
Analysis and Comparison of Nutrition Profiles of Canine Milk with Bovine and Caprine Milk




Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Collection of Milk Samples
2.3. Analysis of Basic Chemical Composition
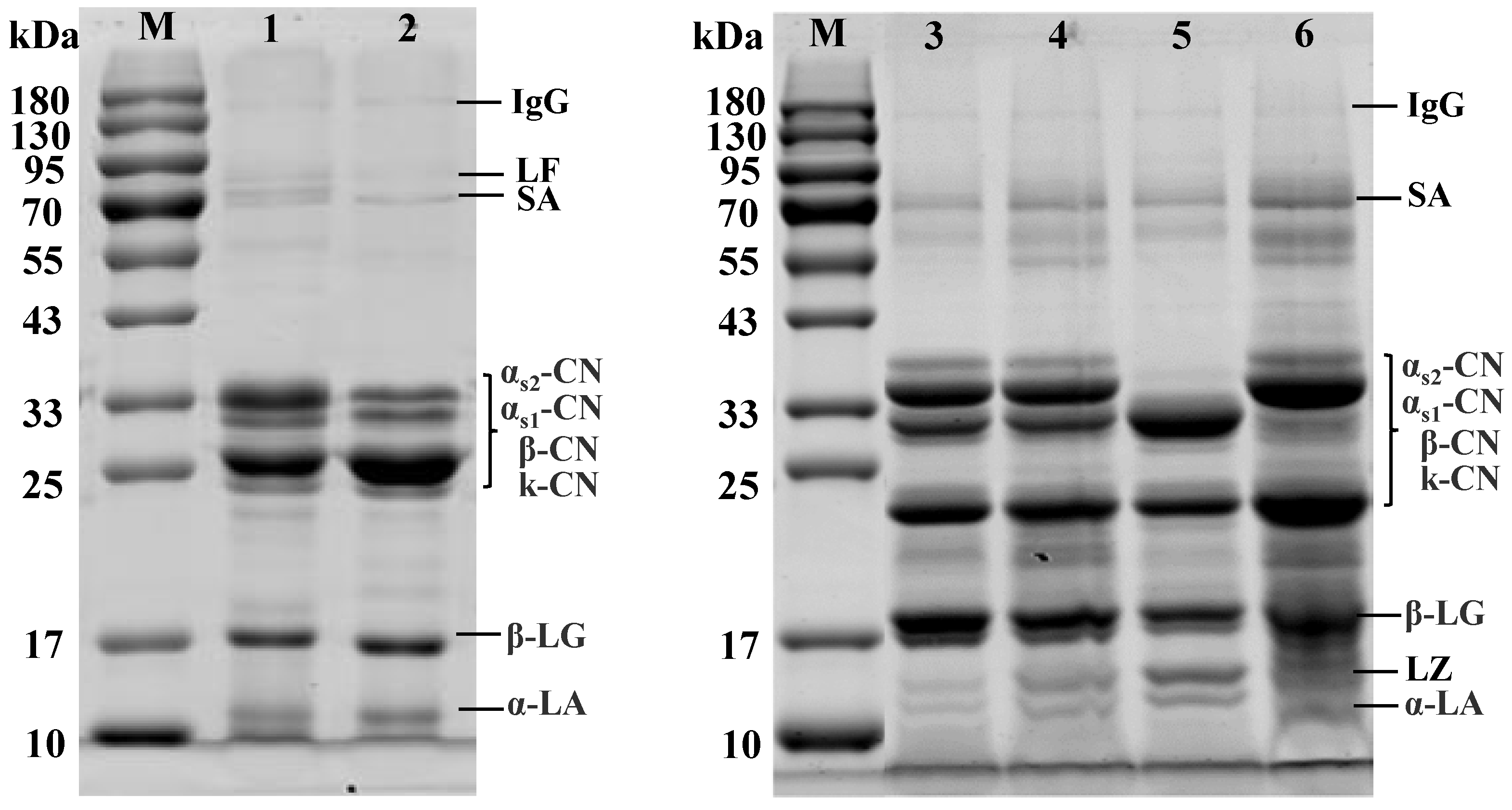
2.4. SDS-PAGE Analysis
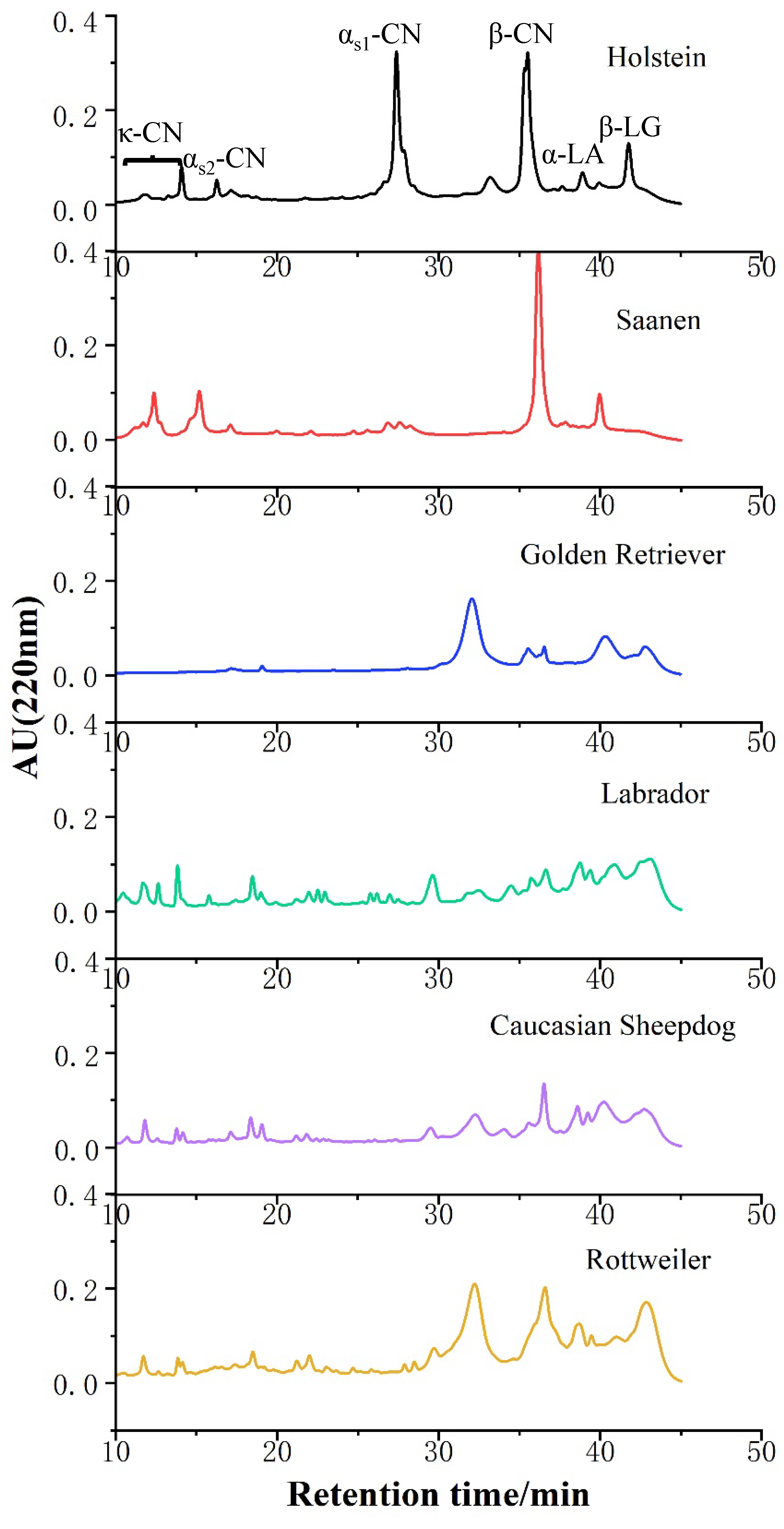
2.5. Analysis of Reverse Phase High Performance Liquid Chromatography (RP-HPLC)
2.6. Analysis of Fatty Acids (FAs)
2.7. Analysis of Minerals
2.8. Statistical Analysis
3. Results and Discussion
3.1. Analysis of the Chemical Composition of Bovine, Caprine, and Canine Milk
3.2. Protein Profiles of Bovine, Caprine, and Canine Milk by SDS-PAGE
3.3. Identification of Proteins in Canine, Bovine, and Caprine Milk by RP-HPLC
3.4. The Fatty Acid Profile Composition of Canine, Bovine, and Caprine Milk
3.5. Mineral Contents in Canine, Bovine, and Caprine Milk
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nakamura, T.; Urashima, T.; Mizukami, T.; Fukushima, M.; Arai, I.; Senshu, T.; Imazu, K.; Nakao, T.; Saito, T.; Ye, Z.; et al. Composition and oligosaccharides of a milk sample of the giant panda, Aduropoda melanoleuca. Comp. Biochem. Physiol. Part B 2003, 135, 439–448. [Google Scholar] [CrossRef]
- Tonini, B.; Fusi, E.; Rizzi, R.; Bontempo, V. Nucleotides in canine colostrum and milk at different stages of lactation. Arch. Anim. Nutr. 2010, 64, 337–341. [Google Scholar] [CrossRef]
- Field, M.C. The immunological components of human milk and their effect on immune development in infant. J. Nutr. 2005, 135, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Boza, J.; Sanz Sampelayo, M.R. Aspectos nutricionales de la leche de cabra. Ann. Acad. Cienc. Vet. Andal. Oriental. 1997, 10, 109–139. [Google Scholar]
- Corbee, R.J.; Tryfonidou, M.A.; Beckers, I.P.; Hazewinkel, H.A. Composition and use of puppy milk replacers in german shepherd puppies in the netherlands. J. Anim. Physiol. Anim. Nutr. 2012, 96, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Debraekeleer, J. Comparative analysis of milk replacers for puppies and kittens. J. Anim. Physiol. Anim. Nutr. 1998, 80, 185–193. [Google Scholar] [CrossRef]
- Macias Rostami, S.; Benet, T.; Spears, J.; Reynolds, A.; Satyaraj, E.; Sprenger, N.; Austin, S. Milk oligosaccharides over time of lactation from different dog breeds. PLoS ONE 2014, 9, e99824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mila, H.; Feugier, A.; Grellet, A.; Anne, J.; Gonnier, M.; Martin, M.; Rossig, L.; Chastant-Maillard, S. Immunoglobulin G concentration in canine colostrum: Evaluation and variability. J. Reprod. Immunol. 2015, 112, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Wrigglesworth, D.J.; Goonatilleke, E.; Haydock, R.; Hughes, K.R.; Lebrilla, C.B.; Swanson, K.S.; Jones, P.; Watson, P. High-throughput glycomic analyses reveal unique oligosaccharide profiles of canine and feline milk samples. PLoS ONE 2020, 15, e0243323. [Google Scholar] [CrossRef]
- Barlowska, J.; Szwajkowska, M.; Litwinczuk, Z.; Krol, J. Nutritional value and technological suitability of milk from various animal species used for dairy production. Compr. Rev. Food Sci. Food Saf. 2011, 10, 291–302. [Google Scholar] [CrossRef]
- Giffard, C.; Seino, M.; Markwell, P.; Bektash, R. Benefits of bovine colostrum on fecal quality in recently weaned puppies. J. Nutr. 2004, 134, 2126S–2127S. [Google Scholar] [CrossRef]
- Satyaraj, E.; Reynolds, A.; Pelker, R.; Labuda, J.; Zhang, P.; Sun, P. Supplementation of diets with bovine colostrum influences immune function in dogs. Br. J. Nutr. 2013, 110, 2216–2221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, E.J. Gastrointestinal aspects of food allergy: A review. J. Small Anim. Pract. 2008, 35, 145–152. [Google Scholar] [CrossRef]
- Zhang, H.; Yao, J.; Zhao, D.; Liu, H.; Li, J.; Guo, M. Changes in Chemical Composition of Alxa Bactrian Camel Milk During Lactation. J. Dairy Sci. 2005, 88, 3402–3410. [Google Scholar] [CrossRef]
- IDF 105E; Milk—Determination of Fat Content. IDF: Schaerbeek, Belgium, 2008.
- IDF 021B; Milk, Cream and Evaporated Milk-Determination of Total Solids Content (Reference Method). IDF: Schaerbeek, Belgium, 1987.
- IDF 027; Determination of the Ash Content of Processed Cheese Products. IDF: Schaerbeek, Belgium, 1964.
- Wang, H.; Sun, X.; Song, X.; Guo, M. Effects of kefir grains from different origins on proteolysis and volatile profile of goat milk kefir. Food Chem. 2021, 339, 128099. [Google Scholar] [CrossRef]
- Bonizzi, I.; Buffoni, J.N.; Feligini, M. Quantification of bovine casein fractions by direct chromatographic analysis of milk. Approaching the application to a real production context. J. Chromatogr. A 2009, 1216, 165–168. [Google Scholar] [CrossRef]
- Bobe, G.; Beitz, D.C.; Freeman, A.E.; Lindberg, G.L. Separation and Quantification of Bovine Milk Proteins by Reversed-Phase High-Performance Liquid Chromatography. J. Agric. Food Chem. 1998, 46, 458–463. [Google Scholar] [CrossRef]
- Bonfatti, V.; Grigoletto, L.; Cecchinato, A.; Gallo, L.; Carnier, P. Validation of a new reversed-phase high-performance liquid chromatography method for separation and quantification of bovine milk protein genetic variants. J. Chromatogr.A 2008, 1195, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, X.; Liu, L.; da Zhang, H.; Zhang, Y.; Hao Chang, Y.; Zhu, Q.P. Comparative lipidomics analysis of human, bovine and caprine milk by UHPLC-Q-TOF-MS. Food Chem. 2020, 310, 125865. [Google Scholar] [CrossRef]
- GB 5009.93; National Food Safety Standard. Determination of Multi-Element in Foods. GB: Beijing, China, 2003.
- Park, Y.W. Rheological characteristics of goat and sheep milk. Small Rumin. Res. 2007, 68, 73–87. [Google Scholar] [CrossRef]
- Heinze, C.; Freeman, L.; Martin, C.; Power, M.; Fascetti, A. Comparison of the nutrient composition of commercial dog milk replacers with that of dog milk. J. Am. Vet. Med. A 2014, 244, 1413–1422. [Google Scholar] [CrossRef] [Green Version]
- Oftedal, O. Lactation in the Dog milk composition and intake by Puppies. J. Nutr. 1984, 114, 803–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baines, F.M. Milk substitutes and the hand rearing of orphan puppies and kittens. J. Small Anim. Pract. 2010, 22, 555–578. [Google Scholar]
- Molinari, C.E.; Casadio, Y.S.; Hartmann, B.T.; Livk, A.; Bringans, S.; Arthur, P.G.; Hartmann, P.E. Proteome mapping of human skim milk proteins in term and preterm milk. J. Proteome Res. 2012, 11, 1696–1714. [Google Scholar] [CrossRef] [PubMed]
- Emmett, P.M.; Rogers, I.S. Properties of human milk and their relationship with maternal nutrition. Early Hum. Dev. 1997, 49, S7–S28. [Google Scholar] [CrossRef]
- Ha, M.; Bekhit, A.E.-D.; McConnell, M.; Mason, S.; Carne, A. Fractionation of whey proteins from red deer (Cervus elaphus) milk and comparison with whey proteins from cow, sheep and goat milks. Small Rumin. Res. 2014, 120, 125–134. [Google Scholar] [CrossRef]
- Halliday, J.A.; Bell, K.; Shaw, D.C. Feline and canine milk lysozymes. Comp. Biochen. Phys. B 1993, 106, 859–865. [Google Scholar] [CrossRef]
- Moatsou, G.; Moschopoulou, E.; Mollé, D.; Gagnaire, V.; Kandarakis, I.; Léonil, J. Comparative study of the protein fraction of goat milk from the Indigenous greek breed and from international breeds. Food Chem. 2008, 106, 509–520. [Google Scholar] [CrossRef]
- Landete-Castillejos, T.; Garcia, A.; Molina, P.; Vergara, H.; Garde, J.; Gallego, L. Milk production and composition in captive iberian red deer (Cervus elaphus hispanicus) effect of birth date. J. Anim. Sci. 2000, 78, 2771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenness, R. Comparative aspects of milk proteins. J. Dairy Res. 1979, 46, 197–210. [Google Scholar] [CrossRef]
- Park, Y. Hypo-allergenic and therapeutic significance of goat milk. Small Rumin. Res. 1994, 14, 151–159. [Google Scholar] [CrossRef]
- Lara-Villoslada, F.; Olivares, M.; Jimenez, J.; Boza, J.; Xaus, J. Goat milk is less immunogenic than cow milk in a murine model of atopy. J. Pediatr. Gastr. Nutr. 2004, 39, 354–360. [Google Scholar] [CrossRef] [Green Version]
- El-Zahar, K.; Sitohy, M.; Choiset, Y.; Métro, F.; Haertlé, T.; Chobert, J.-M. Peptic hydrolysis of ovine β-lactoglobulin and α-lactalbumin exceptional susceptibility of native ovine β-lactoglobulin to pepsinolysis. Int. Dairy J. 2005, 15, 17–27. [Google Scholar] [CrossRef]
- Yurchenko, S.; Sats, A.; Tatar, V.; Kaart, T.; Mootse, H.; Joudu, I. Fatty acid profile of milk from saanen and swedish landrace goats. Food Chem. 2018, 254, 326–332. [Google Scholar] [CrossRef]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [Green Version]
- Kumari, M.; Kozyrskyj, A.L. Gut microbial metabolism defines host metabolism: An emerging perspective in obesity and allergic inflammation. Obes. Rev. 2017, 18, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Zhao, G.; Xiang, J.; Zou, X.; Jin, Q.; Wang, X. Lipid composition and structural characteristics of bovine, caprine and human milk fat globules. Int. Dairy J. 2016, 56, 64–73. [Google Scholar] [CrossRef]
- Konuspayeva, G.; Lemarie, É.; Faye, B.; Loiseau, G.; Montet, D. Fatty acid and cholesterol composition of camel’s (camelus bactrianus, camelus dromedariusand hybrids) milk in kazakhstan. Dairy Sci. Technol. 2008, 88, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Koletzko, B.; Rodriguez-Palmero, M. Polyunsaturated fatty acids in human milk and their role in early infant development. J. Mammary Gland. Biol. Neoplasia 1999, 4, 269–284. [Google Scholar] [CrossRef]
- Bauer, J.; Heinemann, K.; Lees, G.; Waldron, M. Retinal functions of young dogs are improved and maternal plasma phospholipids are altered with diets containing long-chain n-3 polyunsaturated fatty acids during gestation, lactation, and after weaning. J. Nutr. 2006, 136, 1991S–1994S. [Google Scholar] [CrossRef]
- Zicker, S.; Jewell, D.; Yamka, R.; Milgram, N. Evaluation of cognitive learning, memory, psychomotor, immunologic, and retinal functions in healthy puppies fed foods fortified with docosahexaenoic acid–rich fish oil from 8 to 52 weeks of age. J. Am. Vet. Med. A 2012, 241, 583–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Pasha, T.; Farooq, U.; Ditta, Y.; Ilyas, M.; Ahmad, H. Mapping of calcium and phosphorus status of buffaloes in different cropping zone of punjab province. J Anim. Plant Sci. Pak. 2012, 22, 315–318. [Google Scholar]
- Haenlein, G.F.W. Past, present, and future perspectives of small ruminant dairy research. J. Dairy Sci. 2001, 84, 2097–2115. [Google Scholar] [CrossRef]
- Anderson, R.S.; Carlos, G.M.; Robinson, I.P.; Booles, D.; Burger, I.H.; Whyte, A.L. Zinc, copper, iron and calcium concentrations in bitch milk. J. Nutr. 1991, 121, S81–S82. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Nutrient | Holstein | Saanen | Golden Retriever | Labrador | Caucasian Sheepdog | Rottweiler |
|---|---|---|---|---|---|---|
| Moisture | 88.64 ± 0.11 a | 87.62 ± 0.1 a | 79.41 ± 0.16 b | 78.45 ± 0.65 b | 78.79 ± 0.08 b | 64.97 ± 0.47 c |
| Dry matter | 11.36 ± 0.11 a | 12.38 ± 0.1 a | 20.59 ± 0.16 b | 22.23 ± 1.73 c | 21.21 ± 0.08 bc | 35.03 ± 0.47 d |
| Crude protein | 3.12 ± 0.14 a | 3.09 ± 0.05 a | 7.57 ± 0.5 b | 8.34 ± 0.06 c | 6.62 ± 0.06 d | 17.34 ± 0.28 e |
| Fat | 3.71 ± 0.24 a | 4.45 ± 0.31 b | 8.92 ± 0.17 c | 9.94 ± 0.52 d | 9.56 ± 0.28 d | 14.31 ± 0.29 e |
| Ash | 0.82 ± 0.01 a | 0.88 ± 0.02 a | 1.23 ± 0.04 c | 1.19 ± 0.08 bc | 1.11 ± 0.04 b | 1.81 ± 0.05 d |
| Lactose | 3.71 ± 0.25 a | 3.95 ± 0.38 a | 2.86 ± 0.64 ab | 2.76 ± 1.53 ab | 3.92 ± 0.33 a | 1.56 ± 0.33 b |
| Holstein | Saanen | Golden Retriever | Labrador | Caucasian Sheepdog | Rottweiler | |
|---|---|---|---|---|---|---|
| αs1-CN | 9.45 ± 0.37 b | 3.16 ± 0.44 a | 9.67 ± 0.60 b | 4.27 ± 0.24 a | 7.76 ± 1.04 b | 30.98 ± 3.95 c |
| αs2-CN | 1.86 ± 0.04 a | 7.26 ± 0.28 b | ND | 4.83 ± 0.78 c | 4.30 ± 0.43 c | 6.73 ± 0.63 b |
| β-CN | 6.59 ± 0.31 a | 9.19 ± 0.66 b | 2.27 ± 0.83 c | 5.19 ± 0.46 d | 6.50 ± 0.49 a | 17.26 ± 0.17 e |
| κ-CN | 3.20 ± 0.02 a | 5.74 ± 0.16 b | ND | ND | ND | ND |
| β-LG | 3.35 ± 0.82 a | 2.79 ± 0.76 a | 7.84 ± 1.25 b | 12.56 ± 3.11 d | 11.5 ± 0.73 cd | 8.69 ± 0.88 bc |
| α-LA | 0.78 ± 0.18 a | 0.67 ± 0.07 a | 0.92 ± 0.10 a | 4.08 ± 0.18 b | 3.59 ± 0.35 c | 4.99 ± 0.49 d |
| Holstein | Saanen | Golden Retriever | Labrador | Caucasian Sheepdog | Rottweiler | |
|---|---|---|---|---|---|---|
| C4:0 | 0.48 ± 0.11 a | 0.64 ± 0.19 a | 0.25 ± 0.08 b | 0.24 ± 0.04 b | 0.26 ± 0.11 b | 0.15 ± 0.06 b |
| C6:0 | 0.94 ± 0.18 ab | 2.06 ± 0.31 c | 0.76 ± 0.04 ab | 0.82 ± 0.1 ab | 0.62 ± 0.06 a | 1.01 ± 0.25 b |
| C10:0 | 0.78 ± 0.04 b | 2.43 ± 0.48 a | 0.30 ± 0.1 c | 0.17 ± 0.03 c | 0.43 ± 0.03 bc | 0.19 ± 0.03 c |
| C12:0 | 2.29 ± 0.08 a | 2.94 ± 0.21 b | 0.31 ± 0.02 d | 0.09 ± 0.01 e | 0.69 ± 0.03 c | 0.10 ± 0.02 e |
| C14:0 | 13.36 ± 0.6 a | 13.93 ± 0.54 a | 3.25 ± 0.08 c | 3.23 ± 0.7 c | 5.28 ± 0.14 b | 1.63 ± 0.06 d |
| C14:1 | 0.68 ± 0.04 a | 0.28 ± 0.05 b | 0.08 ± 0.01 c | 0.12 ± 0.06 c | 0.16 ± 0.01 c | ND |
| C16:0 | 38.50 ± 0.87 a | 36.71 ± 0.93 ab | 33.96 ± 0.7 ab | 32.39 ± 7.88 ab | 32.68 ± 5.58 ab | 30.03 ± 5.03 b |
| C16:1 | ND | ND | 2.28 ± 0.17 a | 4.11 ± 0.16 b | 1.77 ± 0.08 c | 3.36 ± 0.02 d |
| C17:0 | 1.47 ± 0.01 a | 1.01 ± 0.04 b | 0.41 ± 0.07 c | 0.40 ± 0.04 c | 0.42 ± 0.02 c | 0.43 ± 0.03 c |
| C17:1 | ND | ND | ND | 0.31 ± 0.02 a | 0.01 ± 0.01 b | 0.23 ± 0.02 a |
| C18:0 | 11.76 ± 0.07 bc | 12.15 ± 0.76 c | 12.03 ± 0.83 c | 12.15 ± 1.88 c | 9.50 ± 1.41 a | 9.86 ± 0.76 ab |
| C18:1n9t | 5.30 ± 0.57 a | 0.46 ± 0.11 b | 12.81 ± 1.58 d | 11.12 ± 1.09 cd | 9.27 ± 1.50 c | 11.59 ± 2.17 cd |
| C18:1n9c | 20.24 ± 0.18 ab | 23.03 ± 0.59 a | 14.52 ± 2.54 c | 16.88 ± 4.82 bc | 17.15 ± 0.51 bc | 17.69 ± 3.35 bc |
| C18:2n6t | ND | ND | 0.21 ± 0.13 a | 0.43 ± 0.23 a | 0.17 ± 0.05 a | 0.33 ± 0.03 a |
| C18:2n6c | 1.32 ± 0.06 a | 1.08 ± 0.25 a | 11.79 ± 2.03 b | 11.24 ± 4.56 b | 13.58 ± 1.19 b | 15.41 ± 2.79 b |
| C20:0 | 0.39 ± 0.09 b | 0.73 ± 0.11 c | 0.19 ± 0.01 a | 0.21 ± 0.10 ab | 1.09 ± 0.16 d | 0.20 ± 0.05 a |
| C18:3n6 | 0.11 ± 0.02 a | 0.16 ± 0.02 a | 0.26 ± 0.11 a | 0.32 ± 0.01 a | 1.48 ± 0.47 b | 0.32 ± 0.05 a |
| C20:1 | 0.17 ± 0.02 a | 0.11 ± 0.02 a | 0.24 ± 0.06 a | 0.59 ± 0.11 b | 0.50 ± 0.08 b | 0.56 ± 0.08 b |
| C18:3n3 | 0.30 ± 0.07 cd | 0.60 ± 0.02 e | 0.19 ± 0.01 bc | 0.09 ± 0.06 a | 0.08 ± 0.02 ab | 0.40 ± 0.03 d |
| C21:0 | ND | 0.20 ± 0.11 a | 1.38 ± 0.84 b | 0.66 ± 0.03 ab | 0.76 ± 0.05 ab | 0.91 ± 0.15 ab |
| C20:2 | 0.74 ± 0.06 a | 0.96 ± 0.35 ab | 2.58 ± 0.44 d | 1.09 ± 0.17 abc | 1.33 ± 0.24 bc | 1.47 ± 0.09 c |
| C22:0 | 0.28 ± 0.01 bc | 0.26 ± 0.03 b | 0.36 ± 0.05 c | 0.52 ± 0.07 d | 0.08 ± 0.03 a | 0.16 ± 0.02 a |
| C20:3 | ND | ND | 0.46 ± 0.06 a | 0.53 ± 0.24 a | 0.66 ± 0.04 ab | 0.83 ± 0.11 b |
| C22:1 | 0.20 ± 0.01 a | 0.36 ± 0.08 ab | 0.56 ± 0.05 b | 1.17 ± 0.25 c | 1.34 ± 0.01 c | 0.51 ± 0.12 b |
| C24:1n9 | 0.07 ± 0.01 a | ND | 0.02 ± 0.01 b | ND | 0.01 ± 0.00 c | 0.08 ± 0.05 a |
| C20:4 | 0.24 ± 0.17 a | ND | 0.82 ± 0.28 b | 1.77 ± 0.42 c | 1.29 ± 0.14 bc | 1.43 ± 0.23 c |
| C20:5 | 0.1 ± 0.01 a | 0.13 ± 0.01 a | 0.82 ± 0.13 b | 0.99 ± 0.05 c | 0.08 ± 0.02 a | 0.15 ± 0.02 a |
| C24:0 | 0.23 ± 0.08 a | ND | ND | ND | 0.07 ± 0.01 b | ND |
| C22:6 | ND | ND | ND | 0.43 ± 0.07 a | 0.12 ± 0.01 b | 0.35 ± 0.02 a |
| Sums of fatty acids | ||||||
| SFA | 70.30 ± 0.56 a | 72.99 ± 2.3 a | 53.18 ± 2.4 b | 50.32 ± 10.20 b | 51.49 ± 4.8 b | 47.57 ± 2.6 b |
| MUFA | 26.61 ± 0.85 ab | 24.10 ± 0.27 a | 29.71 ± 2.61 bc | 32.95 ± 3.94 c | 30.2 ± 1.10 bc | 32.63 ± 2.44 c |
| PUFA | 2.91 ± 0.17 a | 2.90 ± 0.38 a | 17.07 ± 2.52 b | 16.83 ± 4.43 b | 18.19 ± 0.45 b | 20.56 ± 3.30 b |
| n-6 | 1.69 ± 0.13 a | 1.25 ± 0.27 a | 13.07 ± 2.19 b | 13.59 ± 4.37 b | 15.96 ± 0.27 b | 17.49 ± 3.11 b |
| n-3 | 0.40 ± 0.05 a | 0.72 ± 0.01 ab | 1.40 ± 0.07 bc | 1.70 ± 0.71 c | 0.90 ± 0.03 ab | 1.71 ± 0.78 c |
| Holstein | Saanen | Golden Retriever | Labrador | Caucasian Sheepdog | Rottweiler | |
|---|---|---|---|---|---|---|
| Mg | 9.63 | 14.3 | 12.8 | 13 | 9.85 | 32.9 |
| K | 126.2 | 152.1 | 76.9 | 104.8 | 113.3 | 189.4 |
| Na | 49.9 | 60.6 | 104.5 | 110.2 | 80.7 | 92.1 |
| Fe | ND | ND | ND | 0.405 | 0.58 | 0.749 |
| Zn | 0.31 | 0.394 | 1.34 | 1.13 | 0.85 | 0.854 |
| Cu | ND | ND | 0.047 | 0.119 | 0.046 | 0.398 |
| Mn (μg/100 mL) | ND | ND | 10.6 | 11.7 | 16.6 | 10.6 |
| Se (μg/100 mL) | 2.36 | 1.06 | 10.6 | 11 | 10 | 39.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Sun, X.; Cheng, J.; Guo, M. Analysis and Comparison of Nutrition Profiles of Canine Milk with Bovine and Caprine Milk. Foods 2022, 11, 472. https://doi.org/10.3390/foods11030472
Zhang M, Sun X, Cheng J, Guo M. Analysis and Comparison of Nutrition Profiles of Canine Milk with Bovine and Caprine Milk. Foods. 2022; 11(3):472. https://doi.org/10.3390/foods11030472
Chicago/Turabian StyleZhang, Mengjie, Xiaomeng Sun, Jianjun Cheng, and Mingruo Guo. 2022. "Analysis and Comparison of Nutrition Profiles of Canine Milk with Bovine and Caprine Milk" Foods 11, no. 3: 472. https://doi.org/10.3390/foods11030472
APA StyleZhang, M., Sun, X., Cheng, J., & Guo, M. (2022). Analysis and Comparison of Nutrition Profiles of Canine Milk with Bovine and Caprine Milk. Foods, 11(3), 472. https://doi.org/10.3390/foods11030472