
Biochemical Characterization and Effects of Cooking Methods on Main Phytochemicals of Red and Purple Potato Tubers, a Natural Functional Food

 ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Tuber Cultivation and Quality Evaluation
2.2. Cooking Treatments
2.3. Chemicals
2.4. Determination of Total Anthocyanin Content
2.5. Single Anthocyanins and Chlorogenic Acid Analyses
2.6. Ascorbic Acid Content (Vitamin C)
2.7. Chaconine and Solanine Content
2.8. Stock Standard Solution of the Analytes and Calibration Curve
2.9. Statistical Analysis
3. Results and Discussion
3.1. Quantitative Analysis of Chlorogenic Acid in Raw Colored and Yellow Tubers and the Effect of Domestic Cooking Methods
3.2. Anthocyanin Fraction Variability and the Effect of Domestic Cooking Methods
3.3. Quantitative Analysis of Vitamin C in Colored and Yellow Tubers and the Effect of Domestic Cooking Methods
3.4. Quantitative Analysis of Glycoalkaloids in Colored and Yellow Tubers and the Effect of Domestic Cooking Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Graeff-Hönninger, S.; Khajehei, F. The Demand for Superfoods: Consumers’ Desire, Production Viability and Bio-Intelligent Transition. In Food Technology Transitions; Piatti, C., Graeff-Hönninger, S., Khajehei, F., Eds.; Springer: Cham, Switzerland, 2019; pp. 81–94. [Google Scholar]
- Martin, C.; Li, J. Medicine is not health care, food is health care: Plant metabolic engineering, diet and human health. New Phytol. 2017, 216, 699–719. [Google Scholar] [CrossRef] [Green Version]
- Romano, A.; D’Amelia, V.; Gallo, V.; Palomba, S.; Carputo, D.; Masi, P. Relationships between composition, microstructure and cooking performances of six potato varieties. Food Res. Int. 2018, 114, 10–19. [Google Scholar] [CrossRef]
- Mishra, T.; Raigond, P.; Thakur, N.; Dutt, S.; Singh, B. Recent updates on healthy phytoconstituents in potato: A nutritional depository. Potato Res. 2020, 63, 323–343. [Google Scholar] [CrossRef]
- Love, S.L.; Pavek, J.J. Positioning the potato as a primary food source of vitamin C. Am. J. Potato Res. 2008, 85, 277–285. [Google Scholar] [CrossRef]
- Carr, A.C.; Maggini, S. Vitamin C and immune function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef] [Green Version]
- Hellmann, H.; Goyer, A.; Navarre, D.A. Antioxidants in potatoes: A functional view on one of the major food crops worldwide. Molecules 2021, 26, 2446. [Google Scholar] [CrossRef]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of α-glucosidase, α-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, e0191025. [Google Scholar] [CrossRef]
- Chen, K.; Wei, X.; Zhang, J.; Pariyani, R.; Jokioja, J.; Kortesniemi, M.; Linderborg, K.M.; Heinonen, J.; Sainio, T.; Zhang, Y.; et al. Effects of anthocyanin extracts from bilberry (Vaccinium myrtillus L.) and purple potato (Solanum tuberosum L. var. ‘Synkeä Sakari’) on the plasma metabolomic profile of zucker diabetic fatty rats. J. Agric. Food Chem. 2020, 68, 9436–9450. [Google Scholar] [CrossRef]
- De Masi, L.; Bontempo, P.; Rigano, D.; Stiuso, P.; Carafa, V.; Nebbioso, A.; Piacente, S.; Montoro, P.; Aversano, R.; D’Amelia, V.; et al. Comparative phytochemical characterization, genetic profile, and antiproliferative activity of polyphenol-rich extracts from pigmented tubers of different Solanum tuberosum varieties. Molecules 2020, 25, 233. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.L.; Yu, Y.Q.; Chen, Z.J.; Wen, G.S.; Wei, F.G.; Zheng, Q.; Wang, C.D.; Xiao, X.L. Stability-increasing effects of anthocyanin glycosyl acylation. Food Chem. 2017, 214, 119–128. [Google Scholar] [CrossRef]
- Oertel, A.; Matros, A.; Hartmann, A.; Arapitsas, P.; Dehmer, K.J.; Martens, S.; Mock, H.P. Metabolite profiling of red and blue potatoes revealed cultivar and tissue specific patterns for anthocyanins and other polyphenols. Planta 2017, 246, 281–297. [Google Scholar] [CrossRef] [PubMed]
- D’Amelia, V.; Aversano, R.; Chiaiese, P.; Carputo, D. The antioxidant properties of plant flavonoids: Their exploitation by molecular plant breeding. Phytochem. Rev. 2018, 17, 611–625. [Google Scholar] [CrossRef]
- Samaniego, I.; Espin, S.; Cuesta, X.; Arias, V.; Rubio, A.; Llerena, W.; Angós, I.; Carrillo, W. Analysis of environmental conditions effect in the phytochemical composition of potato (Solanum tuberosum) cultivars. Plants 2020, 9, 815. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, Y.; Lai, S.; Cao, H.; Guan, Y.; San Cheang, W.; Liu, B.; Zhao, K.; Miao, S.; Riviere, C.; et al. Effects of domestic cooking process on the chemical and biological properties of dietary phytochemicals. Trends Food Sci. Tech. 2019, 85, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Deligios, P.A.; Chergia, A.P.; Sanna, G.; Solinas, S.; Todde, G.; Narvarte, L.; Ledda, L. Climate change adaptation and water saving by innovative irrigation management applied on open field globe artichoke. Sci. Total Environ. 2019, 649, 461–472. [Google Scholar] [CrossRef]
- Woolfe, J.A. The Potato in the Human Diet; Cambridge University Press: Cambridge, MA, USA, 1987. [Google Scholar]
- Carputo, D.; Frusciante, L.; Monti, L.; Parisi, M.; Barone, A. Tuber quality and soft rot resistance of hybrids between Solarium tuberosum and the incongruent wild relative S. commersonii. Am. J. Potato Res. 2002, 79, 345–352. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and measurement of anthocyanins by UV-visible spectroscopy. CPFAC 2001, 1, F1–F2. [Google Scholar] [CrossRef]
- Sarais, G.; D’Urso, G.; Lai, C.; Pirisi, F.M.; Pizza, C.; Montoro, P. Targeted and untargeted mass spectrometric approaches in discrimination between Myrtus communis cultivars from Sardinia region. J. Mass Spectrom. 2016, 51, 704–715. [Google Scholar] [CrossRef]
- Bodart, P.; Kabengera, C.; Noirfalise, A.; Hubert, P.; Angenot, L. Determination of α-Solanine and α-Chaconine in Potatoes by High-Performance Thin-Layer Chromatography/Densitometry. J. AOAC Int. 2000, 83, 1468–1473. [Google Scholar] [CrossRef] [Green Version]
- Lachman, J.; Hamouz, K.; Šulc, M.; Orsak, M.; Pivec, V.; Hejtmankova, A.; Dvorak, P.; Cepl, J. Cultivar differences of total anthocyanins and anthocyanidins in red and purple-fleshed potatoes and their relation to antioxidant activity. Food Chem. 2009, 114, 836–843. [Google Scholar] [CrossRef]
- Ezekiel, R.; Singh, N.; Sharma, S.; Kaur, A. Beneficial phytochemicals in potato—A review. Food Chem. 2013, 50, 487–496. [Google Scholar] [CrossRef]
- Pazderů, K.; Hamouz, K.; Lachman, J.; Kasal, P. Yield potential and antioxidant activity of potatoes with coloured flesh. Plant. Soil Environ. 2015, 61, 417–442. [Google Scholar] [CrossRef]
- Melito, S.; Garramone, R.; Villano, C.; Carputo, D. Chipping ability, specific gravity and resistance to Pectobacterium carotovorum in advanced potato selections. N. Z. J. Crop. Hort. 2017, 45, 81–90. [Google Scholar] [CrossRef]
- Melito, S.; D’Amelia, V.; Garramone, R.; Villano, C.; Carputo, D. Tuber yield and processing traits of potato advanced selections. Adv. Hortic. Sci. 2017, 31, 151–156. [Google Scholar]
- Brown, C.R.; Vales, I.; Yilma, S.; James, S.; Charlton, B.; Culp, D.; Hane, D.; Shock, C.; Feibert, E.; Pavek, M.; et al. “AmaRosa”, a red skinned, red fleshed fingerling with high phytonutrient value. Am. J. Potato Res. 2012, 89, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Ingallina, C.; Spano, M.; Sobolev, A.P.; Esposito, C.; Santarcangelo, C.; Baldi, A.; Daglia, M.; Mannina, L. Characterization of Local Products for Their Industrial Use: The Case of Italian Potato Cultivars Analyzed by Untargeted and Targeted Methodologies. Foods. 2020, 9, 1216. [Google Scholar] [CrossRef] [PubMed]
- Payyavula, R.S.; Shakya, R.; Sengoda, V.G.; Munyaneza, J.E.; Swamy, P.; Navarre, D.A. Synthesis and regulation of chlorogenic acid in potato: Rerouting phenylpropanoid flux in HQT-silenced lines. Plant. Biotech. J. 2015, 13, 551–564. [Google Scholar] [CrossRef]
- Rommens, C.M.; Richael, C.M.; Yan, H.; Navarre, D.A.; Ye, J.; Krucker, M.; Swords, K. Engineered native pathways for high kaempferol and caffeoylquinate production in potato. Plant. Biotechnol. J. 2008, 6, 870–886. [Google Scholar] [CrossRef]
- De Andrade Lima, M.; Andreou, R.; Charalampopoulos, D.; Chatzifragkou, A. Supercritical carbon dioxide extraction of phenolic compounds from potato (Solanum tuberosum) peels. Appl. Sci. 2021, 11, 3410. [Google Scholar] [CrossRef]
- Valiñas, M.A.; Lanteri, M.L.; Ten Have, A.; Andreu, A.B. Chlorogenic acid, anthocyanin and flavan-3-ol biosynthesis in flesh and skin of Andean potato tubers (Solanum tuberosum subsp. andigena). Food Chem. 2017, 229, 837–846. [Google Scholar] [CrossRef]
- Damiano, S.; Iovane, V.; Squillacioti, C.; Mirabella, N.; Prisco, F.; Ariano, A.; Amenta, M.; Giordano, A.; Florio, S.; Ciarcia, R. Red orange and lemon extract prevents the renal toxicity induced by ochratoxin A in rats. J. Cell Physiol. 2020, 235, 5386–5393. [Google Scholar] [CrossRef] [PubMed]
- Antonić, B.; Jančíková, S.; Dordević, D.; Tremlová, B. Grape pomace valorization: A systematic review and meta-analysis. Foods 2020, 9, 1627. [Google Scholar] [CrossRef] [PubMed]
- Karimi, A.; Kazemi, M.; Samani, S.A.; Simal-Gandara, J. Bioactive compounds from by-products of eggplant: Functional properties, potential applications and advances in valorization methods. Trends Food. Sci. Tech. 2021, 112, 518–531. [Google Scholar] [CrossRef]
- Gebrechristos, H.Y.; Chen, W. Utilization of potato peel as eco-friendly products: A review. Food Sci. Nutr. 2018, 6, 1352–1356. [Google Scholar] [CrossRef] [Green Version]
- Sampaio, S.L.; Petropoulos, S.A.; Alexopoulos, A.; Heleno, S.A.; Santos-Buelga, C.; Barros, L.; Ferreira, I.C. Potato peels as sources of functional compounds for the food industry: A review. Trends Food Sci. Technol. 2020, 103, 118–129. [Google Scholar] [CrossRef]
- Oliveira Filho, J.G.; Braga, A.R.C.; de Oliveira, B.R.; Gomes, F.P.; Moreira, V.L.; Pereira, V.A.C.; Egea, M.B. The potential of anthocyanins in smart, active, and bioactive eco-friendly polymer-based films: A review. Food Res. Int. 2021, 142. [Google Scholar] [CrossRef]
- Furrer, A.N.; Chegeni, M.; Ferruzzi, M.G. Impact of potato processing on nutrients, phytochemicals, and human health. Crit. Rev. Food. Sci. Nutr. 2018, 58, 146–168. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Lachman, J.; Hamouz, K.; Musilova, J.; Hejtmankova, K.; Kotikova, Z.; Pazderu, K.; Domkarova, J.; Pivec, V.; Cimr, J. Effect of peeling and three cooking methods on the content of selected phytochemicals in potato tubers with various colour of flesh. Food Chem. 2013, 138, 1189–1197. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marceli, L.F.; Visser, R.G.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef]
- Hugueney, P.; Provenzano, S.; Verriès, C.; Ferrandino, A.; Meudec, E.; Batelli, G.; Merdinoglu, D.; Cheynier, V.; Schubert, A.; Ageorges, A. A novel cation-dependent O-methyltransferase involved in anthocyanin methylation in grapevine. Plant. Physiol. 2009, 150, 2057–2070. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.S.; Griffiths, H.M.; De Jong, D.M.; Cheng, S.; Bodis, M.; Kim, T.S.; De Jong, W.S. The potato developer (D) locus encodes an R2R3 MYB transcription factor that regulates expression of multiple anthocyanin structural genes in tuber skin. Theor. Appl. Genet. 2009, 120, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovinich, N.; Kayanja, G.; Chanoca, A.; Riedl, K.; Otegui, M.S.; Grotewold, E. Not all anthocyanins are born equal: Distinct patterns induced by stress in Arabidopsis. Planta 2014, 240, 931–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachman, J.; Hamouz, K.; Orsák, M.; Pivec, V.; Hejtmánková, K.; Pazderů, K.; Dvořák, P.; Čepl, J. Impact of selected factors–Cultivar, storage, cooking and baking on the content of anthocyanins in coloured-flesh potatoes. Food Chem. 2012, 133, 1107–1116. [Google Scholar] [CrossRef]
- Lemos, M.A.; Aliyu, M.M.; Hungerford, G. Influence of cooking on the levels of bioactive compounds in purple majesty potato observed via chemical and spectroscopic means. Food Chem. 2015, 173, 462–467. [Google Scholar] [CrossRef]
- Rowe, S.; Carr, A.C. Global vitamin C status and prevalence of deficiency: A cause for concern? Nutrients 2020, 12, 2008. [Google Scholar] [CrossRef]
- Külen, O.; Stushnoff, C.; Holm, D.G. Effect of cold storage on total phenolics content, antioxidant activity and vitamin C level of selected potato clones. J. Sci. Food Agric. 2013, 93, 2437–2444. [Google Scholar] [CrossRef]
- Grudzińska, M.; Czerko, Z.; Zarzyńska, K.; Komenda, M.B. Bioactive compounds in potato tubers: Effects of farming system, cooking method, and flesh color. PLoS ONE 2016, 3, e0153980. [Google Scholar] [CrossRef] [Green Version]
- Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.R.; Leblanc, J.-C.; Nebbia, C.S.; et al. Risk assessment of glycoalkaloids in feed and food, in particular in potatoes and potato derived products. EFSA J. 2020, 18, 6222. [Google Scholar] [CrossRef]
- Valcarcel, J.; Reilly, K.; Gaffney, M.; O’Brien, N. Effect of genotype and environment on the glycoalkaloid content of rare, heritage, and commercial potato varieties. J. Food Sci. 2014, 79, T1039–T1048. [Google Scholar] [CrossRef] [Green Version]
- Musita, C.N.; Okoth, M.W.; Abong, G.O.; Omayio, D.G. Glycoalkaloids in commercial potato varieties traded in Nairobi, Kenya. F1000 Res. 2020, 9, 423. [Google Scholar] [CrossRef]
- Friedman, M.; Huang, V.; Quiambao, Q.; Noritake, S.; Liu, J.; Kwon, O.; Chintalapati, S.; Young, J.; Levin, C.E.; Tam, C.; et al. Potato peels and their bioactive glycoalkaloids and phenolic compounds inhibit the growth of pathogenic trichomonads. J. Agric. Food Chem. 2018, 66, 7942–7947. [Google Scholar] [CrossRef]
- Nie, X.; Zhang, G.; Lv, S.; Guo, H. Steroidal glycoalkaloids in potato foods as affected by cooking methods. Int. J. Food Prop. 2018, 21, 1875–1887. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | TY | TSG | CC |
|---|---|---|---|
| Musica | 0.59 ± 0.13 a | 1.069 ± 0 | 4.00 ± 0.58 b |
| Magenta Love | 0.14 ± 0.08 b | 1.078 ± 0 | 2.00 ± 0 c |
| Scano di Montiferro | 0.20 ± 0.01 b | 1.095 ± 0 | 7.00 ± 0.58 a |
| Violet Queen | 0.21 ± 0.04 b | 1.079 ± 0 | 4.00 ± 0 b |
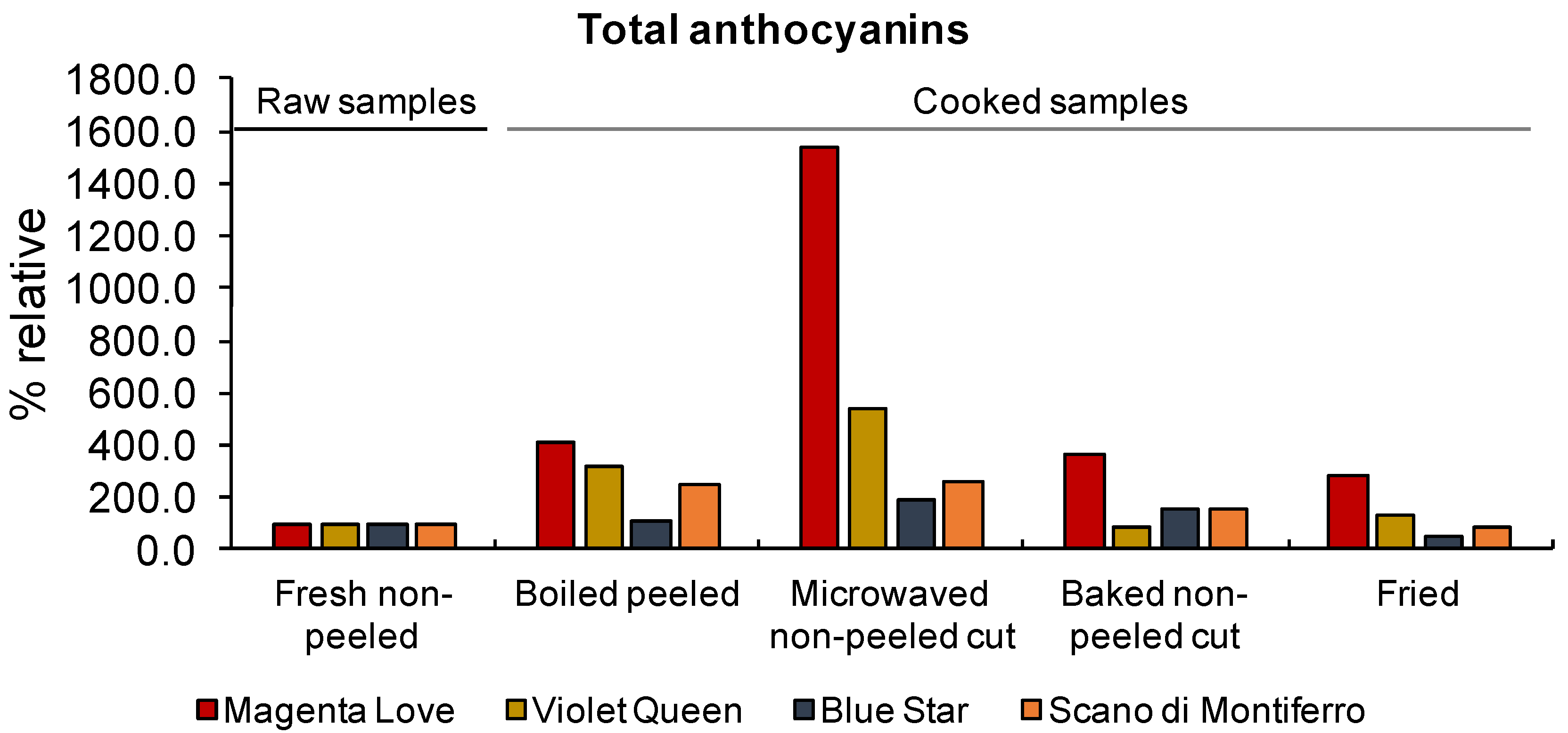
| Variety | Fresh Non-Peeled | Fresh Peeled | Boiled Peeled | Microwaved Non-Peeled Cut | Baked Non-Peeled Cut | Fried |
|---|---|---|---|---|---|---|
| Musica | 81.1 ± 4.3 (a) | 83.5 ± 12.2 (a) | 15.2 ± 1.9 (a) | 595.7 ± 16.4 (a) | 121.7 ± 18.9 (a) | 226.3 ± 21.2 (a) |
| Magenta Love | 3724.8 ± 152.5 (b) | 2615.6 ± 198.6 (b) | 8074.8 ± 221.5 (b) | 13,530.3 ± 315.6 (b) | 3668.8 ± 88.7 (b) | 2776.1 ± 51.8 (b) |
| Violet Queen | 1966.5 ± 74.1 (c) | 1382.7 ± 101.8 (c) | 4315.5 ± 84.6 (c) | 3748.7 ± 86.9 (c) | 2457.7 ± 95.6 (c) | 1002.3 ± 16.7 (c) |
| Blue star | 387.5 ± 33.6 (d) | 361.4 ± 24.1 (d) | 388.7 ± 21.3 (d) | 3115.7 ± 25.7 (d) | 1175.6 ± 102.3 (d) | 371.0 ± 3.9 (d) |
| Scano di Montiferro | 1685.6 ± 57.9 (c) | 1059.9 ± 31.2 (c) | 2119.8 ± 14.2 (e) | 5785.7 ± 66.4 (e) | 1594.3 ± 109.7 (e) | 865.0 ± 11.2 (c) |
| Variety | Raw Non-Peeled | Raw Peeled |
|---|---|---|
| Musica | nd | nd |
| Magenta Love | 471.2 ± 30.1 | 1054.1 ± 1.4 |
| Violet Queen | 1688.3 ± 70.2 | 1311.0 ± 115.7 |
| Blue Star | 131.3 ± 10.7 | 136.7 ± 15.9 |
| Scano di Montiferro | 984.5 ± 37.5 | 1213.0 ± 50.1 |
| Variety | Petunidin 3-O rutinoside * | Petunidin 3-O-caffeoyl-rutinoside-5-O-glucoside * | Petunidin 3-O-p-coumaroyl rutinoside 5-O-glucoside * | Petunidin 3-O-feruloyl rutinoside 5-O-glucoside * | Malvidin 3-O-p-coumaroyl-rutinoside-5-O-glucoside * | Malvidin 3-O-p-feruloyl-rutinoside-5-O-glucoside * | |
|---|---|---|---|---|---|---|---|
| Violet Queen | Fresh non-peeled | 19.4 ± 2.0 b | 27.4 ± 1.3 b | 360.9 ± 13.0 b | 17.0 ± 0.8 c | 98.7 ± 3.2 b | 12.8 ± 0.9 b |
| Fresh peeled | 17.6 ± 1.0 b | 60.5 ± 2.9 b | 912.5 ± 33.9 b | 31.7 ± 0.4 c | 177.0 ± 3.8 b | 22.3 ± 1.6 a | |
| Blue Star | Fresh non-peeled | <0.5 a | 11.3 ± 0.8 a | 24.8 ± 1.3 a | 11.4 ± 0.7 b | 15.5 ± 0.7 a | 11.5 ± 1.0 a |
| Fresh peeled | <0.5 a | 15.4 ± 0.8 a | 32.9 ± 1.8 a | 8.9 ± 0.3 b | 11.5 ± 0.9 a | 8.9 ± 0.6 b | |
| Scano di Montiferro | Fresh non-peeled | 31.3 ± 1.8 c | 16.2 ± 1.0 a | 13.3 ± 1.0 a | N.D. a | 350.1 ± 15.5 c | 29.5 ± 1.9 c |
| Fresh peeled | 36.7 ± 2.0 c | 21.9 ± 1.1 a | 42.3 ± 1.4 a | N.D. a | 443.1 ± 17.3 c | 34.8 ± 1.9 c |
| Variety | Petunidin 3-O-rutinoside-5-glucoside * | Pelargonidin 3-O-rutinoside * | Pelargonidin 3-O-p-caffeoyl- rutinoside-5-O-glucoside * | Pelargonidin 3-O-cis-p-coumaroyl-rutinoside-5-O-glucoside * | Pelargonidin 3-O-p-coumaroyl- rutinoside-5-O-glucoside * | Peonidin 3-O-p-coumaroyl- rutinoside-5-O-glucoside * | Pelargonidin 3-O-p-feruloyl- rutinoside-5-O-glucoside * | Peonidin 3-O-p-feruloyl- rutinoside-5-O-glucoside * | |
|---|---|---|---|---|---|---|---|---|---|
| Magenta Love | Fresh non-peeled | <0.5 mg/kg | <0.5 | 23.8 ± 1.1 | 26.7 ± 2.4 | 129.1 ± 9.0 | 30.0 ± 2.0 | 25.3 ± 1.0 | 20.6 ± 1.0 |
| Fresh peeled | 54.0 ± 4.1 | 19.4 ± 1.9 | 39.0 ± 2.5 | 82.3 ± 6.5 | 573.6 ± 21.8 | 90.2 ± 6.8 | 42.6 ± 2.9 | 16.2 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amelia, V.; Sarais, G.; Fais, G.; Dessì, D.; Giannini, V.; Garramone, R.; Carputo, D.; Melito, S. Biochemical Characterization and Effects of Cooking Methods on Main Phytochemicals of Red and Purple Potato Tubers, a Natural Functional Food. Foods 2022, 11, 384. https://doi.org/10.3390/foods11030384
D’Amelia V, Sarais G, Fais G, Dessì D, Giannini V, Garramone R, Carputo D, Melito S. Biochemical Characterization and Effects of Cooking Methods on Main Phytochemicals of Red and Purple Potato Tubers, a Natural Functional Food. Foods. 2022; 11(3):384. https://doi.org/10.3390/foods11030384
Chicago/Turabian StyleD’Amelia, Vincenzo, Giorgia Sarais, Giacomo Fais, Debora Dessì, Vittoria Giannini, Raffaele Garramone, Domenico Carputo, and Sara Melito. 2022. "Biochemical Characterization and Effects of Cooking Methods on Main Phytochemicals of Red and Purple Potato Tubers, a Natural Functional Food" Foods 11, no. 3: 384. https://doi.org/10.3390/foods11030384
APA StyleD’Amelia, V., Sarais, G., Fais, G., Dessì, D., Giannini, V., Garramone, R., Carputo, D., & Melito, S. (2022). Biochemical Characterization and Effects of Cooking Methods on Main Phytochemicals of Red and Purple Potato Tubers, a Natural Functional Food. Foods, 11(3), 384. https://doi.org/10.3390/foods11030384