
Environmental Factors Affecting the Diversity and Composition of Environmental Microorganisms in the Shaoxing Rice Wine Producing Area

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. High-Throughput Sequencing Analysis
2.2.1. Microbial Enrichment in Pretreatment Samples
2.2.2. DNA Extraction and PCR Amplification
2.2.3. Data Processing and Analysis
2.3. Environmental Factors
2.4. Statistical Analysis
3. Results and Discussions
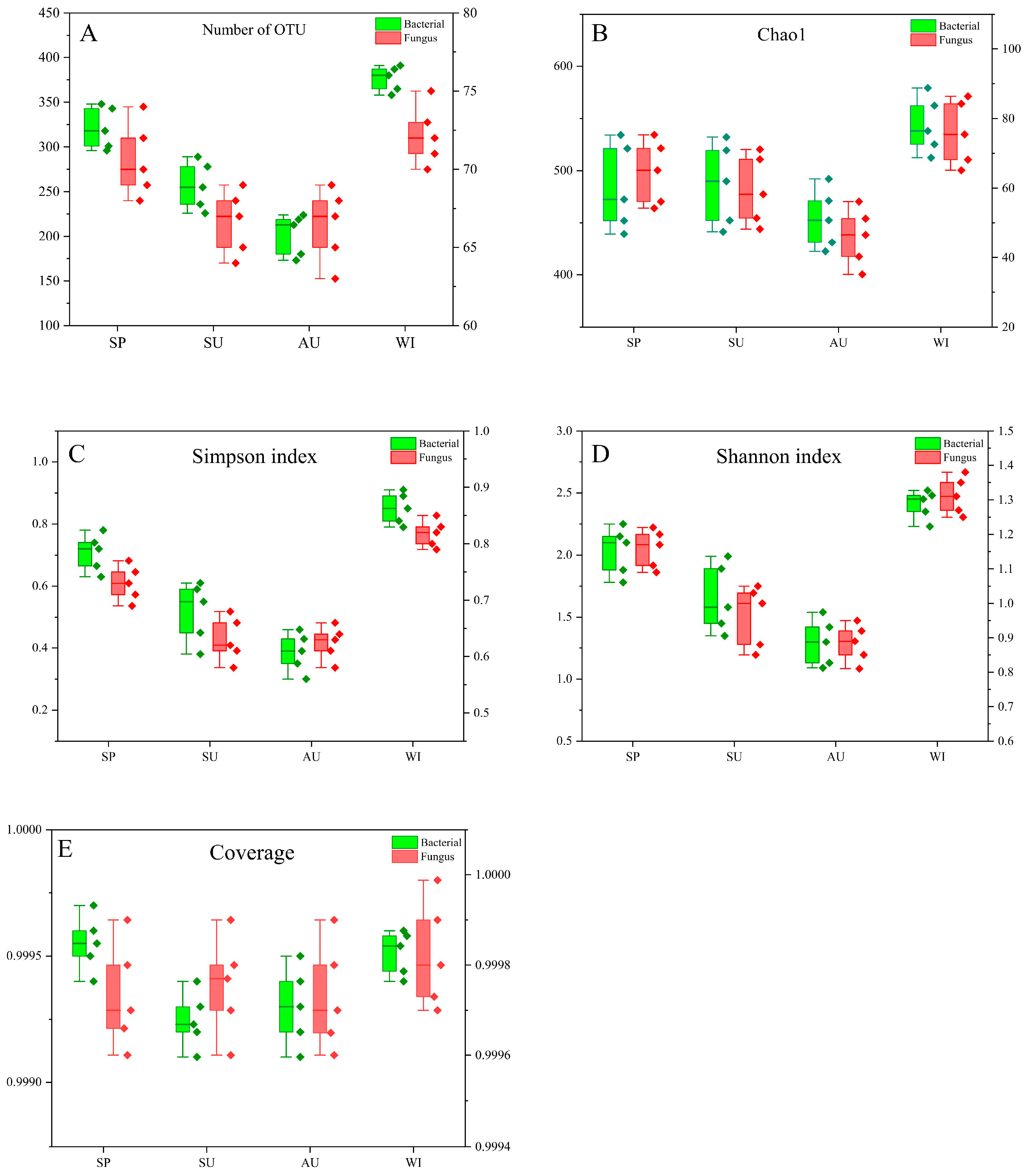
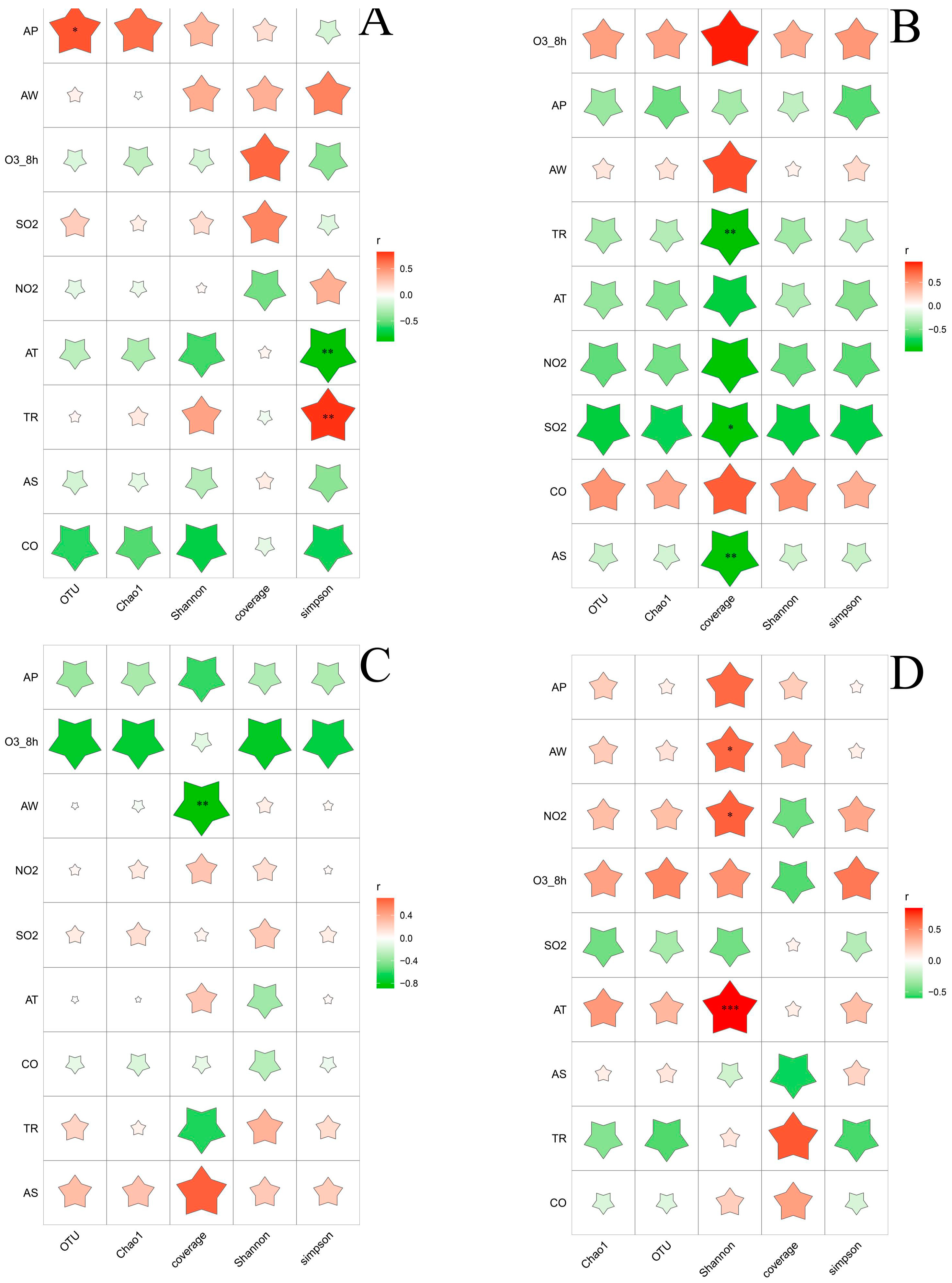
3.1. Correlation between Environmental Factors and Alpha Diversity of Bacterial and Fungal Communities
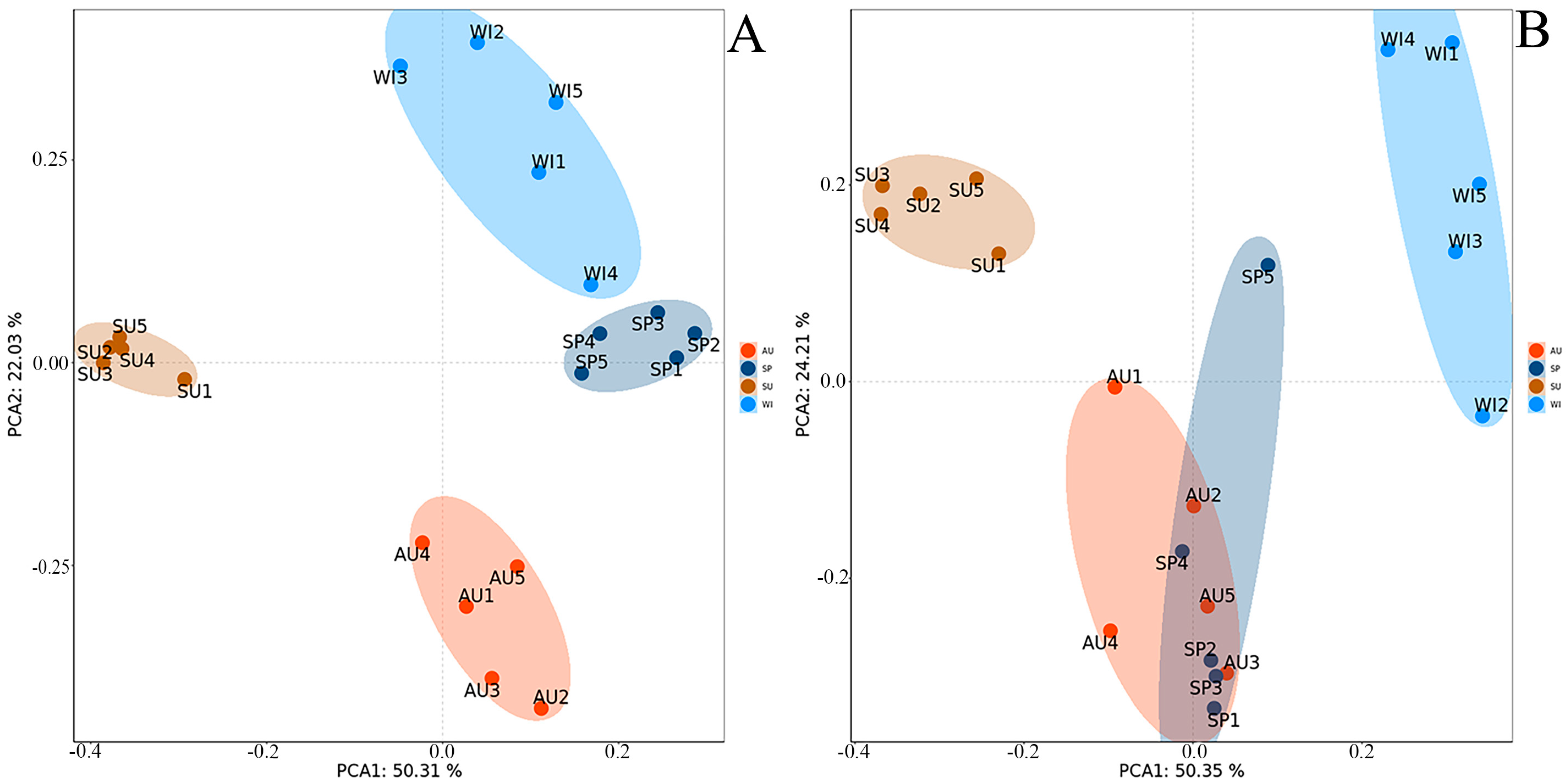
3.2. Similarity Analysis of Bacterial and Fungal Community Structure
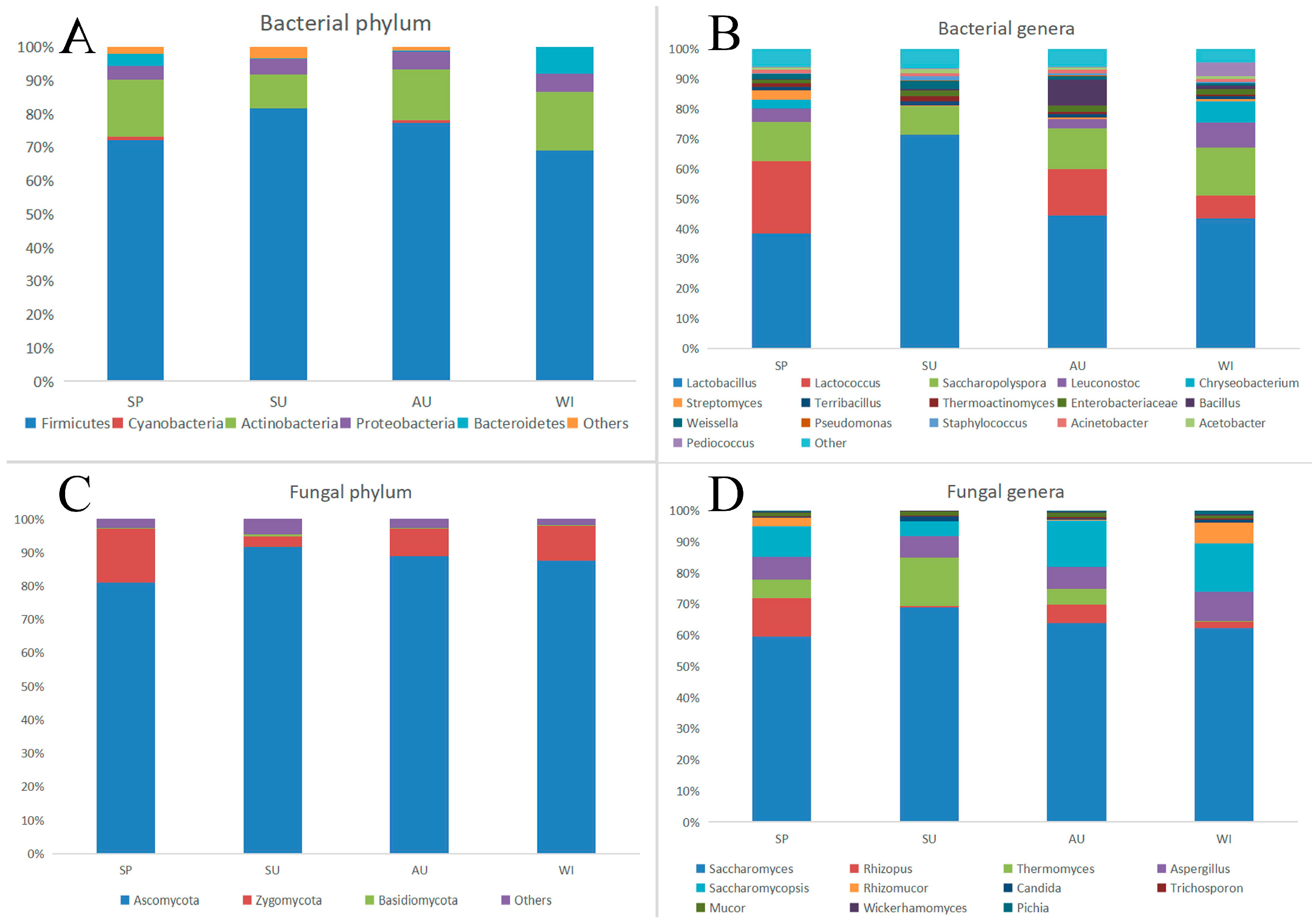
3.3. Structural Changes of Microbial Colonies in Environmental Samples
3.3.1. Bacterial Community Structure
3.3.2. Comparison of Fungal Community Structure
3.4. Relationship between Environmental Factors and Dominant Microbial Communities
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cao, Y.; Xie, G.F.; Wu, C.; Lu, J.A. A Study on Characteristic Flavor Compounds in Traditional Chinese Rice Wine—Guyue Longshan Rice Wine. J. Inst. Brew. 2010, 116, 182–189. [Google Scholar] [CrossRef]
- Chen, C.; Liu, Y.; Tian, H.X.; Ai, L.Z.; Yu, H.Y. Metagenomic analysis reveals the impact of JIUYAO microbial diversity on fermentation and the volatile profile of Shaoxing-jiu. Food Microbiol. 2020, 86, 103326. [Google Scholar] [CrossRef] [PubMed]
- France, M.T.; Remold, S.K. Interference Competition Among Household Strains of Pseudomonas. Microb. Ecol. 2016, 72, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.P.; Hu, J.; Xu, Y.Z.; Xue, J.B.; Zhou, J.D.; Han, X.; Ji, Z.W.; Mao, J. Combined use of single molecule real-time DNA sequencing technology and culture-dependent methods to analyze the functional microorganisms in inoculated raw wheat Qu. Food Res. Int. 2020, 132, 109062. [Google Scholar] [CrossRef]
- Zhang, K.; Li, Q.; Wu, W.; Yang, J.; Zou, W. Wheat Qu and Its Production Technology, Microbiota, Flavor, and Metabolites. J. Food Sci. 2019, 84, 2373–2386. [Google Scholar] [CrossRef]
- Lv, X.G.; Jia, R.B.; Li, Y.; Chen, F.; Chen, Z.C.; Liu, B.; Chen, S.J.; Rao, P.F.; Ni, L. Characterization of the dominant bacterial communities of traditional fermentation starters for Hong Qu glutinous rice wine by means of MALDI-TOF mass spectrometry fingerprinting, 16S rRNA gene sequencing and species-specific PCRs. Food Control 2016, 67, 292–302. [Google Scholar] [CrossRef]
- Huang, Z.R.; Hong, J.L.; Xu, J.X.; Li, L.; Guo, W.L.; Pan, Y.Y.; Chen, S.J.; Bai, W.D.; Rao, P.F.; Ni, L.; et al. Exploring core functional microbiota responsible for the production of volatile flavour during the traditional brewing of Wuyi Hong Qu glutinous rice wine. Food Microbiol. 2018, 76, 487–496. [Google Scholar] [CrossRef]
- Huang, Z.; Shen, Y.; Huang, X.; Qiao, M.; He, R.K.; Song, L. Microbial diversity of representative traditional fermented sausages in different regions of China. J. Appl. Microbiol. 2021, 130, 133–141. [Google Scholar] [CrossRef]
- Du, H.; Wang, X.; Zhang, Y.; Xu, Y. Exploring the impacts of raw materials and environments on the microbiota in Chinese Daqu starter. Int. J. Food Microbiol. 2019, 297, 32–40. [Google Scholar] [CrossRef]
- Zhu, Y.; Cao, Y.Y.; Yang, M.; We, P.C.; Cao, L.; Ma, J.; Zhang, Z.M.; Zhang, W.B. Bacterial diversity and community in Qula from the Qinghai-Tibetan Plateau in China. Peerj 2018, 6, e6044. [Google Scholar] [CrossRef]
- Peng, Q.; Chen, X.; Zheng, H.; Meng, K.; Wu, J.; Xie, G.; Zhang, L.; Feng, X.; Li, L.; Fang, S.; et al. Spatial and temporal distribution of environmental microbiota in Chinese rice wine (Huangjiu) natural fermentation wineries. Food Biosci. 2023, 55, 102929. [Google Scholar] [CrossRef]
- Zhao, X.X.; Xiang, F.S.; Tang, F.X.; Cai, W.C.; Guo, Z.; Hou, Q.C.; Yang, X.Q.; Song, W.; Shan, C.H. Bacterial Communities and Prediction of Microbial Metabolic Pathway in Rice Wine Koji from Different Regions in China. Front. Microbiol. 2022, 12, 748779. [Google Scholar] [CrossRef]
- Liu, X.Y.; Shi, Y.; Kong, L.Y.; Tong, L.H.; Cao, H.X.; Zhou, H.; Lv, Y.Z. Long-Term Application of Bio-Compost Increased Soil Microbial Community Diversity and Altered Its Composition and Network. Microorganisms 2022, 10, 462. [Google Scholar] [CrossRef] [PubMed]
- Valverde, J.R.; Gullón, S.; Pérez Mellado, R.J.P.o. Looking for rhizobacterial ecological indicators in agricultural soils using 16S rRNA metagenomic amplicon data. PLoS ONE 2016, 11, e0165204. [Google Scholar] [CrossRef] [PubMed]
- Malas, B.; Mohamad, M.; Yazji, S. Nisin production conditions optimization and its effect on Bacillus cereus and Listeria monocytogenes. Int. Food Res. J. 2017, 24, 900–903. [Google Scholar]
- Zheng, X.W.; Yan, Z.; Han, B.Z.; Zwietering, M.H.; Samson, R.A.; Boekhout, T.; Nout, M.J.R. Complex microbiota of a Chinese “Fen” liquor fermentation starter (Fen-Daqu), revealed by culture-dependent and culture-independent methods. Food Microbiol. 2012, 31, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Costello, P.J.; Lee, T.H.; Henschke, P. Ability of lactic acid bacteria to produce N-heterocycles causing mousy off-flavour in wine. Aust. J. Grape Wine Res. 2001, 7, 160–167. [Google Scholar] [CrossRef]
- Xia, Y.; Luo, H.B.; Wu, Z.Y.; Zhang, W.X. Microbial diversity in jiuqu and its fermentation features: Saccharification, alcohol fermentation and flavors generation. Appl. Microbiol. Biotechnol. 2023, 107, 25–41. [Google Scholar] [CrossRef]
- Gao, L.; Zhou, J.; He, G.Q. Effect of microbial interaction on flavor quality in Chinese baijiu fermentation. Front. Nutr. 2022, 9, 960712. [Google Scholar] [CrossRef]
- Hu, Y.L.; Lei, X.Y.; Zhang, X.M.; Guan, T.W.; Wang, L.Y.; Zhang, Z.J.; Yu, X.; Tu, J.M.; Peng, N.; Liang, Y.X.; et al. Characteristics of the Microbial Community in the Production of Chinese Rice-Flavor Baijiu and Comparisons With the Microflora of Other Flavors of Baijiu. Front. Microbiol. 2021, 12, 673670. [Google Scholar] [CrossRef]
- Kameda, T.; Aoki, H.; Yanaka, N.; Kumrungsee, T.; Kato, N. Production of Isoflavone Aglycone-enriched Tempeh with Rhizopus stolonifer. Food Sci. Technol. Res. 2018, 24, 493–499. [Google Scholar] [CrossRef]
- Liu, D.F.; Zhang, H.T.; Lin, C.C.; Xu, B.G. Optimization of rice wine fermentation process based on the simultaneous saccharification and fermentation kinetic model. Chin. J. Chem. Eng. 2016, 24, 1406–1412. [Google Scholar] [CrossRef]
- Yu, H.Y.; Guo, W.; Xie, T.; Ai, L.Z.; Tian, H.X.; Chen, C. Aroma characteristics of traditional Huangjiu produced around Winter Solstice revealed by sensory evaluation, gas chromatography-mass spectrometry and gas chromatography-ion mobility spectrometry. Food Res. Int. 2021, 145, 110421. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Y.; Li, Q.W.; Guo, W.; Chen, C.; Ai, L.Z.; Tian, H.X. Dynamic analysis of volatile metabolites and microbial community and their correlations during the fermentation process of traditional Huangjiu (Chinese rice wine) produced around Winter Solstice. Food Chem. X 2023, 18, 100620. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, C.Y.; Xiang, X.Q.; Xu, H.L.; Han, G.Q. Analysis of microbial diversity and succession during Xiaoqu Baijiu fermentation using high-throughput sequencing technology. Eng. Life Sci. 2022, 22, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Xia, Q.L.; Chen, J.B.; Jin, Z.; Aniya. Understanding of microbial diversity in three representative Qu in China and characterization of the volatile compounds in the corresponding Chinese rice wine. Lwt-Food Sci. Technol. 2022, 164, 113680. [Google Scholar] [CrossRef]
- Lv, X.C.; Weng, X.; Zhang, W.; Rao, P.F.; Ni, L. Microbial diversity of traditional fermentation starters for Hong Qu glutinous rice wine as determined by PCR-mediated DGGE. Food Control 2012, 28, 426–434. [Google Scholar] [CrossRef]
- Liu, S.P.; Zhou, Y.; Ma, D.N.; Zhang, S.Y.; Dong, Y.; Zhang, X.; Mao, J. Environment microorganism and mature daqu powder shaped microbial community formation in mechanically strong-flavor daqu. Food Biosci. 2023, 52, 102467. [Google Scholar] [CrossRef]
- Liu, D.F.; Zhang, H.T.; Xiong, W.L.; Hu, J.H.; Xu, B.G.; Lin, C.C.; Xu, L.; Jiang, L.H. Effect of Temperature on Chinese Rice Wine Brewing with High Concentration Presteamed Whole Sticky Rice. Biomed Res. Int. 2014, 2014, 426929. [Google Scholar] [CrossRef]
- Wu, X.Y.; Jing, R.X.; Chen, W.H.; Geng, X.J.; Li, M.; Yang, F.Z.; Yan, Y.Z.; Liu, Y. High-throughput sequencing of the microbial diversity of roasted-sesame-like flavored Daqu with different characteristics. 3 Biotech 2020, 10, 502. [Google Scholar] [CrossRef]
- Scrimgeour, N.; Nordestgaard, S.; Lloyd, N.D.R.; Wilkes, E.N. Exploring the effect of elevated storage temperature on wine composition. Aust. J. Grape Wine Res. 2015, 21, 713–722. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Q.; Zhang, L.; Huang, X.; Wu, J.; Cheng, Y.; Xie, G.; Feng, X.; Chen, X. Environmental Factors Affecting the Diversity and Composition of Environmental Microorganisms in the Shaoxing Rice Wine Producing Area. Foods 2023, 12, 3564. https://doi.org/10.3390/foods12193564
Peng Q, Zhang L, Huang X, Wu J, Cheng Y, Xie G, Feng X, Chen X. Environmental Factors Affecting the Diversity and Composition of Environmental Microorganisms in the Shaoxing Rice Wine Producing Area. Foods. 2023; 12(19):3564. https://doi.org/10.3390/foods12193564
Chicago/Turabian StylePeng, Qi, Lili Zhang, Xiaoli Huang, Jianjiang Wu, Yujun Cheng, Guangfa Xie, Xinxin Feng, and Xueping Chen. 2023. "Environmental Factors Affecting the Diversity and Composition of Environmental Microorganisms in the Shaoxing Rice Wine Producing Area" Foods 12, no. 19: 3564. https://doi.org/10.3390/foods12193564
APA StylePeng, Q., Zhang, L., Huang, X., Wu, J., Cheng, Y., Xie, G., Feng, X., & Chen, X. (2023). Environmental Factors Affecting the Diversity and Composition of Environmental Microorganisms in the Shaoxing Rice Wine Producing Area. Foods, 12(19), 3564. https://doi.org/10.3390/foods12193564