
Structural Characteristics of Insoluble Dietary Fiber from Okara with Different Particle Sizes and Their Prebiotic Effects in Rats Fed High-Fat Diet

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of OIDFs with Three Different Particle Sizes
2.2. Determination of the Basic Components of the Three OIDFs
2.3. Structure of the Three Different Granularity of OIDFs
2.3.1. Particle Size Determination
2.3.2. Scanning Electron Microscopy (SEM)
2.3.3. Fourier Infrared Spectrum (FT-IR)
2.3.4. Determination of Thermogravimetry (TG)
2.3.5. Determination of X-ray Diffraction (XRD)
2.4. Physicochemical Properties of the Three OIDFs
2.4.1. Water-Holding Capacity (WHC), Oil-Holding Capacity (OHC) and Water-Swelling Capacity (WSC)
2.4.2. Adsorption Capacity of the Three Particle Sizes of OIDF
Cholesterol-Adsorption Capacity (CAC)
Sodium Cholate-Adsorption Capacity (SCAC)
2.5. Animals and Experimental Diets
2.6. Lipid Analysis
2.7. Histological Analysis
2.8. RNA Extraction and Quantitative Real-Time PCR Analysis
2.9. Western Blot Analyses
2.10. Gut Microbiota Sequencing Analysis
2.11. Statistical Analysis
3. Results
3.1. Determination of the Basic Components of the Three OIDFs
3.2. Structure of the Three Different Granularity of OIDFs
3.3. Physicochemical Properties of the Three OIDFs
3.4. Effects of Different Particle Sizes of OIDF on Blood Lipid Levels in High-Fat Diet Rats
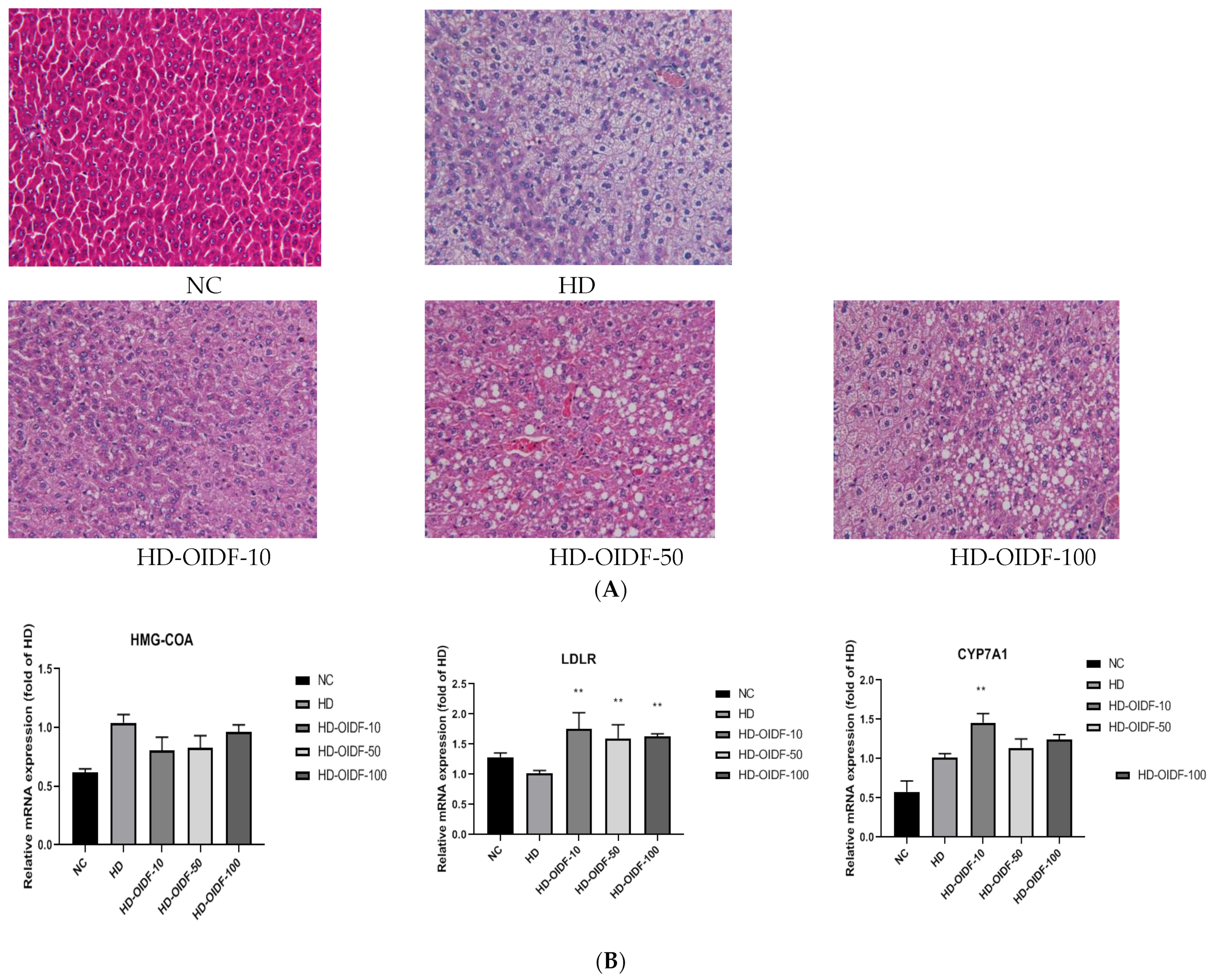
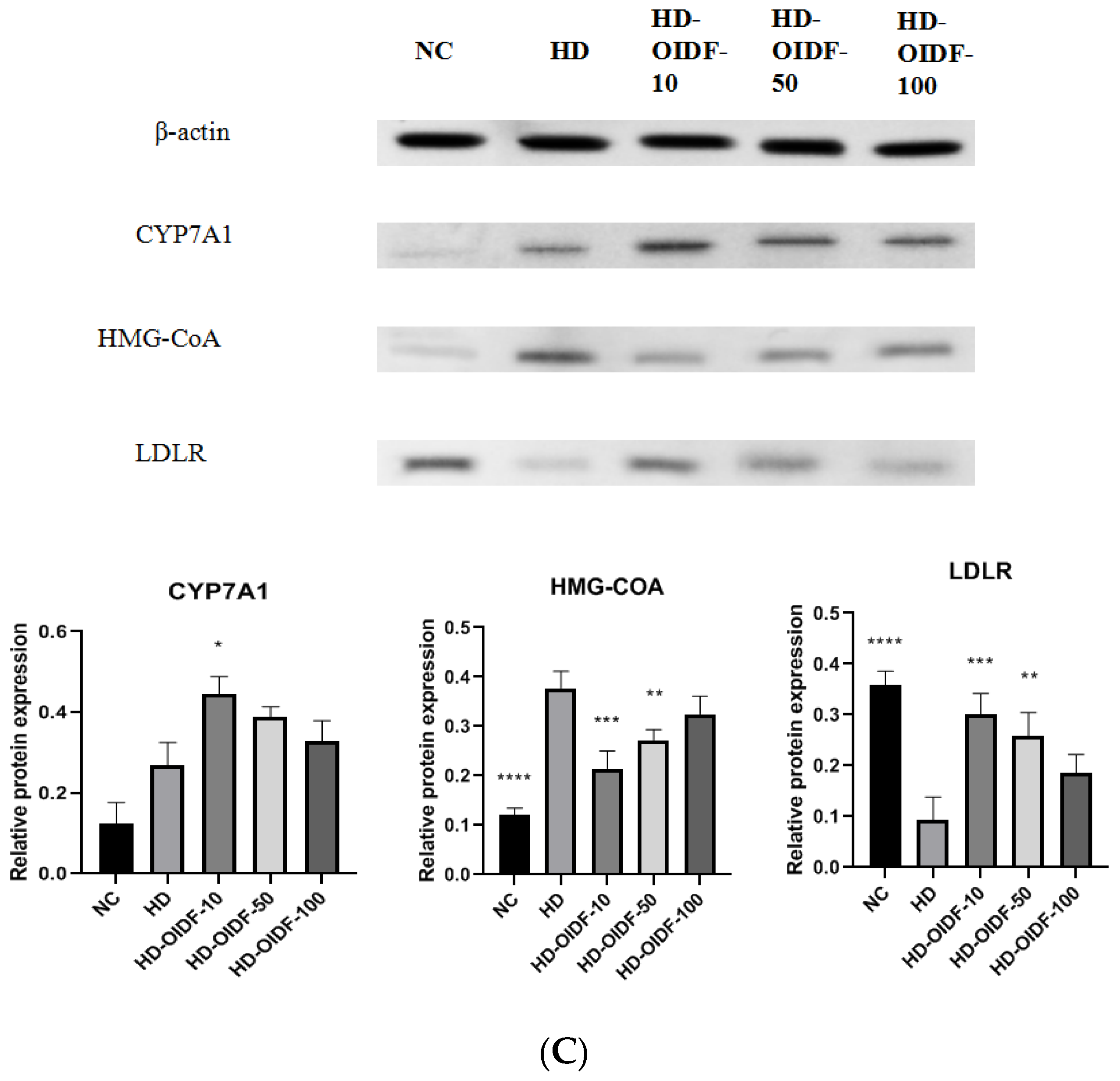
3.5. Effects of Different Particle Sizes of OIDF on Hepatic Steatosis in High-Fat Diet Rats
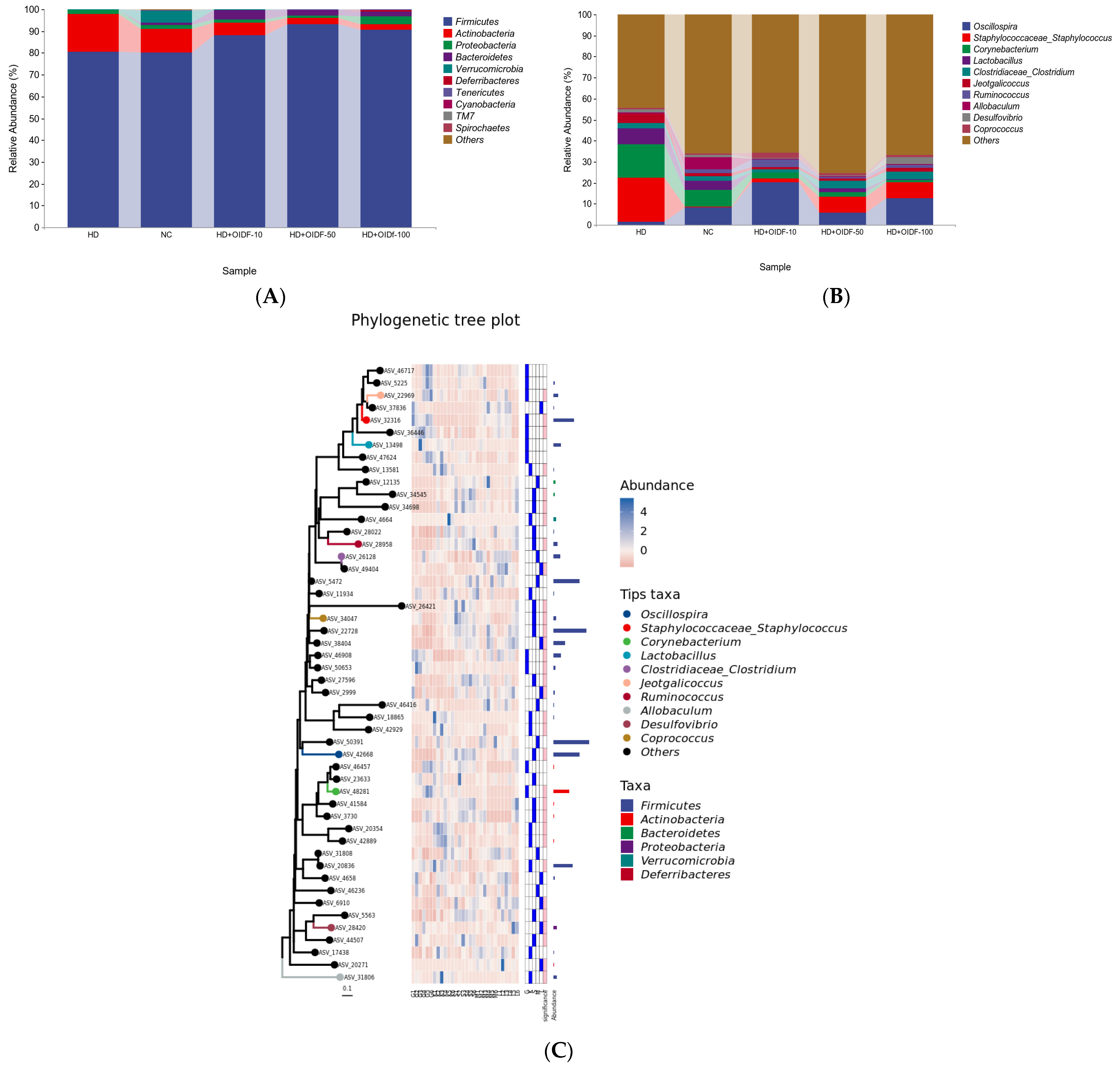
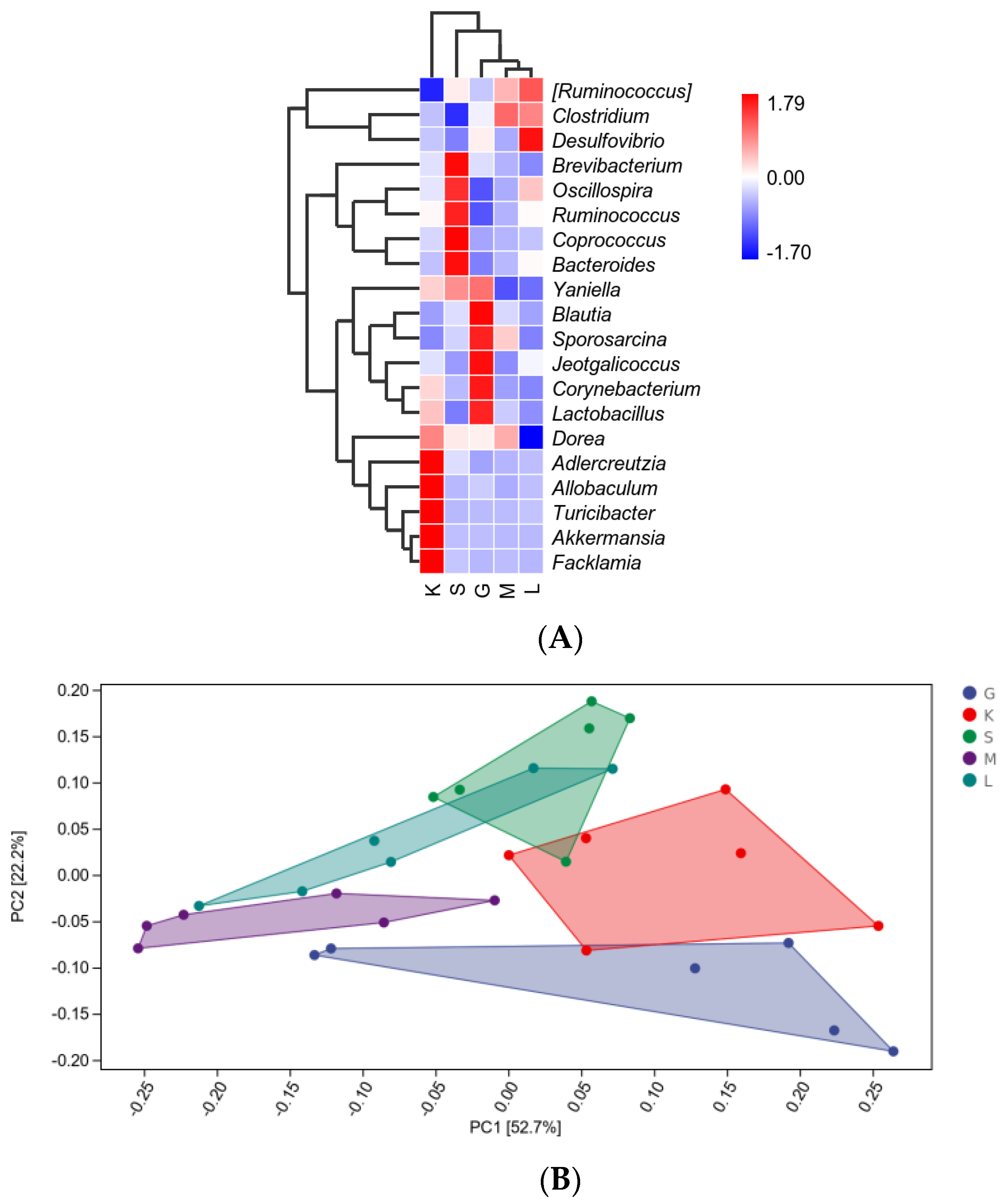
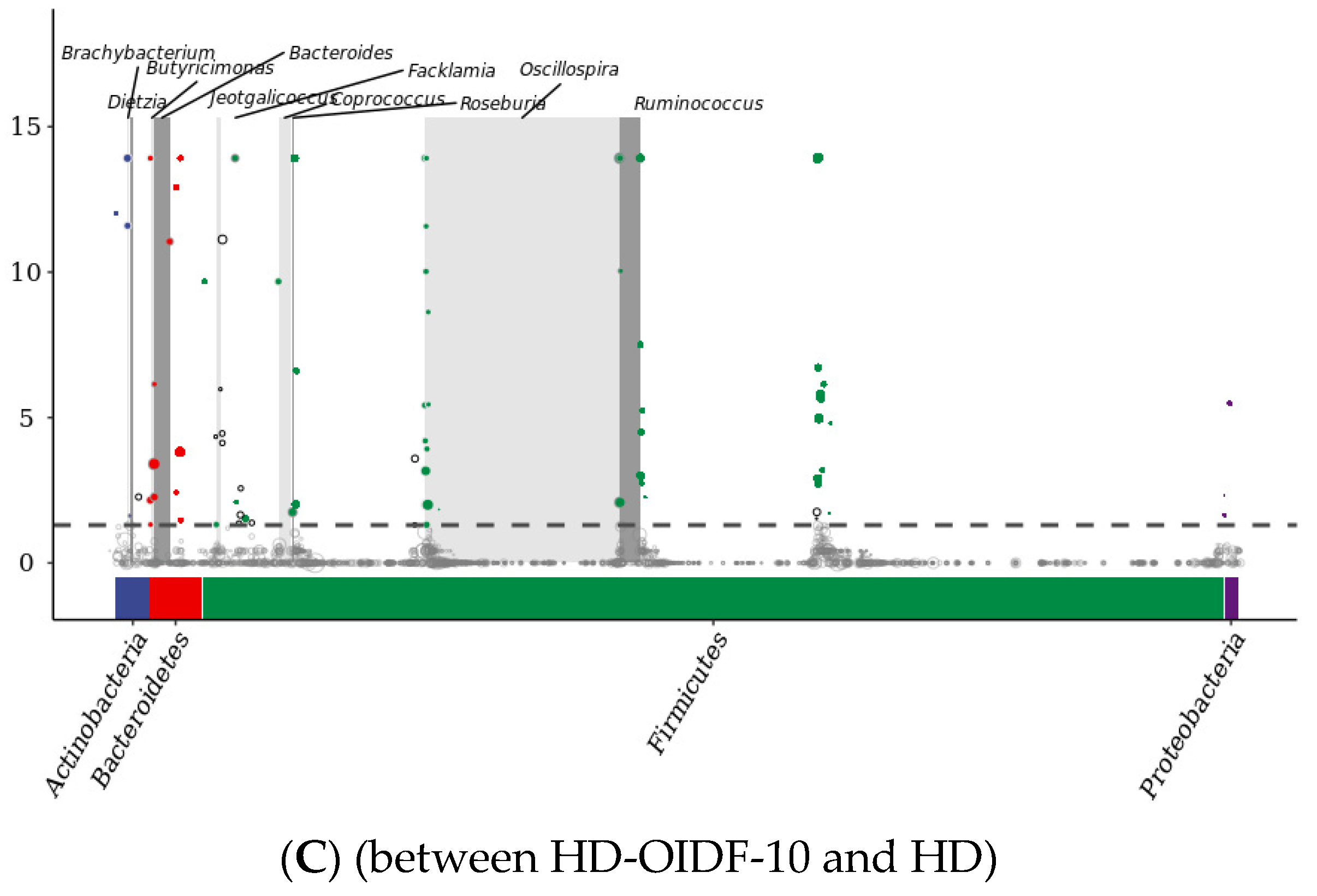
3.6. Bacterial Composition Analysis between Different Populations
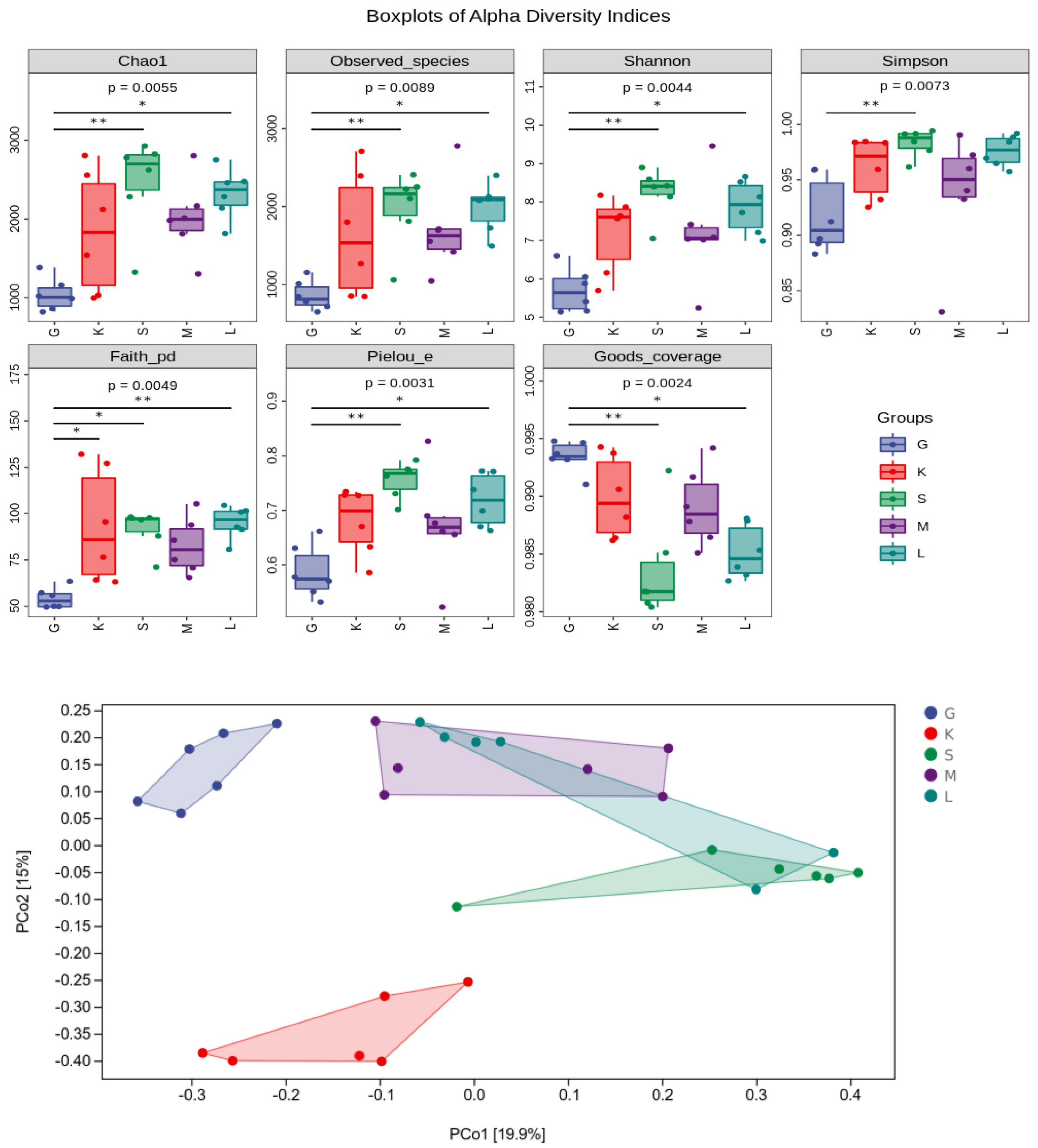
3.7. Alpha Diversity Index and Beta diversity PCoA Analysis
3.8. Alpha Diversity Index and Beta Diversity PCoA Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, S.N.; Yu, H.S.; Gu, C.; He, Y.; Zhang, T.; Xu, X. Preventive effect of soybean insoluble dietary fiber on high fat diet induced obesity in mice. Sci. Technol. Food Ind. 2020, 41, 295–301, 314. [Google Scholar]
- Swallah, M.S.; Fan, H.; Wang, S.; Yu, H.; Piao, C. Prebiotic Impacts of Soybean Residue (Okara) on Eubiosis/Dysbiosis Condition of the Gut and the Possible Effects on Liver and Kidney Functions. Molecules 2021, 26, 326. [Google Scholar] [CrossRef] [PubMed]
- Lyu, B.; Wang, Y.; Fu, H.; Li, J.; Yang, X.; Shen, Y.; Swallah, M.S.; Yu, Z.; Li, Y.; Wang, H.; et al. Intake of high-purity insoluble dietary fiber from Okara for the amelioration of colonic environment disturbance caused by acute ulcerative colitis. Food Funct. 2022, 13, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Ussar, S.; Griffin, N.W.; Bezy, O.; Fujisaka, S.; Vienberg, S.; Softic, S.; Deng, L.; Bry, L.; Gordon, J.I.; Kahn, C.R. Interactions between gut microbiota, host genetics and diet modulate the predisposition to obesity and metabolic syndrome. Cell Metab. 2015, 22, 516–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantu-Jungles, T.M.; Hamaker, B.R. New view on dietary fiber selection for predictable shifts in gut microbiota. MBio 2020, 11, e02179-19. [Google Scholar] [CrossRef] [Green Version]
- Adams, S.; Che, D.; Hailong, J.; Han, R.; Qin, G.; Danquah, K. Dietary supplementation of pulverised Astragalus membranaceus improved performance, immunity and diarrhoea incidence in weaned piglets. Indian J. Anim. Res. 2018, B-936. [Google Scholar] [CrossRef]
- Adams, S.; Sello, C.T.; Qin, G.X.; Che, D.; Han, R. Does dietary fiber affect the levels of nutritional components after feed formulation? Fibers 2018, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Adams, S.; Che, D.; Qin, G.; Rui, H.; Sello, C.T.; Hailong, J. Interactions of Dietary Fibre with Nutritional Components on Gut Microbial Composition, Function and Health in Monogastrics. Curr. Protein Pept. Sci. 2018, 19, 1011–1023. [Google Scholar] [CrossRef]
- Abedin, A.; Evera, E.; Qingzhen, Z.; Zewei, S. Gastrointestinal Interaction between Dietary Amino Acids and Gut Microbiota: With Special Emphasis on Host Nutrition. Curr. Protein Pept. Sci. 2020, 21, 785–798. [Google Scholar]
- Adams, S.; Xiangjie, K.; Hailong, J.; Guixin, Q.; Sossah, F.L.; Dongsheng, C. Prebiotic effects of alfalfa (Medicago sativa) fiber on cecal bacterial composition, short-chain fatty acids, and diarrhea incidence in weaning piglets. RSC Adv. 2019, 9, 13586–13599. [Google Scholar] [CrossRef] [Green Version]
- Piel, C.; Montagne, L.; Sève, B.; Lalleès, J.-P. Increasing digesta viscosity using carboxymethylcellulose in weaned piglets stimulates ileal goblet cell numbers and maturation. J. Nutr. 2005, 135, 86–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamont, J.T. Mucus: The front line of intestinal mucosal defense. Ann. N. Y. Acad. Sci. 1992, 664, 190–201. [Google Scholar] [CrossRef]
- Li, L.; Ma, L.; Fu, P. Gut microbiota–derived short-chain fatty acids and kidney diseases. Drug Des. Dev. Ther. 2017, 11, 3531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, F.Z.; Nelson, E.; Chu, P.Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-fiber diet and acetate supplementation change the gut microbiota and prevent the development of hypertension and heart failure in hypertensive mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Lee DP, S.; Gan, A.X.; Kim, J.E. Incorporation of biovalorised okara in biscuits: Improvements of nutritional, antioxidant, physical, and sensory properties. LWT 2020, 134, 109902. [Google Scholar]
- Chan, L.Y.; Takahashi, M.; Lim, P.J.; Aoyama, S.; Makino, S.; Ferdinandus, F.; Ng, S.Y.; Arai, S.; Fujita, H.; Tan, H.C.; et al. Eurotium Cristatum Fermented Okara as a Potential Food Ingredient to Combat Diabetes. Sci. Rep. 2019, 9, 17536. [Google Scholar] [CrossRef]
- Swallah, M.S.; Yu, H.; Piao, C.; Fu, H.; Yakubu, Z.; Sossah, F.L. Synergistic two-way interactions of dietary polyphenols and dietary components on the gut microbial composition: Is there a positive, negative, or neutralizing effect in the prevention and management of metabolic diseases? Curr. Protein Pept. Sci. 2021, 22, 313–327. [Google Scholar] [CrossRef]
- Xu, T.; Wu, X.; Liu, J.; Sun, J.; Wang, X.; Fan, G.; Meng, X.; Zhang, J.; Zhang, Y. The regulatory roles of dietary fibers on host health via gut microbiota-derived short chain fatty acids. Curr. Opin. Pharmacol. 2022, 62, 36–42. [Google Scholar] [CrossRef]
- Rodríguez, R.; Jimenez, A.; Fernández-Bolanos, J.; Guillen, R.; Heredia, A. Dietary fibre from vegetable products as source of functional ingredients. Trends Food Sci. Technol. 2006, 17, 3–15. [Google Scholar] [CrossRef]
- Zhang, H.; Troise, A.D.; Qi, Y.; Wu, G.; Zhang, H.; Fogliano, V. Insoluble dietary fibre scavenges reactive carbonyl species under simulated physiological conditions: The key role of fibre-bound polyphenols. Food Chem. 2021, 349, 129018. [Google Scholar] [CrossRef]
- Lyu, B.; Wang, H.; Swallah, M.S.; Fu, H.; Shen, Y.; Guo, Z.; Tong, X.; Li, Y.; Yu, H.; Jiang, L. Structure, Properties and Potential Bioactivities of High-purity Insoluble Fibre from Soybean Dregs (Okara). Food Chem. 2021, 364, 130402. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Wang, Y.; Wu, J.; Wang, Z. Functional Properties and Morphological Characters of Soluble Dietary Fibers in Different Edible Parts of Angelica Keiskei. J. Food Sci. 2016, 81, C2189. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Teng, F.; McClements, D.J.; Zhang, S.; Li, Y.; Wang, Z. Effect of cavitation jet processing on the physicochemical properties and structural characteristics of okara dietary fiber. Food Res. Int. 2020, 134, 109251. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Fan, L.; Yang, Y.; Jiang, Q.; Xu, Y.; Xia, W. Characterization of surimi particles stabilized novel Pickering emulsions: Effect of particles concentration, pH and NaCl levels. Food Hydrocoll. 2021, 117, 106731. [Google Scholar] [CrossRef]
- Ma, M.; Mu, T. Modification of deoiled cumin dietary fiber with laccase and cellulase under high hydrostatic pressure. Carbohydr. Polym. 2016, 136, 87–94. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Q.; Sun, J.; Jiang, B.; Yan, J. Extraction of water-soluble polysaccharide and the antioxidant activity from Semen cassiae. J. Food Drug Anal. 2014, 22, 492–499. [Google Scholar] [CrossRef] [Green Version]
- Alba, K.; Macnaughtan, W.; Laws, A.P.; Foster, T.J.; Campbell, G.M.; Kontogiorgos, V. Fractionation and characterisation of dietary fibre from blackcurrant pomace. Food Hydrocoll. 2018, 81, 398–408. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Niu, M.; Zhang, B.; Zhao, S.; Xiong, S. Structural characteristics and functional properties of rice bran dietary fiber modified by enzymatic and enzyme-micronization treatments. LWT-Food Sci. Technol. 2017, 75, 344–351. [Google Scholar] [CrossRef]
- Wang, Q.H.; Shu, Z.P.; Xu, B.Q.; Xing, N.; Jiao, W.J.; Yang, B.Y.; Kuang, H.X. Structural characterization and antioxidant activities of polysaccharides from Citrus aurantium L. Int. J. Biol. Macromol. 2014, 67, 112–123. [Google Scholar] [CrossRef]
- Tanpichai, S.; Biswas, S.K.; Witayakran, S.; Yano, H. Water Hyacinth: A Sustainable Lignin-Poor Cellulose Source for the Production of Cellulose Nanofibers. ACS Sustain. Chem. Eng. 2019, 7, 18884–18893. [Google Scholar] [CrossRef]
- Zhang, Z.; Song, H.; Peng, Z.; Luo, Q.; Ming, J.; Zhao, G. Characterization of stipe and cap powders of mushroom (Lentinus edodes) prepared by different grinding methods. J. Food Eng. 2012, 109, 406–413. [Google Scholar] [CrossRef]
- Jiang, L.; Xu, Q.X.; Qiao, M.; Ma, F.F.; Thakur, K.; Wei, Z.J. Effect of superfine grinding on properties of Vaccinium bracteatum Thunb leaves powder. Food Sci. Biotechnol. 2017, 26, 1571–1578. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Kuga, S.; Wang, C.; Zhao, Y.; Wu, M.; Huang, Y. Green preparation of cellulose nanocrystal and its application. ACS Sustain. Chem. Eng. 2018, 6, 2954–2960. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, H.; Li, Y.; Huang, Q. Fabrication of milled cellulose particles-stabilized Pickering emulsions. Food Hydrocoll. 2018, 77, 427–435. [Google Scholar] [CrossRef]
- Park, J.; Choi, I.; Kim, Y. Cookies formulated from fresh okara using starch, soy flour and hydroxypropyl methylcellulose have high quality and nutritional value. LWT-Food Sci. Technol. 2015, 63, 660–666. [Google Scholar] [CrossRef]
- Ahmed, J.; Al-Attar, H.; Arfat, Y.A. Effect of particle size on compositional, functional, pasting and rheological properties of commercial water chestnut flour. Food Hydrocoll. 2016, 52, 888–895. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, Y. Physicochemical and functional properties of coconut (Cocos nucifera L.) cake dietary fibres: Effects of cellulase hydrolysis, acid treatment and particle size distribution—ScienceDirect. Food Chem. 2018, 257, 135–142. [Google Scholar] [CrossRef]
- Zhou, A.L.; Hergert, N.; Rompato, G.; Lefevre, M. Whole Grain Oats Improve Insulin Sensitivity and Plasma Cholesterol Profile and Modify Gut Microbiota Composition in C57BL/6J Mice. J. Nutr. 2015, 145, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Hoang, M.H.; Houng, S.J.; Jun, H.J.; Lee, J.H.; Choi, J.W.; Kim, S.H.; Kim, Y.R.; Lee, S.J. Barley Intake Induces Bile Acid Excretion by Reduced Expression of Intestinal ASBT and NPC1L1 in C57BL/6J Mice. J. Agric. Food Chem. 2011, 59, 6798–6805. [Google Scholar] [CrossRef]
- Kemper, M.F.; Srivastava, S.; Todd King, M.; Clarke, K.; Veech, R.L.; Pawlosky, R.J. An Ester of β-Hydroxybutyrate Regulates Cholesterol Biosynthesis in Rats and a Cholesterol Biomarker in Humans. Lipids 2015, 50, 1185–1193. [Google Scholar] [CrossRef]
- Levinson, S.S.; Wagner, S.G. Implications of reverse cholesterol transport: Recent studies. Clin. Chim. Acta 2015, 439, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Musso, G.; Gambino, R.; Cassader, M. Cholesterol metabolism and the pathogenesis of non-alcoholic steatohepatitis. Prog. Lipid Res. 2013, 52, 175–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Lin, X.; Xue, B.; Luo, J.; Gao, L.; Wang, Y.; Ou, S.; Peng, X. Impact of polyphenols combined with high-fat diet on rats’ gut microbiota. J. Funct. Foods 2016, 26, 763–771. [Google Scholar] [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A Central, Enigmatic Component of the Human Gut Microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef]
- Xie, X.; Zhang, L.; Yuan, S.; Li, H.; Zheng, C.; Xie, S.; Sun, Y.; Zhang, C.; Wang, R.; Jin, Y. Val-Val-Tyr-Pro protects against non-alcoholic steatohepatitis in mice by modulating the gut microbiota and gut-liver axis activation. J. Cell. Mol. Med. 2021, 25, 1439–1455. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | NC | HFD/OIDF-10/OIDF-50/OIDF-100 | ||
|---|---|---|---|---|
| gm | kcal | gm | kcal | |
| Casein, 80 Mesh | 200 | 800 | 200 | 800 |
| L-Cystine | 3 | 12 | 3 | 12 |
| Corn Starch | 315 | 1260 | 0 | 0 |
| Maltodextrin 10 | 35 | 140 | 125 | 500 |
| Sucrose | 350 | 1400 | 68.8 | 275.2 |
| Cellulose, BW200 | 50 | 0 | 50 | 0 |
| Soybean Oil | 25 | 225 | 25 | 225 |
| Lard | 20 | 180 | 245 | 2205 |
| Mineral Mix | 10 | 0 | 10 | 0 |
| DiCalcium Phosphate | 13 | 0 | 13 | 0 |
| Calcium Carbonate | 5.5 | 0 | 5.5 | 0 |
| Potassium Citrate,1H2O | 16.5 | 0 | 16.5 | 0 |
| Vitamin Mix | 10 | 40 | 10 | 40 |
| Choline Bitartrate | 2 | 0 | 2 | 0 |
| FD&C Blue Dye#1 | 0.05 | 0 | 0.05 | 0 |
| Total | 1055.05 | 4057 | 773.85 | 4057 |
| Composition | ||||
| gm% | kcal% | gm% | kcal% | |
| Protein | 19.2 | 20 | 26.2 | 20 |
| Carbohydrate | 67.3 | 70 | 26.3 | 20 |
| Fat | 4.3 | 10 | 34.9 | 60 |
| Total | 100 | 100 | ||
| kcal/gm | 3.85 | 5.24 | ||
| OIDF | Protein (%) | Ash (%) | Moisture (%) | TDF (%) | IDF (%) | SDF (%) |
|---|---|---|---|---|---|---|
| OIDF-10 | 1.63 ± 0.28 a | 2.95 ± 0.11 a | 6.06 ± 0.56 a | 90.06 ± 0.12 a | 89.08 ± 0.04 a | 0.98 ± 0.09 a |
| OIDF-50 | 1.66 ± 0.35 a | 3.05 ± 0.23 a | 5.87 ± 0.68 b | 89.42 ± 0.08 b | 88.63 ± 0.16 a | 0.79 ± 0.13 b |
| OIDF-100 | 1.85 ± 0.24 b | 2.98 ± 0.18 a | 5.36 ± 0.84 c | 89.81 ± 0.06 b | 89.18 ± 0.03 a | 0.63 ± 0.11 c |
| Taxonomy | NC | HD | HD-OIDF-10 | HD-OIDF-50 | HD-OIDF-100 | |
|---|---|---|---|---|---|---|
| phylum | Firmicutes | 80.32% | 80.67% | 88.25% | 93.04% | 90.64% |
| Actinobacteria | 10.75% | 17.04% | 5.65% | 2.94% | 2.37% | |
| Proteobacteria | 1.69% | 2.06% | 1.40% | 1.15% | 3.89% | |
| Bacteroidetes | 1.27% | 0.03% | 4.44% | 2.34% | 2.06% | |
| Verrucomicrobia | 5.40% | 0.09% | 0.10% | 0.01% | 0.01% | |
| Deferribacteres | 0.01% | 0.01% | 0.05% | 0.31% | 0.81% | |
| Tenericutes | 0.23% | 0.02% | 0.08% | 0.12% | 0.08% | |
| Cyanobacteria | 0.04% | 0.00% | 0.00% | 0.03% | 0.01% | |
| TM7 | 0.02% | 0.00% | 0.01% | 0.00% | 0.02% | |
| Spirochaetes | 0.01% | 0.00% | 0.00% | 0.00% | 0.00% | |
| Others | 0.26% | 0.08% | 0.03% | 0.07% | 0.12% | |
| Genus | Oscillospira | 1.38% | 8.40% | 20.25% | 5.71% | 12.58% |
| Staphylococcaceae | 21.00% | 0.47% | 1.68% | 7.87% | 7.68% | |
| Corynebacterium | 15.81% | 7.72% | 3.10% | 2.08% | 1.05% | |
| Lactobacillus | 7.75% | 4.22% | 0.10% | 1.77% | 0.51% | |
| Clostridiaceae | 2.51% | 2.21% | 1.27% | 3.63% | 3.43% | |
| Jeotgalicoccus | 4.18% | 1.67% | 1.06% | 0.95% | 1.86% | |
| Ruminococcus | 0.11% | 1.65% | 3.62% | 0.91% | 1.62% | |
| Allobaculum | 0.58% | 5.88% | 0.26% | 0.06% | 0.37% | |
| Desulfovibrio | 1.41% | 0.85% | 0.33% | 0.64% | 3.18% | |
| Coprococcus | 0.74% | 1.01% | 2.69% | 0.82% | 0.90% | |
| Others | 44.53% | 65.91% | 65.64% | 75.57% | 66.82% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, H.; Zhang, Y.; Swallah, M.S.; Wang, S.; Zhang, J.; Fang, J.; Lu, J.; Yu, H. Structural Characteristics of Insoluble Dietary Fiber from Okara with Different Particle Sizes and Their Prebiotic Effects in Rats Fed High-Fat Diet. Foods 2022, 11, 1298. https://doi.org/10.3390/foods11091298
Fan H, Zhang Y, Swallah MS, Wang S, Zhang J, Fang J, Lu J, Yu H. Structural Characteristics of Insoluble Dietary Fiber from Okara with Different Particle Sizes and Their Prebiotic Effects in Rats Fed High-Fat Diet. Foods. 2022; 11(9):1298. https://doi.org/10.3390/foods11091298
Chicago/Turabian StyleFan, Hongliang, Ying Zhang, Mohammed Sharif Swallah, Sainan Wang, Jiarui Zhang, Jiaqi Fang, Jiahong Lu, and Hansong Yu. 2022. "Structural Characteristics of Insoluble Dietary Fiber from Okara with Different Particle Sizes and Their Prebiotic Effects in Rats Fed High-Fat Diet" Foods 11, no. 9: 1298. https://doi.org/10.3390/foods11091298
APA StyleFan, H., Zhang, Y., Swallah, M. S., Wang, S., Zhang, J., Fang, J., Lu, J., & Yu, H. (2022). Structural Characteristics of Insoluble Dietary Fiber from Okara with Different Particle Sizes and Their Prebiotic Effects in Rats Fed High-Fat Diet. Foods, 11(9), 1298. https://doi.org/10.3390/foods11091298