


Freshwater Clam Extract Attenuates Indomethacin-Induced Gastric Damage In Vitro and In Vivo

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
2.2. Preparation of Water Extract of Freshwater Clam (WEC) and Analysis of Its Amino Acid Composition
2.3. Cell Culture and Cell Viability Analysis
2.4. Reactive Oxygen Species (ROS) Production Analysis
2.5. Animal Experiment
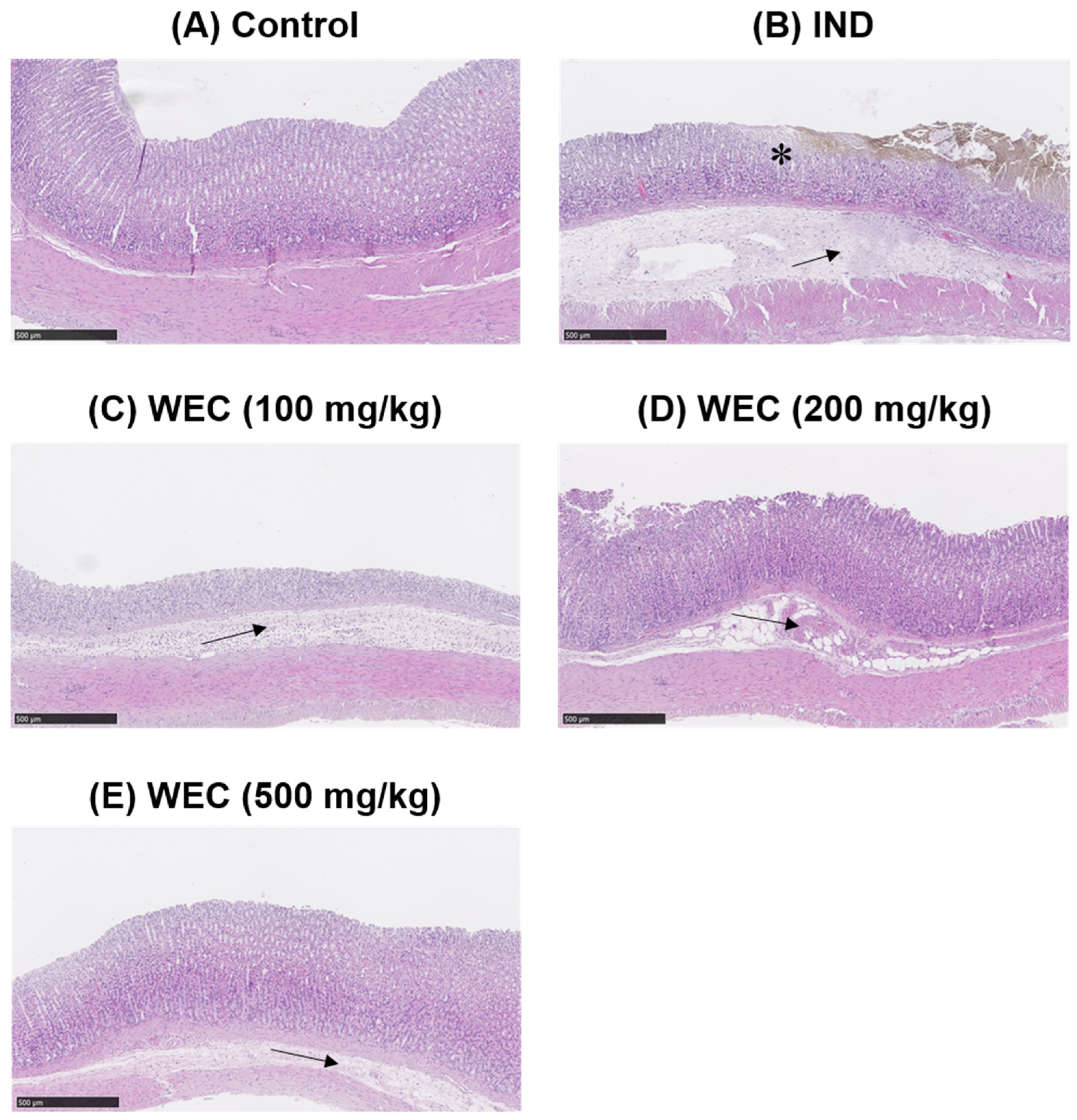
2.6. Ulcer Area and Histopathological Assessment
2.7. Analysis of the Contents of Glutathione (GSH) and Prostaglandin E2 (PGE2) in Gastric Mucosa
2.8. Protein Expression Analysis
2.9. Statistical Analysis
3. Results and Discussion
3.1. Amino Acid Composition of WEC and Its Hydrolysate
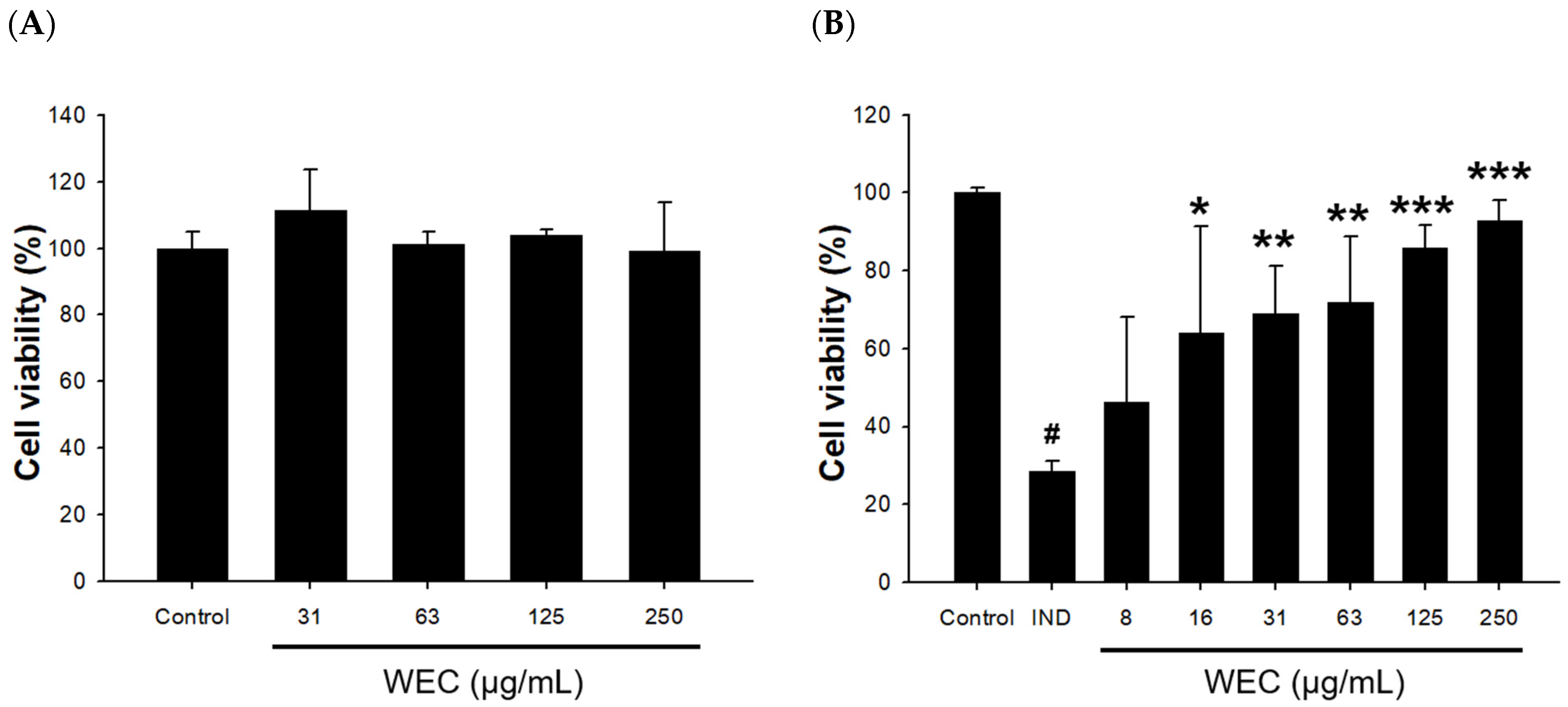
3.2. Effects of WEC on Cell Viability and IND-Induced Cytotoxicity in Rat Gastric Mucosa RGM-1 Cells
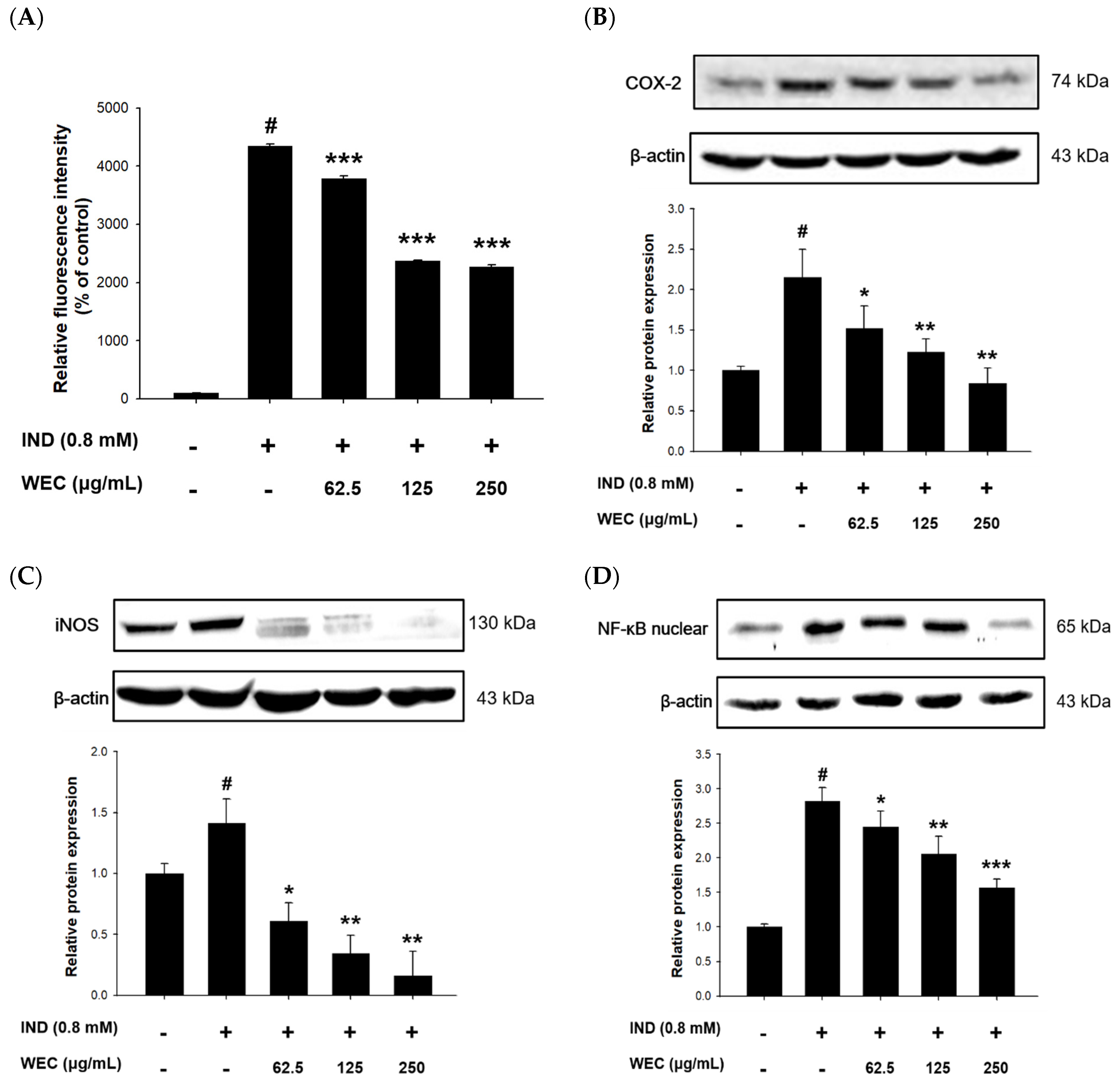
3.3. Effects of WEC on IND-Induced Oxidative Stress and Associated Protein Levels
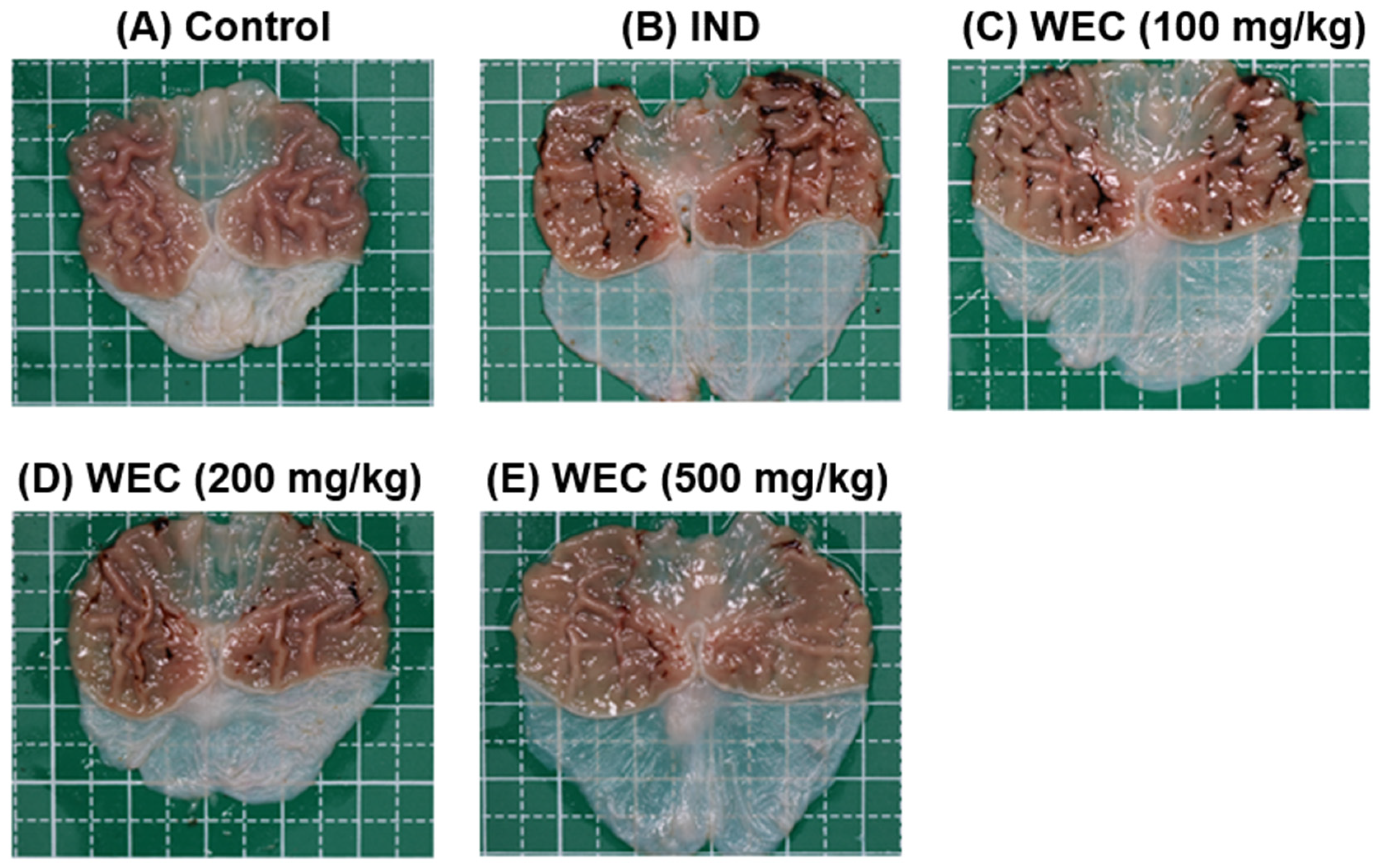
3.4. Effects of WEC on IND-Induced Gastric Ulcers in Wistar Rats
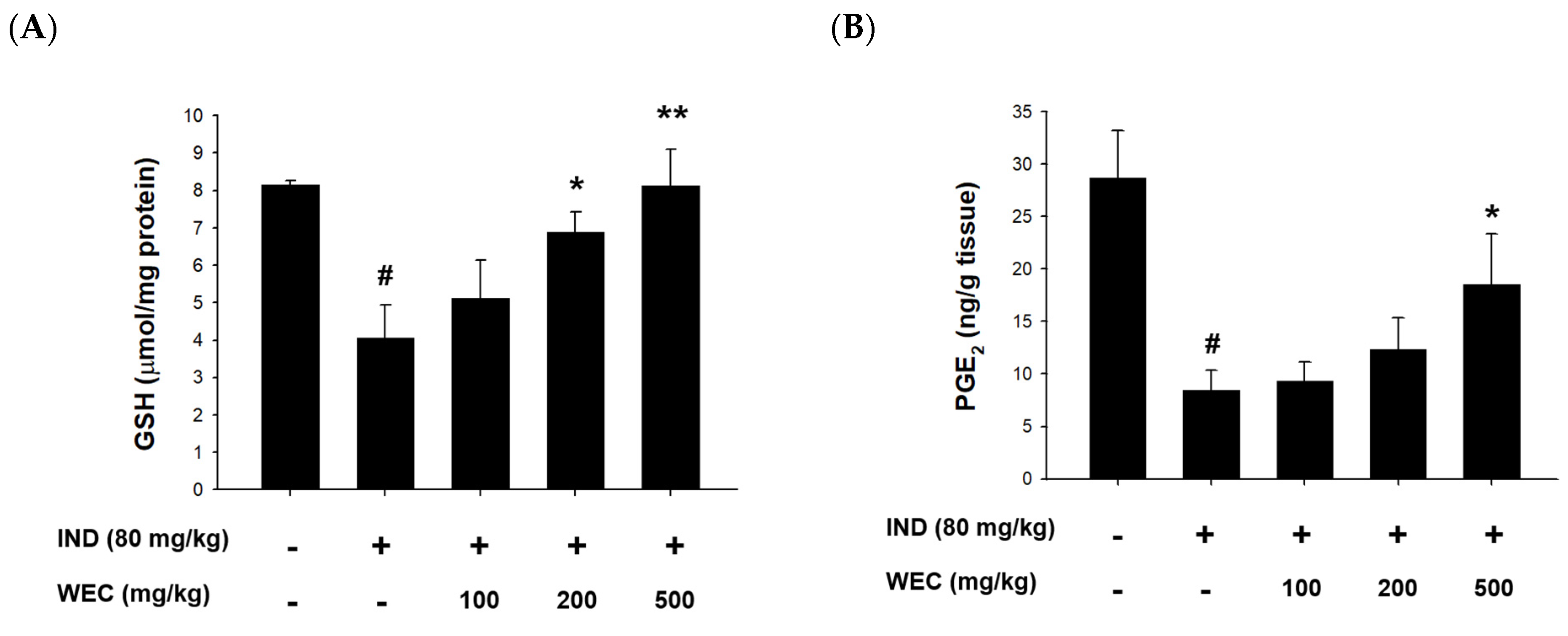
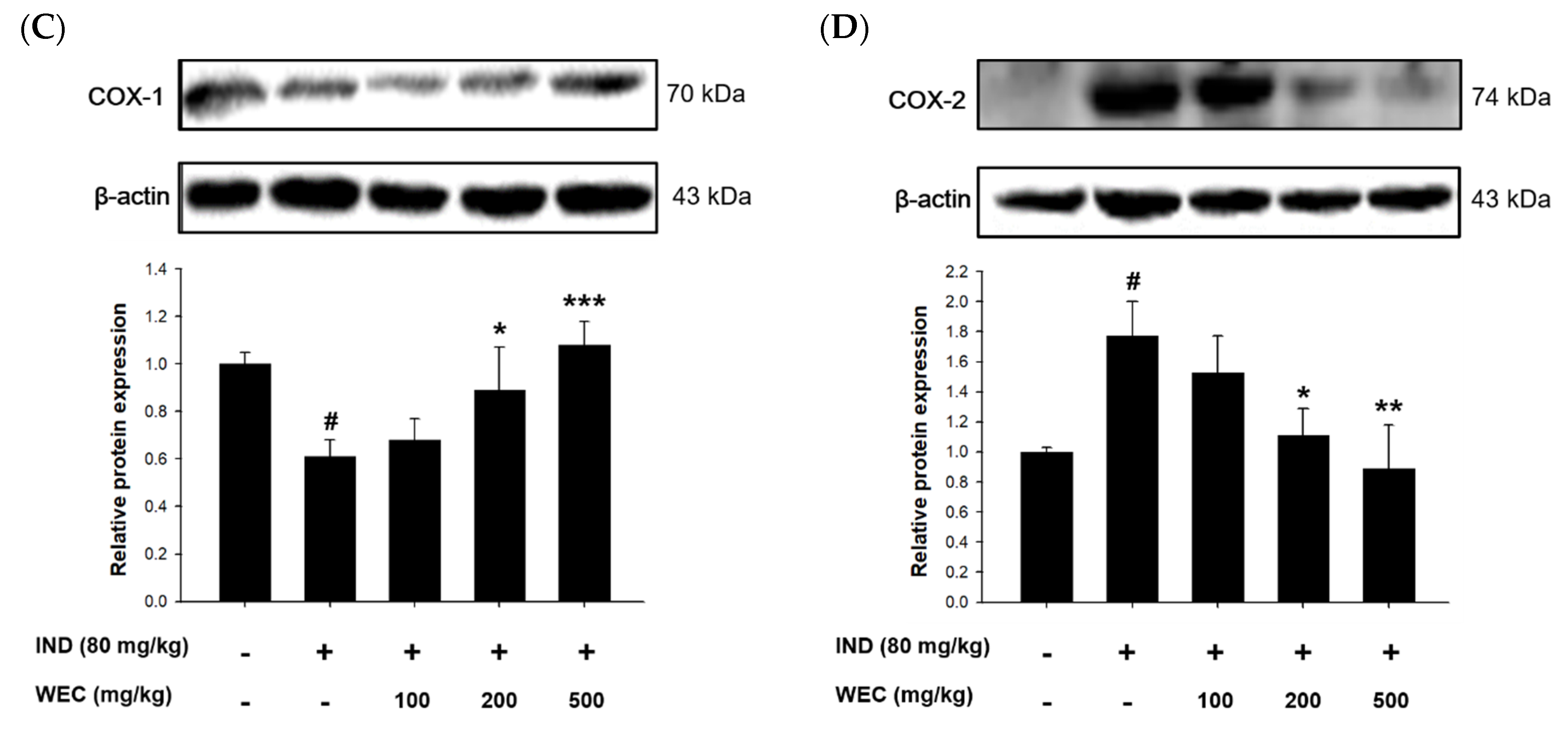
3.5. Effects of WEC on the Contents of GSH and PGE2 and Protein Levels of COX-1 and COX-2 of Gastric Mucosa in IND-Treated Wistar Rats
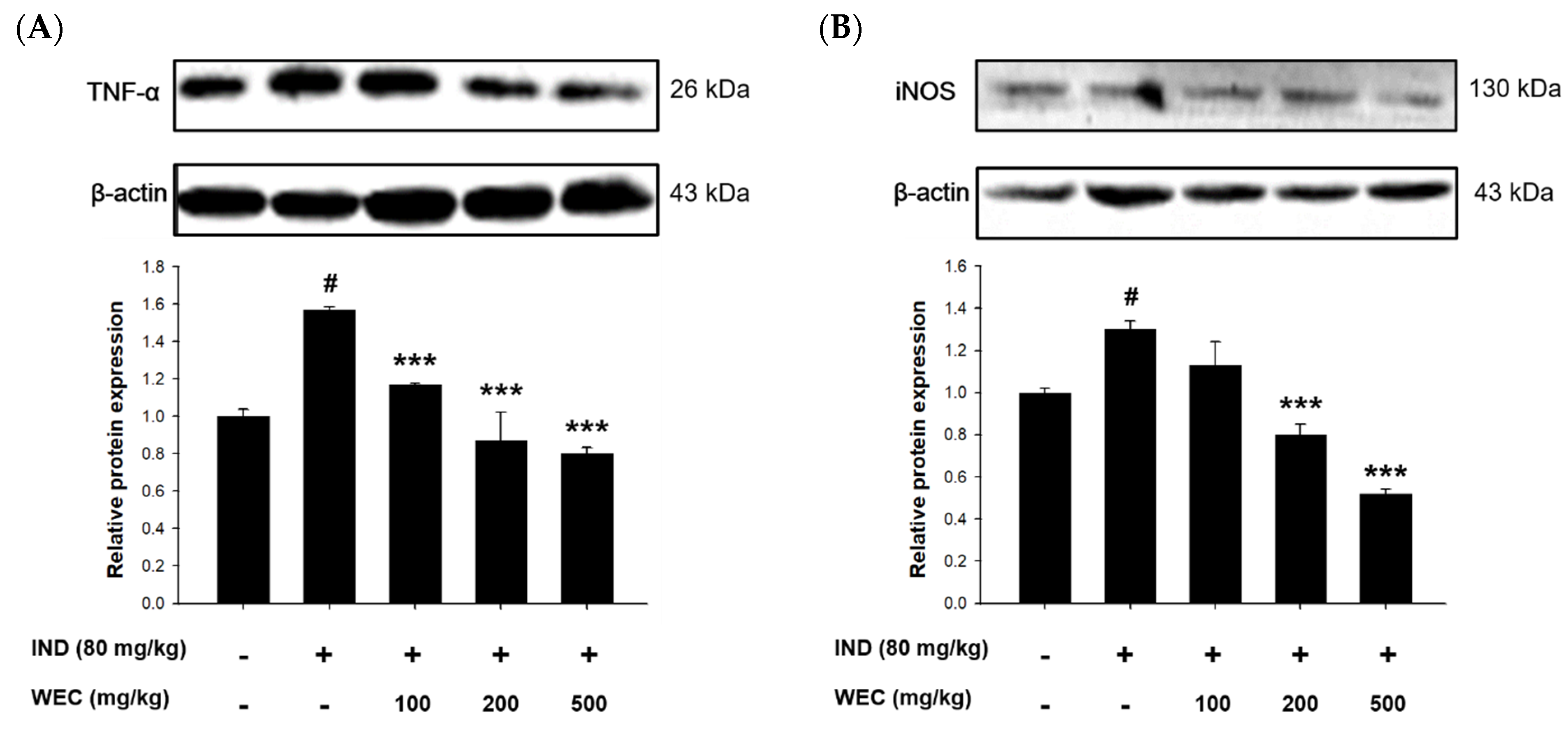
3.6. Effects of WEC on Protein Levels of TNF-α and iNOS of Gastric Mucosa in IND-Treated Wistar Rats
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuna, L.; Jakab, J.; Smolic, R.; Raguz-Lucic, N.; Vcev, A.; Smolic, M. Peptic ulcer disease: A brief review of conventional therapy and herbal treatment options. J. Clin. Med. 2019, 8, 179. [Google Scholar] [CrossRef] [Green Version]
- Lanas, A.; Chan, F.K.L. Peptic ulcer disease. Lancet 2017, 390, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Shiotani, A.; Yamaoka, Y.; El-Zimaity, H.M.; Saeed, M.A.; Qureshi, W.A.; Graham, D.Y. NSAID gastric ulceration: Predictive value of gastric pH, mucosal density of polymorphonuclear leukocytes, or levels of IL-8 or nitrite. Dig. Dis. Sci. 2002, 47, 38–43. [Google Scholar] [CrossRef]
- Hansson, L.E.; Nyren, O.; Hsing, A.W.; Bergstrom, R.; Josefsson, S.; Chow, W.H.; Fraumeni, J.F., Jr.; Adami, H.O. The risk of stomach cancer in patients with gastric or duodenal ulcer disease. N. Engl. J. Med. 1996, 335, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Barsouk, A. Epidemiology of gastric cancer: Global trends, risk factors and prevention. Prz. Gastroenterol. 2019, 14, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Badr, G.M.; Al-Mulhim, J.A. The protective effect of aged garlic extract on nonsteroidal anti-inflammatory drug-induced gastric inflammations in male albino rats. Evid. Based Complement. Alternat. Med. 2014, 2014, 759642. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.M.; Sakr, H.F. Effect of selenium and grape seed extract on indomethacin-induced gastric ulcers in rats. J. Physiol. Biochem. 2013, 69, 527–537. [Google Scholar] [CrossRef]
- Vemula, S.K.; Chawada, M.B.; Thakur, K.S.; Vahalia, M.K. Antiulcer activity of Amlapitta Mishran suspension in rats: A pilot study. Anc. Sci. Life 2012, 32, 112–115. [Google Scholar]
- Zhang, X.; Tajima, K.; Kageyama, K.; Kyoi, T. Irsogladine maleate suppresses indomethacin-induced elevation of proinflammatory cytokines and gastric injury in rats. World J. Gastroenterol. 2008, 14, 4784–4790. [Google Scholar] [CrossRef]
- Wallace, J.L. Prostaglandins, NSAIDs, and gastric mucosal protection: Why doesn’t the stomach digest itself? Physiol. Rev. 2008, 88, 1547–1565. [Google Scholar] [CrossRef]
- Chiou, S.K.; Hodges, A.; Hoa, N. Suppression of growth arrest and DNA damage-inducible 45α expression confers resistance to sulindac and indomethacin-induced gastric mucosal injury. J. Pharmacol. Exp. Ther. 2010, 334, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Birsen, I.; Izgut-Uysal, V.N. Protective effects of apelin on gastric mucosa. Tissue Cell 2022, 78, 101885. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, B.W.; Kwon, H.J.; Nam, S.W. Curative effect of selenium against indomethacin-induced gastric ulcers in rats. J. Microbiol. Biotechnol. 2011, 21, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Sigthorsson, G.; Crane, R.; Simon, T.; Hoover, M.; Quan, H.; Bolognese, J.; Bjarnason, I. COX-2 inhibition with rofecoxib does not increase intestinal permeability in healthy subjects: A double blind crossover study comparing rofecoxib with placebo and indomethacin. Gut 2000, 47, 527–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, P.S.; Tung, Y.T.; Lee, W.T.; Yen, G.C. Protective effect of camellia oil (Camellia oleifera Abel.) against ethanol-induced acute oxidative injury of the gastric mucosa in mice. J. Agric. Food Chem. 2017, 65, 4932–4941. [Google Scholar] [CrossRef] [PubMed]
- Musumba, C.; Pritchard, D.M.; Pirmohamed, M. Review article: Cellular and molecular mechanisms of NSAID-induced peptic ulcers. Aliment. Pharmacol. Ther. 2009, 30, 517–531. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [Green Version]
- Mazumder, S.; De, R.; Debsharma, S.; Bindu, S.; Maity, P.; Sarkar, S.; Saha, S.J.; Siddiqui, A.A.; Banerjee, C.; Nag, S.; et al. Indomethacin impairs mitochondrial dynamics by activating the PKCζ-p38-DRP1 pathway and inducing apoptosis in gastric cancer and normal mucosal cells. J. Biol. Chem. 2019, 294, 8238–8258. [Google Scholar] [CrossRef]
- Park, Y.M.; Won, J.H.; Yun, K.J.; Ryu, J.H.; Han, Y.N.; Choi, S.K.; Lee, K.T. Preventive effect of Ginkgo biloba extract (GBB) on the lipopolysaccharide-induced expressions of inducible nitric oxide synthase and cyclooxygenase-2 via suppression of nuclear factor-κB in raw 264.7 cells. Biol. Pharm. Bull. 2006, 29, 985–990. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Tsai, J.S.; Hung, L.B.; Pan, B.S. Hypocholesterolemic effect of compounded freshwater clam protein hydrolysate and gracilaria. Food Chem. 2010, 123, 395–399. [Google Scholar] [CrossRef]
- Chijimatsu, T.; Tatsuguchi, I.; Oda, H.; Mochizuki, S. A freshwater clam (Corbicula fluminea) extract reduces cholesterol level and hepatic lipids in normal rats and xenobiotics-induced hypercholesterolemic rats. J. Agric. Food Chem. 2009, 57, 3108–3112. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.L.; Hsu, W.H.; Tu, C.M.; Wang, W.H.; Yang, C.Y.; Lee, H.K.; Chin, T.Y. The effects of freshwater clam (Corbicula fluminea) extract on activated hepatic stellate cells. Evid. Based Complement. Alternat. Med. 2021, 2021, 6065168. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Hsu, C.C.; Yen, G.C. Hepatoprotection by freshwater clam extract against CCl4-induced hepatic damage in rats. Am. J. Chin. Med. 2010, 38, 881–894. [Google Scholar] [CrossRef] [PubMed]
- Liao, N.; Chen, S.; Ye, X.; Zhong, J.; Wu, N.; Dong, S.; Yang, B.; Liu, D. Antioxidant and anti-tumor activity of a polysaccharide from freshwater clam, Corbicula fluminea. Food Funct. 2013, 4, 539–548. [Google Scholar] [CrossRef]
- Huang, Y.T.; Huang, Y.H.; Hour, T.C.; Pan, B.S.; Liu, Y.C.; Pan, M.H. Apoptosis-inducing active components from Corbicula fluminea through activation of caspase-2 and production of reactive oxygen species in human leukemia HL-60 cells. Food Chem. Toxicol. 2006, 44, 1261–1272. [Google Scholar] [CrossRef]
- Chen, T.Y.; Lin, B.C.; Shiao, M.S.; Pan, B.S. Lipid-lowering and LDL-oxidation inhibitory effects of aqueous extract of freshwater clam (Corbicula fluminea)—Using tilapia as an animal model. J. Food Sci. 2008, 73, H148–H154. [Google Scholar] [CrossRef]
- Huang, S.C.; Lin, J.J.; Lee, M.F.; Liu, Y.C.; Pan, B.S. Freshwater clam extracts alleviate dyslipidaemia of tilapia fed a high-fat diet as an animal model. J. Funct. Foods 2016, 25, 559–567. [Google Scholar] [CrossRef]
- Furukawa, O.; Nakamura, E.; Okabe, S. Characterization of a novel cell damage model induced by acid and pepsin using rat gastric epithelial cells: Protective effect of sucralfate. J. Gastroenterol. Hepatol. 1997, 12, 115–121. [Google Scholar] [CrossRef]
- Robert, A.; Böttcher, W.; Golanska, E.; Kauffman, G.L., Jr. Lack of correlation between mucus gel thickness and gastric cytoprotection in rats. Gastroenterology 1984, 86, 670–674. [Google Scholar] [CrossRef]
- Beck, P.L.; Xavier, R.; Lu, N.; Nanda, N.N.; Dinauer, M.; Podolsky, D.K.; Seed, B. Mechanisms of NSAID-induced gastrointestinal injury defined using mutant mice. Gastroenterology 2000, 119, 699–705. [Google Scholar] [CrossRef]
- Tsai, H.Y.; Shih, Y.Y.; Yeh, Y.T.; Huang, C.H.; Liao, C.A.; Hu, C.Y.; Nagabhushanam, K.; Ho, C.T.; Chen, Y.K. Pterostilbene and its derivative 3′-hydroxypterostilbene ameliorated nonalcoholic fatty liver disease through synergistic modulation of the gut microbiota and sirt1/AMPK signaling pathway. J. Agric. Food Chem. 2022, 70, 4966–4980. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.H.; Liu, P.Y.; Lin, Y.L.; Tsai, J.S. Hypoglycemic peptide-enriched hydrolysates of Corbicula fluminea and Chlorella sorokiniana possess synergistic hypoglycemic activity through inhibiting α-glucosidase and dipeptidyl peptidase-4 activity. J. Sci. Food Agric. 2022, 102, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.H.; Vo, T.S.; Ngo, D.N.; Wijesekara, I.; Kim, S.K. Biological activities and potential health benefits of bioactive peptides derived from marine organisms. Int. J. Biol. Macromol. 2012, 51, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, C.; Qin, X.; Cao, W.; Chen, J.; Li, Y.; Zheng, H.; Lin, H.; Chen, Z. Hepatoprotective effect of clam (Corbicula fluminea) protein hydrolysate on alcohol-induced liver injury in mice and partial identification of a hepatoprotective peptide from the hydrolysate. Food Sci. Technol. 2022, 42, e61522. [Google Scholar] [CrossRef]
- Urushidani, T.; Okabe, S.; Takeuchi, K.; Takagi, K. Effects of various amino acids on indomethacin-induced gastric ulcers in rats. Jpn J. Pharmacol. 1977, 27, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Abu Bakar, N.; Hakim Abdullah, M.N.; Lim, V.; Yong, Y.K. Gastroprotective effect of polypeptide-k isolated from Momordica charantia’s seeds on multiple experimental gastric ulcer models in rats. Evid. Based Complement. Alternat. Med. 2022, 2022, 6098929. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D. 5-hydroxytryptamine (serotonin) in the gastrointestinal tract. Curr. Opin. Endocrinol. Diabetes Obes 2013, 20, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Ren, M.; Zhang, S.H.; Zeng, X.F.; Liu, H.; Qiao, S.Y. Branched-chain amino acids are beneficial to maintain growth performance and intestinal immune-related gunction in weaned piglets fed protein restricted diet. Asian-Australas J. Anim. Sci. 2015, 28, 1742–1750. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Zhao, Y.J.; Dong, H.Z.; Su, G.W.; Zhao, M.M. Structure-activity relationship of antioxidant dipeptides: Dominant role of Tyr, Trp, Cys and Met residues. J. Funct. Foods 2016, 21, 485–496. [Google Scholar] [CrossRef]
- Nagarjun, S.; Dhadde, S.B.; Veerapur, V.P.; Thippeswamy, B.S.; Chandakavathe, B.N. Ameliorative effect of chromium-D-phenylalanine complex on indomethacin-induced inflammatory bowel disease in rats. Biomed. Pharmacother. 2017, 89, 1061–1066. [Google Scholar] [CrossRef]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Adhikary, B.; Yadav, S.K.; Roy, K.; Bandyopadhyay, S.K.; Chattopadhyay, S. Black tea and theaflavins assist healing of indomethacin-induced gastric ulceration in mice by antioxidative action. Evid. Based Complement. Alternat. Med. 2011, 2011, 546560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, K. Pathogenesis of NSAID-induced gastric damage: Importance of cyclooxygenase inhibition and gastric hypermotility. World J. Gastroenterol. 2012, 18, 2147–2160. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.; Matsui, H.; Kaneko, T.; Nagano, Y.; Shimokawa, O.; Udo, J.; Hirayama, A.; Hyodo, I.; Indo, H.P.; Majima, H.J. Lansoprazole inhibits mitochondrial superoxide production and cellular lipid peroxidation induced by indomethacin in RGM1 cells. J. Clin. Biochem. Nutr. 2011, 49, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.T.; Lee, J.Y.; Lee, J.; Kim, H.; Yoon, K.S.; Choe, W.; Kang, I. Oleic acid reduces lipopolysaccharide-induced expression of iNOS and COX-2 in BV2 murine microglial cells: Possible involvement of reactive oxygen species, p38 MAPK, and IKK/NF-κB signaling pathways. Neurosci. Lett. 2009, 464, 93–97. [Google Scholar] [CrossRef]
- He, F.; Wu, C.; Li, P.; Li, N.; Zhang, D.; Zhu, Q.; Ren, W.; Peng, Y. Functions and signaling pathways of amino acids in intestinal inflammation. Biomed. Res. Int. 2018, 2018, 9171905. [Google Scholar] [CrossRef]
- Kwon, D.A.; Kim, Y.S.; Baek, S.H.; Kim, S.K.; Kim, H.K.; Jo, S.K.; Jung, U.; Park, H.R.; Lee, H.S. Protective effects of a standardized extract (HemoHIM) using indomethacin- and ethanol/HCl-induced gastric mucosal injury models. Pharm. Biol. 2019, 57, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Athaydes, B.R.; Alves, G.M.; Assis, A.; Gomes, J.V.D.; Rodrigues, R.P.; Campagnaro, B.P.; Nogueira, B.V.; Silveira, D.; Kuster, R.M.; Pereira, T.M.C.; et al. Avocado seeds (Persea americana Mill.) prevents indomethacin-induced gastric ulcer in mice. Food Res. Int. 2019, 119, 751–760. [Google Scholar] [CrossRef]
- Zeng, Y.; Guan, Y.; Han, W.; Sun, Y. Antioxidant peptides from freshwater clam extract using enzymatic hydrolysis. Afr. J. Food Sci. 2014, 8, 148–154. [Google Scholar]
- Chi, C.-F.; Hu, F.-Y.; Wang, B.; Li, T.; Ding, G.-F. Antioxidant and anticancer peptides from the protein hydrolysate of blood clam (Tegillarca granosa) muscle. J. Funct. Foods 2015, 15, 301–313. [Google Scholar] [CrossRef]
- Othman, A.I.; El-Missiry, M.A.; Amer, M.A. The protective action of melatonin on indomethacin-induced gastric and testicular oxidative stress in rats. Redox Rep. 2001, 6, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Beatty, P.; Reed, D.J. Influence of cysteine upon the glutathione status of isolated rat hepatocytes. Biochem. Pharmacol. 1981, 30, 1227–1230. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.M.; Chou, S.T.; Lin, S.C.; Su, Z.Y.; Sheen, L.Y. Protective effects of water extract of clam on normal and CCl4-induced damage in primary cultured rat hepatocytes. Am. J. Chin. Med. 2010, 38, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.H.; Zhang, Z.B.; Zheng, Y.F.; Chen, H.M.; Yu, X.T.; Chen, X.Y.; Zhang, X.; Xie, J.H.; Su, Z.Q.; Feng, X.X.; et al. Gastroprotective effect of andrographolide sodium bisulfite against indomethacin-induced gastric ulceration in rats. Int. Immunopharmacol. 2015, 26, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Peskar, B.M. Role of cyclooxygenase isoforms in gastric mucosal defence. J. Physiol. Paris 2001, 95, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.H.; Ke, C.H.; Chen, C.C.; Chuang, C.H.; Liao, K.W.; Shiao, Y.H.; Lin, C.S. The effects of freshwater clam (Corbicula fluminea) extract on serum tumor necrosis factor-α (TNF-α) in prediabetic patients in Taiwan. Mar. Drugs 2022, 20, 261. [Google Scholar] [CrossRef]
- Martin-Venegas, R.; Brufau, M.T.; Guerrero-Zamora, A.M.; Mercier, Y.; Geraert, P.A.; Ferrer, R. The methionine precursor DL-2-hydroxy-(4-methylthio)butanoic acid protects intestinal epithelial barrier function. Food Chem. 2013, 141, 1702–1709. [Google Scholar] [CrossRef]
- Mine, Y.; Zhang, H. Anti-inflammatory effects of poly-l-lysine in intestinal mucosal system mediated by calcium-sensing receptor activation. J. Agric. Food Chem. 2015, 63, 10437–10447. [Google Scholar] [CrossRef]
- Ju, Z.; Li, M.; Xu, J.; Howell, D.C.; Li, Z.; Chen, F.E. Recent development on COX-2 inhibitors as promising anti-inflammatory agents: The past 10 years. Acta Pharm. Sin. B 2022, 12, 2790–2807. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Free Amino Acid (mg/g) | Hydrolysate (mg/g) | |
|---|---|---|
| Aspartic acid | 0.41 ± 0.03 | 66 ± 4 |
| Glutamic acid | 0.59 ± 0.05 | 93 ± 9 |
| Serine | 0.85 ± 0.07 | 30 ± 2 |
| Histidine | 0.30 ± 0.02 | 9 ± 1 |
| Glycine | 0.27 ± 0.03 | 24 ± 2 |
| Threonine | 1.64 ± 0.07 | 26 ± 2 |
| Arginine | 2.1 ± 0.1 | 44 ± 3 |
| Alanine | 0.99 ± 0.08 | 30 ± 1 |
| Tyrosine | 0.97 ± 0.06 | 19 ± 1 |
| Cysteine | n.d. | 33 ± 3 |
| Valine | 3.0 ± 0.2 | 30 ± 1 |
| Methionine | 1.35 ± 0.08 | 10 ± 1 |
| Phenylalanine | 2.4 ± 0.2 | 30 ± 2 |
| Isoleucine | 3.4 ± 0.3 | 34 ± 2 |
| Leucine | 6.8 ± 0.5 | 50 ± 5 |
| Lysine | 2.13 ± 0.09 | 56 ± 4 |
| Proline | 0.25 ± 0.01 | 25 ± 1 |
| Tryptophan | 0.62 ± 0.03 | n.d. |
| Asparagine | 0.65 ± 0.04 | n.d. |
| Taurine | 0.24 ± 0.02 | n.d. |
| Group | Ulcer Area (mm2) | Preventive Index (%) |
|---|---|---|
| Control | 0 ± 0 | ─ |
| IND | 75 ± 26 # | ─ |
| WEC (100 mg/kg) | 47 ± 14 * | 38 |
| WEC (200 mg/kg) | 37 ± 9 ** | 51 |
| WEC (500 mg/kg) | 20 ± 6 *** | 74 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isnain, F.S.; Liao, N.-C.; Tsai, H.-Y.; Zhao, Y.-J.; Huang, C.-H.; Hsu, J.-L.; Wardani, A.K.; Chen, Y.-K. Freshwater Clam Extract Attenuates Indomethacin-Induced Gastric Damage In Vitro and In Vivo. Foods 2023, 12, 156. https://doi.org/10.3390/foods12010156
Isnain FS, Liao N-C, Tsai H-Y, Zhao Y-J, Huang C-H, Hsu J-L, Wardani AK, Chen Y-K. Freshwater Clam Extract Attenuates Indomethacin-Induced Gastric Damage In Vitro and In Vivo. Foods. 2023; 12(1):156. https://doi.org/10.3390/foods12010156
Chicago/Turabian StyleIsnain, Fuad Sauqi, Nai-Chen Liao, Hui-Yun Tsai, Yu-Jie Zhao, Chien-Hua Huang, Jue-Liang Hsu, Agustin Krisna Wardani, and Yu-Kuo Chen. 2023. "Freshwater Clam Extract Attenuates Indomethacin-Induced Gastric Damage In Vitro and In Vivo" Foods 12, no. 1: 156. https://doi.org/10.3390/foods12010156
APA StyleIsnain, F. S., Liao, N.-C., Tsai, H.-Y., Zhao, Y.-J., Huang, C.-H., Hsu, J.-L., Wardani, A. K., & Chen, Y.-K. (2023). Freshwater Clam Extract Attenuates Indomethacin-Induced Gastric Damage In Vitro and In Vivo. Foods, 12(1), 156. https://doi.org/10.3390/foods12010156