


Combined Effects of BEIIb and SSIIa Alleles on Amylose Contents, Starch Fine Structures and Physicochemical Properties of Indica Rice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
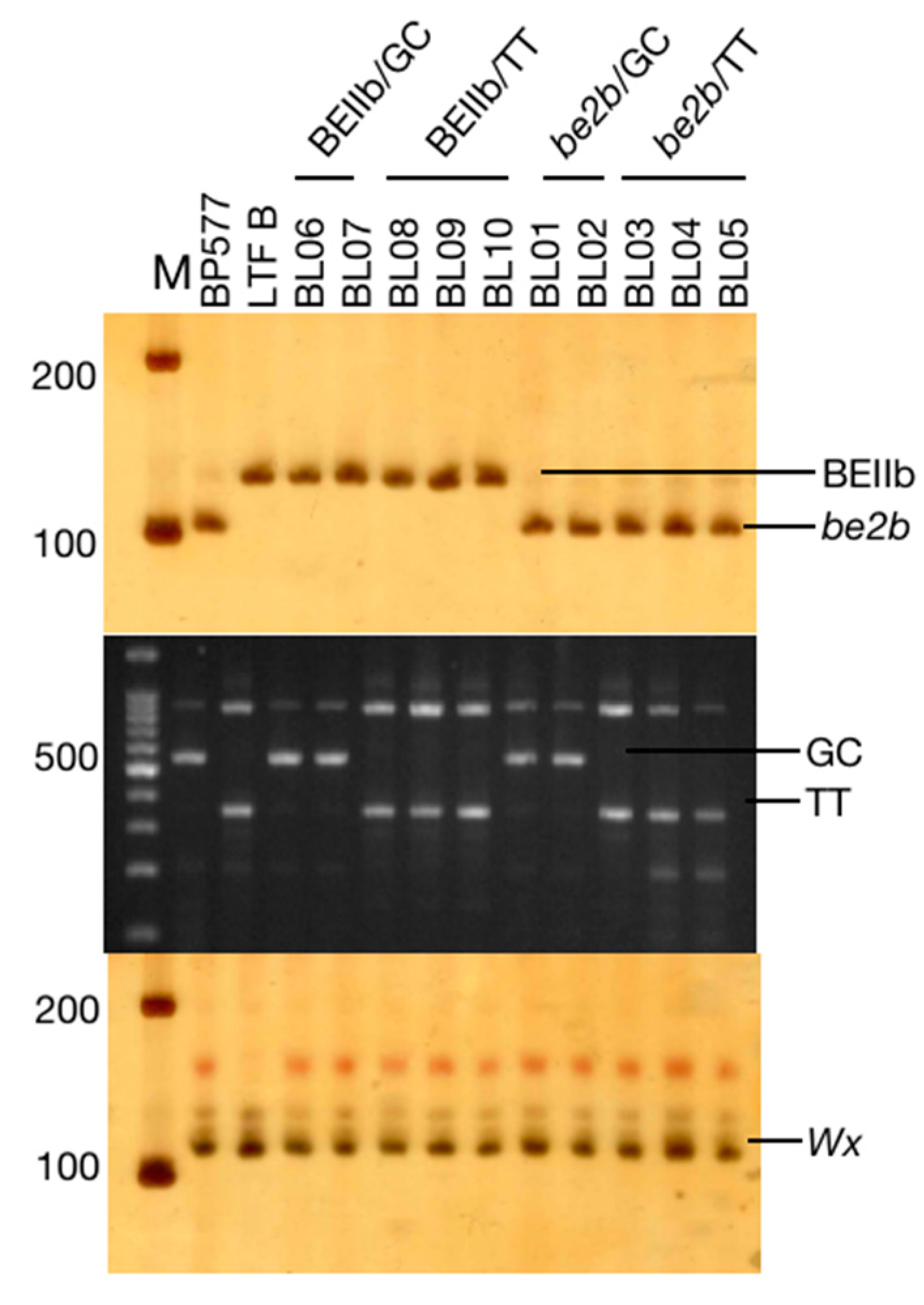
2.2. Genotyping of Breeding Lines
2.3. Starch Isolation and Debranching
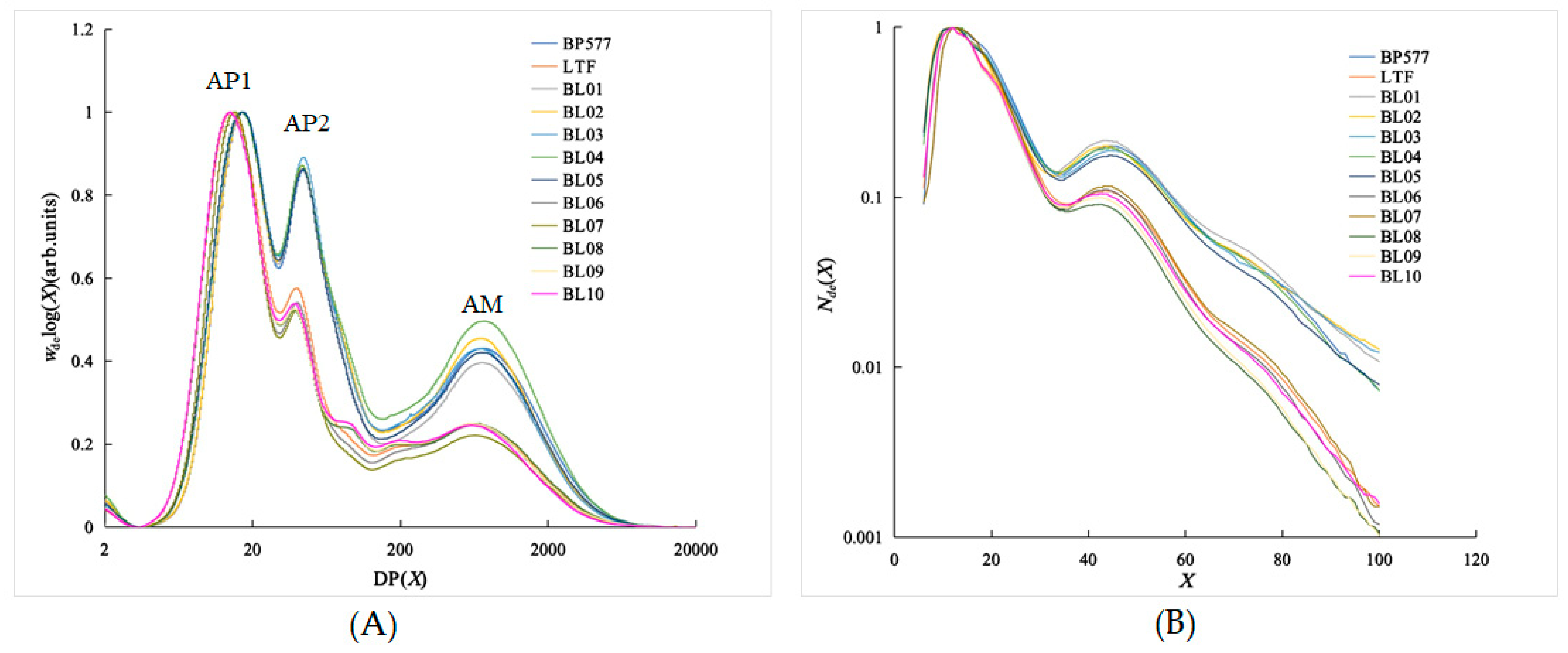
2.4. Size-Exclusion Chromatography (SEC)
2.5. Amylose Content Measurement
2.6. FACE Analysis
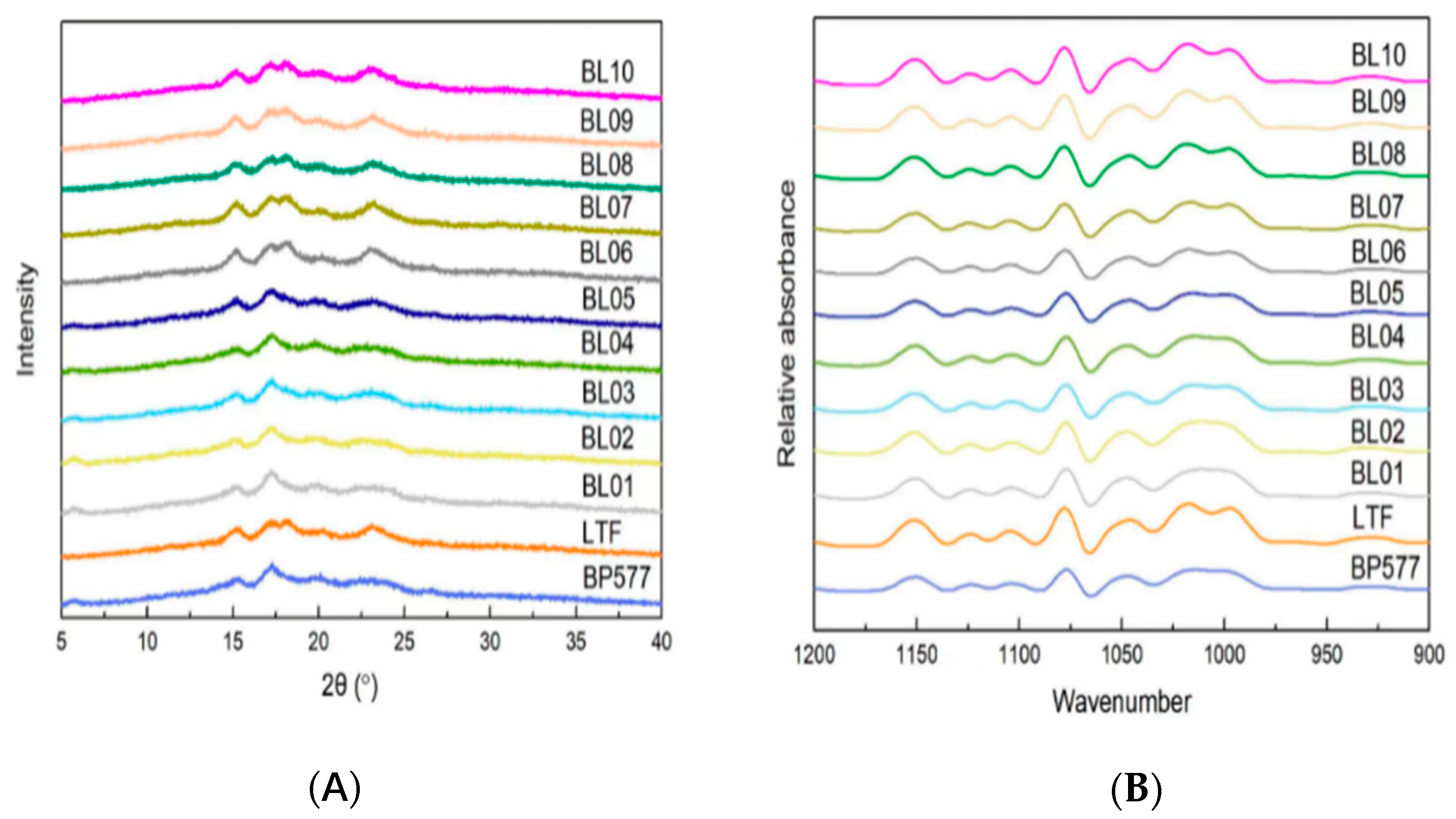
2.7. X-ray Diffraction
2.8. Attenuated Total Reflectance-Fourier Transform Infrared Spectroscopy
2.9. RVA Pasting Viscosity
2.10. DSC Thermal (Gelatinization) Properties
2.11. Statistical Analysis
3. Results and Discussion
3.1. Genotyping of The Breeding Lines
3.2. SEC Chain-Length Distributions of the Debranched Starch
3.3. Crystalline Structure
3.4. Thermal Properties
3.5. Pasting Viscosities
3.6. Relationship between Different SSIIa/BEIIb Genotypes
3.7. Relationships between Fine Structure and Physicochemical Properties
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AC | Amylose content |
| BD | Breakdown |
| BE | Branching enzyme |
| CLD | Chain length distribution |
| CPV | Cold paste viscosity |
| CS | Consistency viscosity |
| CEQ | Cooking and eating quality |
| DP | Degree of polymerization |
| FACE | Fluorophore-assisted carbohydrate electrophoresis |
| GT | Gelatinization temperature |
| HPV | Hot paste viscosity |
| PV | Peak viscosity |
| RC | relative crystallinity |
| RIL | Recombinant inbred line |
| RVA | Rapid Visco-Analyzer |
| RVU | Rapid Visco Unit |
| SB | Setback |
| SEC | Size-exclusion chromatography |
| SNP | Single nucleotide polymorphism |
| SS | Soluble starch synthase |
| Tc | conclusion temperature |
| To | Onset temperature |
| Tp | peak temperature |
| Wx | Waxy |
| ΔH | Enthalpy of gelatinization |
References
- Bao, J.; Shen, S.; Sun, M.; Corke, H. Analysis of genotypic diversity in the starch physicochemical properties of nonwaxy rice: Apparent amylose content, pasting viscosity and gel texture. Starch-Starke 2006, 58, 259–267. [Google Scholar] [CrossRef]
- Singh, N.; Kaur, L.; Sandhu, K.S.; Kaur, J.; Nishinari, K. Relationships between physicochemical, morphological, thermal, rheological properties of rice starches. Food Hydrocoll. 2006, 20, 532–542. [Google Scholar] [CrossRef]
- Tester, R.F.; Karkalas, J.; Qi, X. Starch—Composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Witt, T.; Doutch, J.; Gilbert, E.P.; Gilbert, R.G. Relations between molecular, crystalline, and lamellar structures of amylopectin. Biomacromolecules 2012, 13, 4273–4282. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.-S.; Ryoo, N.; Hahn, T.-R.; Walia, H.; Nakamura, Y. Starch biosynthesis in cereal endosperm. Plant Physiol. Biochem. 2010, 48, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.S. Towards understanding of the genetic and molecular basis of eating and cooking quality of rice. Cereal Foods World 2012, 57, 148–156. [Google Scholar] [CrossRef]
- Hirano, H.Y.; Eiguchi, M.; Sano, Y. A single base change altered the regulation of the Waxy gene at the posttranscriptional level during the domestication of rice. Mol. Biol. Evol. 1998, 15, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Ayres, N.M.; McClung, A.M.; Larkin, P.D.; Bligh, H.F.J.; Jones, C.A.; Park, W.D. Microsatellites and a single-nucleotide polymorphism differentiate apparent amylose classes in an extended pedigree of US rice germ plasm. Theor. Appl. Genet. 1997, 94, 773–781. [Google Scholar] [CrossRef]
- Bao, J.S.; Corke, H.; Sun, M. Microsatellites, single nucleotide polymorphisms and a sequence tagged site in starch-synthesizing genes in relation to starch physicochemical properties in nonwaxy rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 113, 1185–1196. [Google Scholar] [CrossRef]
- Chen, M.-H.; Bergman, C.J.; Pinson, S.R.; Fjellstrom, R.G. Waxy gene haplotypes: Associations with pasting properties in an international rice germplasm collection. J. Cereal Sci. 2008, 48, 781–788. [Google Scholar] [CrossRef]
- Umemoto, T.; Aoki, N. Single-nucleotide polymorphisms in rice starch synthase IIa that alter starch gelatinisation and starch association of the enzyme. Funct. Plant Biol. 2005, 32, 763–768. [Google Scholar] [CrossRef]
- Waters, D.L.E.; Henry, R.J.; Reinke, R.F.; Fitzgerald, M.A. Gelatinization temperature of rice explained by polymorphisms in starch synthase. Plant Biotechnol. J. 2006, 4, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.S.; Corke, H.; Sun, M. Nucleotide diversity in starch synthase IIa and validation of single nucleotide polymorphisms in relation to starch gelatinization temperature and other physicochemical properties in rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 113, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Xiao, P.; Hiratsuka, M.; Sun, M.; Umemoto, T. Granule-bound SSIIa protein content and its relationship with amylopectin structure and gelatinization temperature of rice starch. Starch-Starke 2009, 61, 431–437. [Google Scholar] [CrossRef]
- Nakamura, Y.; Francisco, P.B.; Hosaka, Y.; Sato, A.; Sawada, T.; Kubo, A.; Fujita, N. Essential amino acids of starch synthase IIa differentiate amylopectin structure and starch quality between japonica and indica rice varieties. Plant Mol. Biol. 2005, 58, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Hu, Y.; Deng, G.; Dai, G.; Bao, J. The origin of the A/G single nucleotide polymorphism of starch synthase IIa in rice and its relation to gelatinization temperature. Cereal Chem. 2021, 99, 275–285. [Google Scholar] [CrossRef]
- Fei, Y.; Yang, J.; Wang, F.; Fan, F.; Wenqi, L.I.; Wang, J.; Yang, X.U.; Zhu, J.; Zhong, W. Production of two elite glutinous rice varieties by editing Wx gene. Rice Sci. 2019, 26, 118–127. [Google Scholar]
- Nishi, A.; Nakamura, Y.; Satoh, T.H. Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar] [CrossRef]
- Tappiban, P.; Hu, Y.; Deng, J.; Zhao, J.; Ying, Y.; Zhang, Z.; Xu, F.; Bao, J. Relative importance of branching enzyme isoforms in determining starch fine structure and physicochemical properties of indica rice. Plant Mol. Biol. 2021, 108, 399–412. [Google Scholar] [CrossRef]
- Bao, J.; Zhou, X.; Xu, F.; He, Q.; Park, Y.-J. Genome-wide association study of the resistant starch content in rice grains. Starch-Starke 2017, 69, 1600343. [Google Scholar] [CrossRef]
- Itoh, Y.; Crofts, N.; Abe, M.; Hosaka, Y.; Fujita, N. Characterization of the endosperm starch and the pleiotropic effects of biosynthetic enzymes on their properties in novel mutant rice lines with high resistant starch and amylose content. Plant Sci. 2017, 258, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Kubo, A.; Akdogan, G.; Nakaya, M.; Shojo, A.; Kitamura, S. Structure, physical, and digestive properties of starch from wx ae double-mutant rice. J. Agric. Food Chem. 2010, 58, 4463–4469. [Google Scholar] [CrossRef]
- Wang, K.; Hasjim, J.; Wu, A.C.; Li, E.; Henry, R.J.; Gilbert, R.G. Roles of GBSSI and SSIIa in determining amylose fine structure. Carbohydr. Polym. 2015, 127, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Ida, T.; Crofts, N.; Miura, S.; Matsushima, R.; Fujita, N. Structure and properties of starch in rice double mutants lacking starch synthase (SS) IIa and starch branching enzyme (BE) IIb. J. Appl. Glycosci. 2021, 68, 31–39. [Google Scholar] [CrossRef]
- Doyle, J. DNA protocols for plants CTAB total DNA isolation. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A., Eds.; Springer: Berlin, Germany, 1991; pp. 283–293. [Google Scholar]
- Zhang, Z.; Hu, Y.; Zhao, J.; Zhang, Y.; Ying, Y.; Xu, F.; Bao, J. The role of different Wx and BEIIb allele combinations on fine structures and functional properties of indica rice starches. Carbohydr. Polym. 2021, 278, 118972. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Lu, Y.; Shao, Y.; Zhang, G.; Xiao, P.; Shen, S.; Corke, H.; Bao, J. Molecular marker assisted selection for improvement of the eating, cooking and sensory quality of rice (Oryza sativa L.). J. Cereal Sci. 2009, 51, 159–164. [Google Scholar] [CrossRef]
- Syahariza, Z.A.; Li, E.; Hasjim, J. Extraction and dissolution of starch from cereal grains for accurate structural analysis. Carbohydr. Polym. 2010, 82, 14–20. [Google Scholar] [CrossRef]
- Castro, J.V.; Ward, R.M.; Gilbert, A.R.G.; Fitzgerald, M.A. Measurement of the molecular weight distribution of debranched starch. Biomacromolecules 2005, 6, 2260–2270. [Google Scholar] [CrossRef]
- Wu, A.C.; Li, E.; Gilbert, R.G. Exploring extraction/dissolution procedures for analysis of starch chain-length distributions. Carbohydr. Polym. 2014, 114, 36–42. [Google Scholar] [CrossRef]
- Hanashiro, I.; Abe, J.-I.; Hizukuri, S. A periodic distribution of the chain length of amylopectin as revealed by high-performance anion-exchange chromatography. Carbohydr. Res. 1996, 283, 151–159. [Google Scholar] [CrossRef]
- Zhang, Z.; Fan, X.; Ma, H.; Li, C.; Li, E.; Gilbert, R.G. Characterization of the baking-induced changes in starch molecular and crystalline structures in sugar-snap cookies. Carbohydr. Polym. 2020, 256, 117518. [Google Scholar] [CrossRef] [PubMed]
- Sevenou, O.; Hill, S.; Farhat, I.; Mitchell, J. Organisation of the external region of the starch granule as determined by infrared spectroscopy. Int. J. Biol. Macromol. 2002, 31, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Sun, M.; Corke, H. Analysis of genotypic diversity in starch thermal and retrogradation properties in nonwaxy rice. Carbohydr. Polym. 2007, 67, 174–181. [Google Scholar] [CrossRef]
- Ahmed, S.; Ru, W.; Han, H.; Cheng, L.; Bian, X.; Li, G.; Jin, L.; Wu, P.; Bao, J. Fine molecular structure and its effects on physicochemical properties of starches in potatoes grown in two locations. Food Hydrocoll. 2019, 97, 105172. [Google Scholar] [CrossRef]
- Tappiban, P.; Ying, Y.; Pang, Y.; Sraphet, S.; Srisawad, N.; Smith, D.R.; Wu, P.; Triwitayakorn, K.; Bao, J. Gelatinization, pasting and retrogradation properties and molecular fine structure of starches from seven cassava cultivars. Int. J. Biol. Macromol. 2020, 150, 831–838. [Google Scholar] [CrossRef]
- Lin, L.; Huang, J.; Zhang, L.; Liu, Q.; Wei, C. Effects of inhibition of starch branching enzymes on starch ordered structure and component accumulation in developing kernels of rice. J. Cereal Sci. 2019, 91, 102884. [Google Scholar] [CrossRef]
- Butardo, J.V.M.; Daygon, V.D.; Colgrave, M.L.; Campbell, P.M.; Resurreccion, A.; Cuevas, R.P.; Jobling, S.A.; Tetlow, I.; Rahman, S.; Morell, M.; et al. Biomolecular analyses of starch and starch granule proteins in the high-amylose rice mutant Goami 2. J. Agric. Food Chem. 2012, 60, 11576–11585. [Google Scholar] [CrossRef]
- Butardo, V.M.; Fitzgerald, M.A.; Bird, A.R.; Gidley, M.J.; Flanagan, B.M.; Larroque, O.; Resurreccion, A.P.; Laidlaw, H.K.C.; Jobling, S.A.; Morell, M.K.; et al. Impact of down-regulation of starch branching enzyme IIb in rice by artificial microRNA- and hairpin RNA-mediated RNA silencing. J. Exp. Bot. 2011, 62, 4927–4941. [Google Scholar] [CrossRef] [Green Version]
- Fuwa, H.; Inouchi, N.; Glover, D.V.; Fujita, S.; Sugihara, M.; Yoshioka, S.; Yamada, K.; Sugimoto, Y. Structural and physicochemical properties of endosperm starches possessing different alleles at the amylose-extender and Waxy locus in maize (Zea mays L.). Starch-Starke 1999, 51, 147–151. [Google Scholar] [CrossRef]
- Crofts, N.; Abe, N.; Oitome, N.F.; Matsushima, R.; Hayashi, M.; Tetlow, I.J.; Emes, M.J.; Nakamura, Y.; Fujita, N. Amylopectin biosynthetic enzymes from developing rice seed form enzymatically active protein complexes. J. Exp. Bot. 2015, 66, 4469–4482. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Pang, Y.; Bao, J. Expression profiles and protein complexes of starch biosynthetic enzymes from white-core and waxy mutants induced from high amylose Indica rice. Rice Sci. 2020, 27, 152–161. [Google Scholar]
- Jane, J.; Chen, Y.Y.; Lee, L.F.; McPherson, A.E.; Wong, K.S.; Radosavljevic, M.; Kasemsuwan, T. Effects of amylopectin branch chain length and amylose content on the gelatinization and pasting properties of starch. Cereal Chem. 1999, 76, 629–637. [Google Scholar] [CrossRef]
- Tao, K.; Li, C.; Yu, W.; Gilbert, R.G.; Li, E. How amylose molecular fine structure of rice starch affects functional properties. Carbohydr. Polym. 2018, 204, 24–31. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Genotype | XAP1 | XAP2 | XAM | hAM | hAP2/hAP1 | AC (%) |
|---|---|---|---|---|---|---|---|
| BP577 | GC/be2b | 17.26 ± 0.10 a | 44.04 ± 0.08 ab | 729.47 ± 4.76 a | 0.42 ± 0.01 cd | 0.86 ± 0.00 b | 33.16 ± 0.011 bc |
| LTFB | TT/BEIIb | 14.09 ± 0.19 c | 39.68 ± 0.14 e | 580.61 ± 12.17 e | 0.25 ± 0.00 ef | 0.58 ± 0.00 c | 26.85 ± 0.007 e |
| BL01 | GC/be2b | 17.29 ± 0.07 a | 43.89 ± 0.22 bc | 710.71 ± 4.62 a | 0.40 ± 0.01 d | 0.86 ± 0.00 b | 32.27 ± 0.002 c |
| BL02 | GC/be2b | 17.10 ± 0.06 a | 43.52 ± 0.15 c | 718.95 ± 24.96 a | 0.45 ± 0.00 b | 0.87 ± 0.00 b | 33.84 ± 0.002 ab |
| BL03 | TT/be2b | 17.19 ± 0.10 a | 44.42 ± 0.00 a | 704.57 ± 1.52 ab | 0.44 ± 0.01 bc | 0.89 ± 0.00 a | 32.94 ± 0.004 bc |
| BL04 | TT/be2b | 16.87 ± 0.03 a | 43.60 ± 0.22 bc | 743.92 ± 3.25 a | 0.48 ± 0.02 a | 0.88 ± 0.00 ab | 34.84 ± 0.009 a |
| BL05 | TT/be2b | 17.06 ± 0.03 a | 43.97 ± 0.15 abc | 698.55 ± 7.54 abc | 0.42 ± 0.00 bcd | 0.86 ± 0.00 b | 32.31 ± 0.003 c |
| BL06 | GC/BEIIb | 15.13 ± 0.06 b | 40.50 ± 0.14 d | 639.70 ± 2.72 d | 0.24 ± 0.01 ef | 0.54 ± 0.00 d | 28.26 ± 0.005 d |
| BL07 | GC/BEIIb | 15.16 ± 0.09 b | 40.23 ± 0.14 d | 642.54 ± 10.95 d | 0.22 ± 0.00 f | 0.53 ± 0.00 d | 26.82 ± 0.000 e |
| BL08 | TT/BEIIb | 14.65 ± 0.54 b | 38.67 ± 0.20 f | 651.70 ± 33.37 bcd | 0.25 ± 0.00 e | 0.53 ± 0.00 d | 27.78 ± 0.001 de |
| BL09 | TT/BEIIb | 14.01 ± 0.00 c | 38.60 ± 0.00 f | 643.96 ± 41.11 cd | 0.25 ± 0.00 e | 0.53 ± 0.00 d | 27.35 ± 0.003 de |
| BL10 | TT/BEIIb | 14.09 ± 0.03 c | 39.00 ± 0.13 f | 617.15 ± 11.76 de | 0.24 ± 0.00 ef | 0.53 ± 0.01 d | 27.18 ± 0.009 de |
| Sample | Genotype | fa | fb1 | fb2 | fb3 | |
|---|---|---|---|---|---|---|
| BP577 | GC/be2b | 21.36 ± 0.17 fg | 42.53 ± 0.04 f | 11.55 ± 0.09 c | 24.56 ± 0.05 bc | 26.25 ± 0.03 c |
| LTFB | TT/BEIIb | 27.27 ± 0.25 c | 46.59 ± 0.32 c | 11.20 ± 0.07 d | 14.95 ± 0.51 ef | 21.73 ± 0.22 f |
| BL01 | GC/be2b | 21.05 ± 0.25 gh | 41.74 ± 0.50 g | 11.34 ± 0.07 d | 25.76 ± 0.82 a | 26.94 ± 0.35 a |
| BL02 | GC/be2b | 21.73 ± 0.21 ef | 41.74 ± 0.17 g | 11.01 ± 0.04 e | 25.52 ± 0.34 ab | 26.76 ± 0.15 ab |
| BL03 | TT/be2b | 20.50 ± 0.08 hi | 43.29 ± 0.40 e | 12.31 ± 0.11 a | 23.90 ± 0.42 c | 26.45 ± 0.16 abc |
| BL04 | TT/be2b | 20.29 ± 0.28 i | 43.35 ± 0.07 e | 12.01 ± 0.08 b | 24.35 ± 0.27 c | 26.43 ± 0.11 bc |
| BL05 | TT/be2b | 22.05 ± 0.61 de | 44.06 ± 0.59 d | 11.87 ± 0.09 b | 21.02 ± 1.11 d | 25.34 ± 0.54 d |
| BL06 | GC/BEIIb | 22.43 ± 0.14 d | 52.15 ± 0.34 a | 10.69 ± 0.01 fg | 14.73 ± 0.47 ef | 21.96 ± 0.19 ef |
| BL07 | GC/BEIIb | 22.09 ± 0.31 de | 51.74 ± 0.20 a | 10.82 ± 0.13 f | 15.35 ± 0.03 e | 22.29 ± 0.04 e |
| BL08 | TT/BEIIb | 29.17 ± 0.24 a | 47.68 ± 0.46 b | 10.49 ± 0.03 h | 12.66 ± 0.73 h | 20.57 ± 0.27 h |
| BL09 | TT/BEIIb | 28.71 ± 0.15 ab | 47.46 ± 0.03 b | 10.64 ± 0.05 gh | 13.19 ± 0.14 gh | 20.80 ± 0.08 gh |
| BL10 | TT/BEIIb | 28.16 ± 0.05 b | 47.26 ± 0.08 cb | 10.64 ± 0.05 gh | 13.93 ± 0.08 gf | 21.17 ± 0.03 g |
| Sample | Genotype | RC (%) | 1045/1022 | 1022/995 |
|---|---|---|---|---|
| BP577 | GC/be2b | 21.68 ± 0.33 cd | 0.783 ± 0.010 b | 0.998 ± 0.011 de |
| LTFB | TT/BEIIb | 23.16 ± 0.01 b | 0.654 ± 0.005 de | 1.060 ± 0.011 a–e |
| BL01 | GC/be2b | 20.51 ± 0.13 e | 0.812 ± 0.005 a | 0.937 ± 0.011 e |
| BL02 | GC/be2b | 20.75 ± 0.27 de | 0.782 ± 0.006 ab | 1.020 ± 0.011 cde |
| BL03 | TT/be2b | 21.07 ± 0.16 cde | 0.778 ± 0.002 ab | 0.925 ± 0.011 e |
| BL04 | TT/be2b | 20.89 ± 0.34 de | 0.753 ± 0.015 bc | 0.998 ± 0.011 de |
| BL05 | TT/be2b | 21.88 ± 0.44 c | 0.769 ± 0.001 b | 0.967 ± 0.011 de |
| BL06 | GC/BEIIb | 24.27 ± 0.17 a | 0.722 ± 0.026 c | 1.102 ± 0.011 a–d |
| BL07 | GC/BEIIb | 24.22 ± 0.21 a | 0.742 ± 0.008 bc | 1.042 ± 0.011 b–e |
| BL08 | TT/BEIIb | 23.38 ± 0.14 ab | 0.678 ± 0.007 d | 1.164 ± 0.011 ab |
| BL09 | TT/BEIIb | 23.89 ± 0.25 ab | 0.652 ± 0.012 de | 1.186 ± 0.011 a |
| BL10 | TT/BEIIb | 23.09 ± 0.21 b | 0.634 ± 0.005 e | 1.140 ± 0.011 abc |
| Sample | Genotype | To | Tp | Tc | ΔH |
|---|---|---|---|---|---|
| BP577 | GC/be2b | 74.92 ± 0.06 b | 83.06 ± 0.58 abc | 92.96 ± 0.20 b | 8.08 ± 0.24 bcd |
| LTFB | TT/BEIIb | 61.24 ± 0.10 e | 69.40 ± 0.08 f | 77.46 ± 0.06 f | 7.14 ± 0.12 cd |
| BL01 | GC/be2b | 74.73 ± 0.12 b | 83.70 ± 0.13 a | 93.96 ± 0.00 a | 8.96 ± 0.35 ab |
| BL02 | GC/be2b | 74.01 ± 0.19 c | 82.31 ± 0.44 c | 92.58 ± 0.68 b | 8.37 ± 0.12 abc |
| BL03 | TT/be2b | 73.74 ± 0.22 c | 82.52 ± 0.03 bc | 90.43 ± 0.39 c | 8.41 ± 0.22 abc |
| BL04 | TT/be2b | 69.32 ± 0.11 d | 80.03 ± 0.07 d | 88.99 ± 0.25 d | 5.22 ± 0.06 e |
| BL05 | TT/be2b | 75.52 ± 0.08 a | 83.24 ± 0.10 ab | 89.82 ± 0.35 cd | 6.78 ± 0.52 d |
| BL06 | GC/BEIIb | 73.66 ± 0.26 c | 78.70 ± 0.24 e | 83.42 ± 0.28 e | 9.38 ± 0.34 a |
| BL07 | GC/BEIIb | 73.97 ± 0.19 c | 78.37 ± 0.16 e | 83.67 ± 0.07 e | 9.58 ± 1.22 a |
| BL08 | TT/BEIIb | 58.31 ± 0.06 g | 65.04 ± 0.07 g | 73.09 ± 0.13 g | 6.88 ± 0.07 d |
| BL09 | TT/BEIIb | 58.24 ± 0.25 g | 65.32 ± 0.29 g | 73.56 ± 0.46 g | 7.27 ± 0.11 cd |
| BL10 | TT/BEIIb | 58.81 ± 0.00 f | 65.58 ± 0.22 g | 73.84 ± 0.08 g | 6.92 ± 0.03 d |
| Sample | Genotype | PV | HPV | CPV | BD | SB | CS |
|---|---|---|---|---|---|---|---|
| BP577 | GC/be2b | 115.75 ± 0.00 fg | 105.88 ± 0.38 f | 142.55 ± 2.13 de | 9.88 ± 0.38 cde | 26.80 ± 2.13 ef | 36.67 ± 1.75 bc |
| LTFB | TT/BEIIb | 275.38 ± 6.38 a | 246.88 ± 3.63 a | 357.29 ± 10.04 a | 28.50 ± 2.75 b | 81.92 ± 3.67 bc | 110.42 ± 6.42 a |
| BL01 | GC/be2b | 116.71 ± 2.54 fg | 110.46 ± 4.79 ef | 142.21 ± 13.38 de | 6.25 ± 2.25 de | 25.50 ± 10.84 ef | 31.75 ± 8.59 c |
| BL02 | GC/be2b | 95.65 ± 4.15 h | 89.59 ± 4.67 g | 116.75 ± 9.67 e | 6.07 ± 0.52 de | 21.10 ± 5.52 f | 27.17 ± 5.01 c |
| BL03 | TT/be2b | 126.42 ± 1.09 ef | 119.92 ± 0.50 de | 158.55 ± 1.38 d | 6.50 ± 0.59 de | 32.13 ± 2.46 ef | 38.63 ± 1.88 bc |
| BL04 | TT/be2b | 104.46 ± 0.96 gh | 100.92 ± 2.25 fg | 149.96 ± 3.88 d | 3.54 ± 1.29 e | 45.50 ± 2.92 de | 49.04 ± 1.63 bc |
| BL05 | TT/be2b | 139.96 ± 0.54 e | 131.96 ± 0.13 d | 197.09 ± 0.17 c | 8.01 ± 0.42 cde | 57.13 ± 0.38 d | 65.13 ± 0.04 b |
| BL06 | GC/BEIIb | 231.88 ± 4.63 b | 164.71 ± 4.13 c | 300.13 ± 8.88 b | 67.17 ± 8.75 a | 68.25 ± 4.25 cd | 135.42 ± 13.00 a |
| BL07 | GC/BEIIb | 241.50 ± 4.08 b | 178.92 ± 4.67 b | 306.96 ± 12.04 b | 62.59 ± 8.75 a | 65.46 ± 7.96 cd | 128.05 ± 16.71 a |
| BL08 | TT/BEIIb | 183.38 ± 7.13 d | 166.46 ± 6.96 c | 282.71 ± 8.46 b | 16.92 ± 0.16 bcd | 99.34 ± 1.34 ab | 116.25 ± 1.50 a |
| BL09 | TT/BEIIb | 185.42 ± 3.83 cd | 167.79 ± 1.04 bc | 294.92 ± 21.75 b | 17.63 ± 4.88 bcd | 109.51 ± 17.92 a | 127.13 ± 22.79 a |
| BL10 | TT/BEIIb | 199.59 ± 9.58 c | 179.67 ± 6.00 b | 285.42 ± 0.00 b | 19.92 ± 3.59 bc | 85.84 ± 9.59 bc | 105.75 ± 6.00 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Zhang, Y.; Yu, S.; Deng, G.; Dai, G.; Bao, J. Combined Effects of BEIIb and SSIIa Alleles on Amylose Contents, Starch Fine Structures and Physicochemical Properties of Indica Rice. Foods 2023, 12, 119. https://doi.org/10.3390/foods12010119
Hu Y, Zhang Y, Yu S, Deng G, Dai G, Bao J. Combined Effects of BEIIb and SSIIa Alleles on Amylose Contents, Starch Fine Structures and Physicochemical Properties of Indica Rice. Foods. 2023; 12(1):119. https://doi.org/10.3390/foods12010119
Chicago/Turabian StyleHu, Yaqi, Yanni Zhang, Shouwu Yu, Guofu Deng, Gaoxing Dai, and Jinsong Bao. 2023. "Combined Effects of BEIIb and SSIIa Alleles on Amylose Contents, Starch Fine Structures and Physicochemical Properties of Indica Rice" Foods 12, no. 1: 119. https://doi.org/10.3390/foods12010119
APA StyleHu, Y., Zhang, Y., Yu, S., Deng, G., Dai, G., & Bao, J. (2023). Combined Effects of BEIIb and SSIIa Alleles on Amylose Contents, Starch Fine Structures and Physicochemical Properties of Indica Rice. Foods, 12(1), 119. https://doi.org/10.3390/foods12010119