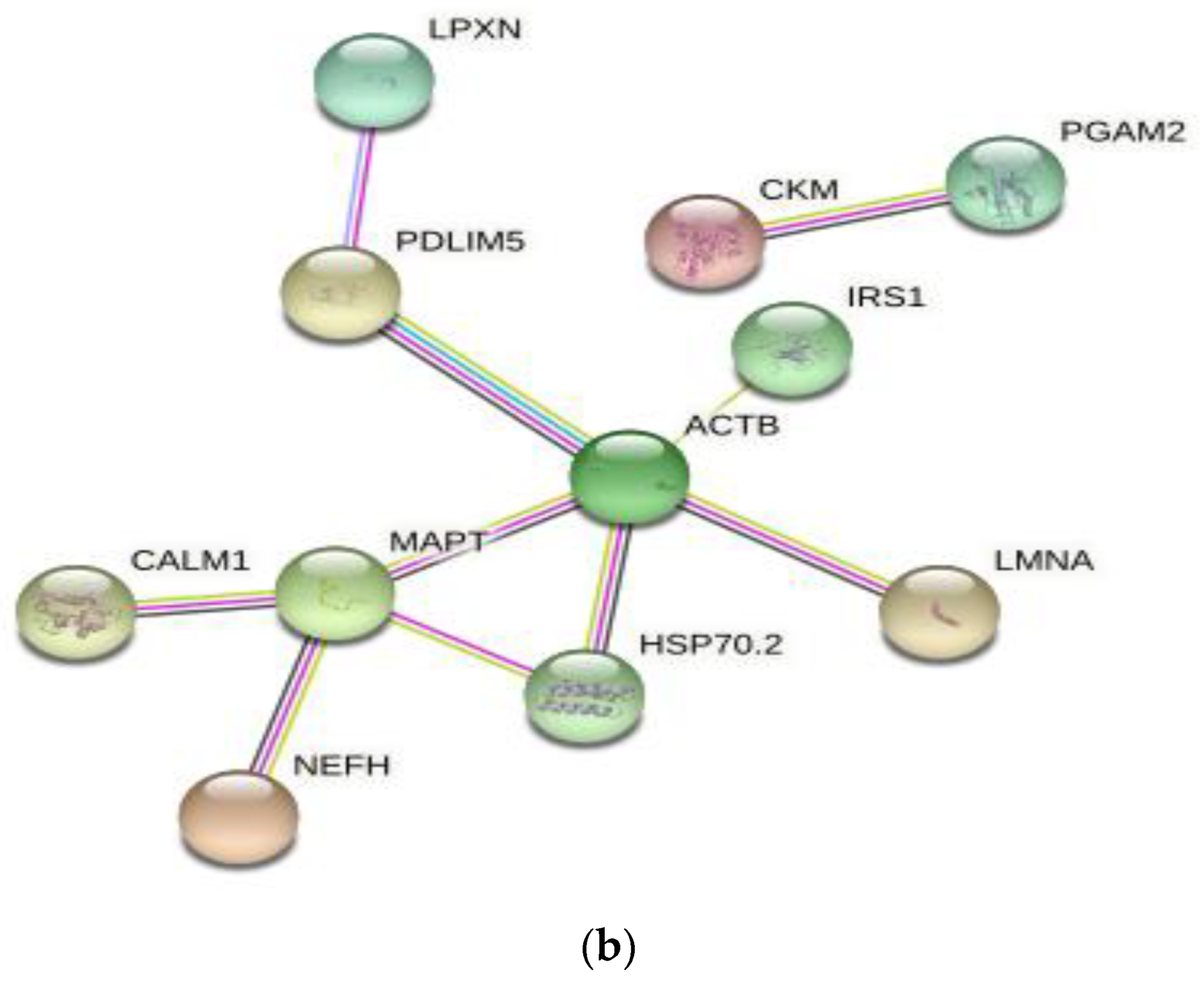
3.4. Differentially Expressed Protein Profiles and Protein-Protein Interaction Network
3.5. Differential Expression of Phosphoproteins in Postmortem Muscle
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ouali, A.; Gagaoua, M.; Boudida, Y.; Becila, S.; Boudjellal, A.; Herrera-Mendez, C.H.; Sentandreu, M.A. Biomarkers of meat tenderness: Present knowledge and perspectives in regards to our current understanding of the mechanisms involved. Meat Sci. 2013, 95, 854–870. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.M.; Sensky, P.L.; Bardsley, R.G.; Buttery, P.J.; Parr, T. Tenderness—An enzymatic view. Meat Sci. 2010, 84, 248–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.B.; Ma, D.; Setyabrata, D.; Farouk, M.M.; Lonergan, S.M.; Huff-Lonergan, E.; Hunt, M.C. Understanding postmortem biochemical processes and post-harvest aging factors to develop novel smart-aging strategies. Meat Sci. 2018, 144, 74–90. [Google Scholar] [CrossRef]
- Huang, F.; Huang, M.; Zhang, H.; Zhang, C.; Zhang, D.; Zhou, G. Changes in apoptotic factors and caspase activation pathways during the postmortem aging of beef muscle. Food Chem. 2016, 190, 110–114. [Google Scholar] [CrossRef]
- Cao, J.-X.; Ou, C.-R.; Zou, Y.-F.; Ye, K.-P.; Zhang, Q.-Q.; Khan, M.A.; Pan, D.-D.; Zhou, G. Activation of caspase-3 and its correlation with shear force in bovine skeletal muscles during postmortem conditioning. J. Anim. Sci. 2013, 91, 4547–4552. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ma, G.; Zhang, Y.; Shi, X.; Han, L.; Yu, Q.; Zhao, S.; Ma, J. Effect of mitochondrial cytochrome c release and its redox state on the mitochondrial-dependent apoptotic cascade reaction and tenderization of yak meat during postmortem aging. Food Res. Int. 2018, 111, 488–497. [Google Scholar] [CrossRef]
- Zhang, J.; Li, M.; Yu, Q.; Han, L.; Ma, Z. Effects of Lysosomal–Mitochondrial Apoptotic Pathway on Tenderness in Post-Mortem Bovine Longissimus Muscle. J. Agric. Food Chem. 2019, 67, 4578–4587. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, Q.; Han, L.; Han, M.; Han, G. Effects of lysosomal iron involvement in the mechanism of mitochondrial apoptosis on postmortem muscle protein degradation. Food Chem. 2020, 328, 127174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, Q.; Han, L.; Chen, C.; Li, H.; Han, G. Study on the apoptosis mediated by cytochrome c and factors that affect the activation of bovine longissimus muscle during postmortem aging. Apoptosis 2017, 22, 777–785. [Google Scholar] [CrossRef]
- Huang, H.; Larsen, M.R.; Palmisano, G.; Dai, J.; Lametsch, R. Quantitative phosphoproteomic analysis of porcine muscle within 24 h postmortem. J. Proteom. 2014, 106, 125–139. [Google Scholar] [CrossRef]
- Du, M.; Li, X.; Li, Z.; Li, M.; Gao, L.; Zhang, D. Phosphorylation inhibits the activity of μ-calpain at different incubation temperatures and Ca2+ concentrations in vitro. Food Chem. 2017, 228, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, X.; Gao, X.; Shen, Q.W.; Du, M.; Zhang, D. Phosphorylation prevents in vitro myofibrillar proteins degradation by μ-calpain. Food Chem. 2017, 218, 455–462. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Zolla, L. Meat science: From proteomics to integrated omics towards system biology. J. Proteom. 2013, 78, 558–577. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, X.; Ni, N.; Liu, Y.; Chen, L.; Wang, Z.; Shen, Q.W.; Zhang, D. Phosphorylation of myofibrillar proteins in post-mortem ovine muscle with different tenderness. J. Sci. Food Agric. 2015, 96, 1474–1483. [Google Scholar] [CrossRef]
- Huang, H.; Larsen, M.R.; Lametsch, R. Changes in phosphorylation of myofibrillar proteins during postmortem development of porcine muscle. Food Chem. 2012, 134, 1999–2006. [Google Scholar] [CrossRef]
- Hunter, T. Signaling—2000 and beyond. Cell 2000, 100, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ma, D.; Kim, Y.H.B. Mitochondrial apoptosis and proteolytic changes of myofibrillar proteins in two different pork muscles during aging. Food Chem. 2020, 319, 126571. [Google Scholar] [CrossRef]
- Wang, L.-L.; Yu, Q.-L.; Han, L.; Ma, X.-L.; Song, R.-D.; Zhao, S.-N.; Zhang, W.-H. Study on the effect of reactive oxygen species-mediated oxidative stress on the activation of mitochondrial apoptosis and the tenderness of yak meat. Food Chem. 2018, 244, 394–402. [Google Scholar] [CrossRef]
- Sun, H.H.; Qiu, N.; Keast, R.; Wang, H.; Li, B.; Huang, Q.; Li, S. Comparative Quantitative Phosphoproteomic Analysis of the Chicken Egg during Incubation Based on Tandem Mass Tag Labeling. J. Agric. Food Chem. 2019, 67, 13353–13361. [Google Scholar] [CrossRef]
- Zhu, C.; Zhu, J.; Duan, Q.; Jiang, Y.; Yin, H.; He, Y.; Li, F.; An, X.-P. Exploration of the lactation function of protein phosphorylation sites in goat mammary tissues by phosphoproteome analysis. BMC Genom. 2021, 22, 703. [Google Scholar] [CrossRef]
- Wang, X. The expanding role of mitochondria in apoptosis. Genes Dev. 2001, 15, 2922–2933. [Google Scholar]
- Armstrong, J.S. Mitochondrial membrane permeabilization: The sine qua non for cell death. BioEssays 2006, 28, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Dillwith, J.W.; Mafi, G.G.; VanOverbeke, D.L.; Ramanathan, R. Metabolite Profile Differences between Beef Longissimus and Psoas Muscles during Display. Meat Muscle Biol. 2017, 1, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Javadov, S.; Karmazyn, M.; Escobales, N. Mitochondrial Permeability Transition Pore Opening as a Promising Therapeutic Target in Cardiac Diseases. J. Pharmacol. Exp. Ther. 2009, 330, 670–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kemp, C.M.; King, D.A.; Shackelford, S.D.; Wheeler, T.L.; Koohmaraie, M. The caspase proteolytic system in callipyge and normal lambs in longissimus, semimembranosus, and infraspinatus muscles during postmortem storage. J. Anim. Sci. 2009, 87, 2943–2951. [Google Scholar] [CrossRef]
- Huang, C.; Hou, C.; Ijaz, M.; Yan, T.; Li, X.; Li, Y.; Zhang, D. Proteomics discovery of protein biomarkers linked to meat quality traits in post-mortem muscles: Current trends and future prospects: A review. Trends Food Sci. Technol. 2020, 105, 416–432. [Google Scholar] [CrossRef]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: The calcium–apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, G.; Guo, Z.; Yu, Q.; Han, L.; Han, M.; Zhu, Y. Study on the apoptosis mediated by apoptosis-inducing-factor and influencing factors of bovine muscle during postmortem aging. Food Chem. 2018, 266, 359–367. [Google Scholar] [CrossRef]
- Sabharwal, S.S.; Schumacker, P.T. Mitochondrial ROS in cancer: Initiators, amplifiers or an Achilles’ heel? Nat. Rev. Cancer 2014, 14, 709–721. [Google Scholar] [CrossRef] [Green Version]
- Lomiwes, D.; Farouk, M.; Wiklund, E.; Young, O. Small heat shock proteins and their role in meat tenderness: A review. Meat Sci. 2014, 96, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-F.; Chen, C.-Y.; Lin, T.-H.; Huang, Z.-W.; Chi, T.-H.; Ma, Y.-S.; Wu, S.-B.; Wei, Y.-H.; Hsieh, M. The protective roles of phosphorylated heat shock protein 27 in human cells harboring myoclonus epilepsy with ragged-red fibers A8344G mtDNA mutation. FEBS J. 2012, 279, 2987–3001. [Google Scholar] [CrossRef]
- Triplett, J.C.; Swomley, A.M.; Cai, J.; Klein, J.B.; Butterfield, D.A. Quantitative Phosphoproteomic Analyses of the Inferior Parietal Lobule from Three Different Pathological Stages of Alzheimer’s Disease. J. Alzheimer’s Dis. 2015, 49, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, M.; Du, M.; Shen, Q.W.; Zhang, D. Dephosphorylation enhances postmortem degradation of myofibrillar proteins. Food Chem. 2018, 245, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Huot, J.; Houle, F.; Spitz, D.; Landry, J. HSP27 phosphorylation-mediated resistance against actin fragmentation and cell death induced by oxidative stress. Cancer Res. 1996, 56, 273–279. [Google Scholar] [PubMed]
- Muroya, S.; Ohnishi-Kameyama, M.; Oe, M.; Nakajima, I.; Shibata, M.; Chikuni, K. Double Phosphorylation of the Myosin Regulatory Light Chain during Rigor Mortis of Bovine Longissimus Muscle. J. Agric. Food Chem. 2007, 55, 3998–4004. [Google Scholar] [CrossRef]
- D’Alessandro, A.; Rinalducci, S.; Marrocco, C.; Zolla, V.; Napolitano, F.; Zolla, L. Love me tender: An Omics window on the bovine meat tenderness network. J. Proteom. 2012, 75, 4360–4380. [Google Scholar] [CrossRef]
- Guillemin, N.; Bonnet, M.; Jurie, C.; Picard, B. Functional analysis of beef tenderness. J. Proteom. 2011, 75, 352–365. [Google Scholar] [CrossRef]
- Chen, L.; Liu, J.; Kaneko, G.; Xie, J.; Wang, G.; Yu, D.; Li, Z.; Ma, L.; Qi, D.; Tian, J.; et al. Quantitative phosphoproteomic analysis of soft and firm grass carp muscle. Food Chem. 2020, 303, 125367. [Google Scholar] [CrossRef]
- Liu, M.; Wei, Y.; Li, X.; Quek, S.Y.; Zhao, J.; Zhong, H.; Zhang, D.; Liu, Y. Quantitative phosphoproteomic analysis of caprine muscle with high and low meat quality. Meat Sci. 2018, 141, 103–111. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Accession | Posit-ion | Amino Acid | Protein Description | Gene Name | Signaling Pathway | LD 0 h | LD 12 h | LD 48 h |
|---|---|---|---|---|---|---|---|---|
| A0A480NRZ3 | 392 | S | Lamin isoform A | LMNA | Regulation of apoptotic signaling pathway | 0.727 | 1.201 | 0.905 |
| A0A5S6I3N7 | 52 | S | Actin, cytoplasmic 1 | ACTB | Apoptosis | 0.144 | 1.023 | 0.495 |
| A0A287B830 | 68 | S | Microtubule-associated protein | MAPT | Apoptotic process | 0.778 | 1.307 | 0.830 |
| Q6S4N2 | 418 | S | Heat shock 70 kDa protein 1B | HSPA1B | Regulation of apoptotic signaling pathway | 0.970 | 1.271 | 0.942 |
| A0A5G2QL63 | 583 | S | Paxillin | PXN | response to reactive oxygen species | 0.898 | 1.316 | 0.860 |
| A0A5G2QTD3 | 112 | T | Calmodulin-1 isoform X1 (Fragment)(predicted) | CALM1 | Calcium signaling pathway | 1.532 | 1.835 | 1.113 |
| I3LLF1 | 1099 | S | Insulin receptor substrate 1 | IRS1 | Aging | 0.827 | 1.132 | 0.863 |
| Q5XLD3 | 140 | Y | Creatine kinase M-type | CKM | ATP metabolic process | 1.250 | 0.855 | 1.158 |
| B5KJG2 | 238 | T | Phosphoglycerate mutase | PGAM2 | ATP metabolic process | 1.654 | 1.208 | 1.575 |
| F1RFH3 | 52 | S | Neurofilament heavy polypeptide | NEFH | Biological regulation | 0.379 | 0.706 | 1.306 |
| A0A287A7G2 | 232 | S | PDZ and LIM domain 5 | PDLIM5 | Cell differentiation | 1.802 | 1.148 | 0.839 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Wang, S.; Ge, W. Mechanisms of Mitochondrial Apoptosis-Mediated Meat Tenderization Based on Quantitative Phosphoproteomic Analysis. Foods 2022, 11, 3751. https://doi.org/10.3390/foods11233751
Zhang J, Wang S, Ge W. Mechanisms of Mitochondrial Apoptosis-Mediated Meat Tenderization Based on Quantitative Phosphoproteomic Analysis. Foods. 2022; 11(23):3751. https://doi.org/10.3390/foods11233751
Chicago/Turabian StyleZhang, Jiaying, Shuangshuang Wang, and Wupeng Ge. 2022. "Mechanisms of Mitochondrial Apoptosis-Mediated Meat Tenderization Based on Quantitative Phosphoproteomic Analysis" Foods 11, no. 23: 3751. https://doi.org/10.3390/foods11233751