
Physicochemical Characteristics, Techno-Functionalities, and Amino Acid Profile of Prionoplus reticularis (Huhu) Larvae and Pupae Protein Extracts

 , , and
, , and
Abstract
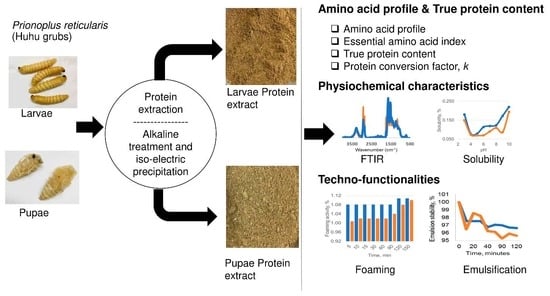
:
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Huhu Grub Protein Extraction Yield
2.3. Amino Acid Profiles of Huhu Grub Protein Extracts
2.4. Determination of the Essential Amino Acid Index (EAAI) and Nitrogen-Protein Conversion Factor
2.5. Determination of the True Protein and Nitrogen-to-Protein Conversion Factors for Huhu Protein Extracts
2.6. Colour Measurement of Huhu Grub Protein Extracts
2.7. Particle Size Analysis of the Huhu Grub Protein Extract Suspensions
2.8. Surface Hydrophobicity of the Huhu Grub Protein Extract Suspensions
2.9. Differential Scanning Calorimetry (DSC) of the Huhu Grub Protein Extracts
2.10. Fourier-Transform Infrared Spectroscopy (FTIR) of the Huhu Grub Protein Extracts
2.11. Foaming Capacity and Stability of the Huhu Grub Protein Extract Suspension
2.12. Water- and Oil-Holding Capacity of the Huhu Grub Protein Extract Suspensions
2.13. Emulsifying Activity/Capacity of the Huhu Grub Protein Extract Suspensions
2.14. Solubility Profile of the Huhu Grub Protein Extract Suspensions
2.15. Coagulation of the Huhu Grub Protein Extracts
2.16. Statistical Analysis
3. Results and Discussion
3.1. Huhu Grub Protein Extract Yield and Protein Recovery
3.2. Amino Acid Composition
3.3. Essential Amino Acid Index (EAAI) of Huhu Grub Protein Extracts
3.4. Crude Protein, True Protein, and Nitrogen-to-Protein Conversion Factors
3.5. Characterisation and Techno-Functionalities of Huhu Grub Protein Extracts
3.5.1. Colour Measurement
3.5.2. Surface Hydrophobicity of Huhu Grub Protein Extracts
3.5.3. Particle Size Distribution of the Extracted Protein Extract
3.5.4. Structural Characterisation of the Huhu Grub Protein Extracts by FTIR and DSC
3.6. Functionalities of Huhu Grub Extracted Proteins
3.6.1. Protein Solubility Profile
3.6.2. Foaming Capacity and Stability
3.6.3. Water- and Oil-Holding Capacity
3.6.4. Emulsifying Capacity and Stability
3.6.5. Coagulation of the Protein Concentrate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Huis, A. Potential of Insects as Food and Feed in Assuring Food Security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-W.; Setyabrata, D.; Lee, Y.J.; Jones, O.G.; Kim, Y.H.B. Pre-treated mealworm larvae and silkworm pupae as a novel protein ingredient in emulsion sausages. Innov. Food Sci. Emerg. Technol. 2016, 38, 116–123. [Google Scholar] [CrossRef]
- Bußler, S.; Rumpold, B.A.; Jander, E.; Rawel, H.M.; Schlüter, O.K. Recovery and techno-functionality of flours and proteins from two edible insect species: Meal worm (Tenebrio molitor) and black soldier fly (Hermetia illucens) larvae. Heliyon 2016, 2, e00218. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Lakemond, C.M.M.; Sagis, L.M.C.; Eisner-Schadler, V.; van Huis, A.; van Boekel, M.A.J.S. Extraction and characterisation of protein fractions from five insect species. Food Chem. 2013, 141, 3341–3348. [Google Scholar] [CrossRef]
- Zhao, X.; Vázquez-Gutiérrez, J.L.; Johansson, D.P.; Landberg, R.; Langton, M. Yellow Mealworm Protein for Food Purposes—Extraction and Functional Properties. PLoS ONE 2016, 11, e0147791. [Google Scholar] [CrossRef] [Green Version]
- Chaijan, M.; Chumthong, K.; Kongchoosi, N.; Chinarak, K.; Panya, A.; Phonsatta, N.; Cheong, L.; Panpipat, W. Characterisation of pH-shift-produced protein isolates from sago palm weevil (Rhynchophorus ferrugineus) larvae. J. Insects Food Feed 2022, 8, 313–324. [Google Scholar] [CrossRef]
- Clarkson, C.; Mirosa, M.; Birch, J. Potential of Extracted Locusta Migratoria Protein Fractions as Value-Added Ingredients. Insects 2018, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Mishyna, M.; Martinez, J.-J.I.; Chen, J.; Benjamin, O. Extraction, characterization and functional properties of soluble proteins from edible grasshopper (Schistocerca gregaria) and honey bee (Apis mellifera). Food Res. Int. 2019, 116, 697–706. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Kavle, R.R.; Carne, A.; Bekhit, A.E.-D.A.; Kebede, B.; Agyei, D. Macronutrients and mineral composition of wild harvested Prionoplus reticularis edible insect at various development stages: Nutritional and mineral safety implications. Int. J. Food Sci. Technol. 2022, 57, 6270–6278. [Google Scholar] [CrossRef]
- Kavle, R.R.; Carne, A.; Bekhit, A.E.-D.A.; Kebede, B.; Agyei, D. Proximate composition and lipid nutritional indices of larvae and pupae of the edible Huhu beetle (Prionoplus reticularis) endemic to New Zealand. J. Food Compos. Anal. 2022, 110, 104578. [Google Scholar] [CrossRef]
- Jayawardena, S.R.; Morton, J.D.; Brennan, C.S.; Bekhit, A.E.-D.A. Utilisation of beef lung protein powder as a functional ingredient to enhance protein and iron content of fresh pasta. Int. J. Food Sci. Technol. 2019, 54, 610–618. [Google Scholar] [CrossRef]
- Yang, F.; Huang, X.; Zhang, C.; Zhang, M.; Huang, C.; Yang, H. Amino acid composition and nutritional value evaluation of Chinese chestnut (Castanea mollissima Blume) and its protein subunit. RSC Adv. 2018, 8, 2653–2659. [Google Scholar] [CrossRef] [Green Version]
- WHO. Energy and protein requirements: Report of a joint FAO/WHO/UNU expert consultation. In Energy and Protein Requirements: Report of a Joint FAO/WHO/UNU Expert Consultation; WHO: Geneva, Switzerland, 1985; p. 206. [Google Scholar]
- Yu, X.; He, Q.; Wang, D. Dynamic Analysis of Major Components in the Different Developmental Stages of Tenebrio molitor. Front. Nutr. 2021, 8, 689746. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Cross, H.R.; Gehring, K.B.; Savell, J.W.; Arnold, A.N.; McNeill, S.H. Composition of free and peptide-bound amino acids in beef chuck, loin, and round cuts1,2. J. Anim. Sci. 2016, 94, 2603–2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafii, M.; Pencharz, P.B.; O Ball, R.; Tomlinson, C.; Elango, R.; Courtney-Martin, G. Bioavailable Methionine Assessed Using the Indicator Amino Acid Oxidation Method Is Greater When Cooked Chickpeas and Steamed Rice Are Combined in Healthy Young Men. J. Nutr. 2020, 150, 1834–1844. [Google Scholar] [CrossRef]
- Boulos, S.; Tännler, A.; Nyström, L. Nitrogen-to-Protein Conversion Factors for Edible Insects on the Swiss Market: T. molitor, A. domesticus, and L. migratoria. Front. Nutr. 2020, 7, 89. [Google Scholar] [CrossRef]
- Queiroz, L.S.; Regnard, M.; Jessen, F.; Mohammadifar, M.A.; Sloth, J.J.; Petersen, H.O.; Ajalloueian, F.; Brouzes, C.M.C.; Fraihi, W.; Fallquist, H.; et al. Physico-chemical and colloidal properties of protein extracted from black soldier fly (Hermetia illucens) larvae. Int. J. Biol. Macromol. 2021, 186, 714–723. [Google Scholar] [CrossRef]
- Fryer, H.J.; Davis, G.E.; Manthorpe, M.; Varon, S. Lowry protein assay using an automatic microtiter plate spectrophotometer. Anal. Biochem. 1986, 153, 262–266. [Google Scholar] [CrossRef]
- Kim, T.-K.; Yong, H.I.; Jeong, C.H.; Han, S.G.; Kim, Y.-B.; Paik, H.-D.; Choi, Y.-S. Technical Functional Properties of Water- and Salt-soluble Proteins Extracted from Edible Insects. Food Sci. Anim. Resour. 2019, 39, 643–654. [Google Scholar] [CrossRef]
- Baigts-Allende, D.; Doost, A.S.; Ramírez-Rodrigues, M.; Dewettinck, K.; Van der Meeren, P.; de Meulenaer, B.; Tzompa-Sosa, D. Insect protein concentrates from Mexican edible insects: Structural and functional characterization. LWT 2021, 152, 112267. [Google Scholar] [CrossRef]
- Gorissen, S.H.M.; Crombag, J.J.R.; Senden, J.M.G.; Waterval, W.A.H.; Bierau, J.; Verdijk, L.B.; van Loon, L.J.C. Protein content and amino acid composition of commercially available plant-based protein isolates. Amino Acids 2018, 50, 1685–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO/WHO. Food Labelling. Available online: http://www.fao.org/documents/card/en/c/5eb2d33b-d2ad-505e-898a-6aeba60ec896/ (accessed on 11 June 2022).
- Kessler, A.T.; Raja, A. Biochemistry, Histidine; StatPearls: Treasure Island, FL, USA, 2019. [Google Scholar]
- Kohlmeier, M. Asparagine. In Nutrient Metabolism; Kohlmeier, M., Ed.; Academic Press: London, UK, 2003; pp. 389–395. [Google Scholar]
- Bunda, M.G.; Tumbokon, B.L.; Serrano, A.E., Jr. Composition, chemical score (CS) and essential amino acid index (EAAI) of the crinkle grass Rhizoclonium sp. as ingredient for aquafeeds. Aquac. Aquar. Conserv. Legis. 2015, 8, 411–420. [Google Scholar]
- Amarender, R.V.; Bhargava, K.; Dossey, A.T.; Gamagedara, S. Lipid and protein extraction from edible insects—Crickets (Gryllidae). LWT 2020, 125, 109222. [Google Scholar] [CrossRef]
- Mosse, J. Nitrogen-to-protein conversion factor for ten cereals and six legumes or oilseeds. A reappraisal of its definition and determination. Variation according to species and to seed protein content. J. Agric. Food Chem. 1990, 38, 18–24. [Google Scholar] [CrossRef]
- Krul, E.S. Calculation of nitrogen-to-protein conversion factors: A review with a focus on soy protein. J. Am. Oil Chem. Soc. 2019, 96, 339–364. [Google Scholar] [CrossRef]
- Boisen, S.; Bech-Andersen, S.; Eggum, B.O. A Critical View on the Conversion Factor 6.25 from Total Nitrogen to Protein. Acta Agric. Scand. 1987, 37, 299–304. [Google Scholar] [CrossRef]
- WHO. The Joint FAO/WHO Expert Meetings on Nutrition (JEMNU): Nitrogen To Protein Conversion Factors for Soy-Based and Milk-Based Ingredients Used in Infant Formula and Follow-Up Formula: Report of The Meeting of The Expert Panel, Geneva, Switzerland, 16–17 July 2019; WHO: Geneva, Switzerland, 2020.
- Chatsuwan, N.; Nalinanon, S.; Puechkamut, Y.; Lamsal, B.P.; Pinsirodom, P. Characteristics, Functional Properties, and Antioxidant Activities of Water-Soluble Proteins Extracted from Grasshoppers, Patanga succincta and Chondracris roseapbrunner. J. Chem. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Azagoh, C.; Ducept, F.; Garcia, R.; Rakotozafy, L.; Cuvelier, M.-E.; Keller, S.; Lewandowski, R.; Mezdour, S. Extraction and physicochemical characterization of Tenebrio molitor proteins. Food Res. Int. 2016, 88, 24–31. [Google Scholar] [CrossRef]
- Howell, N.K.; Herman, H.; Li-Chan, E.C.Y. Elucidation of Protein−Lipid Interactions in a Lysozyme−Corn Oil System by Fourier Transform Raman Spectroscopy. J. Agric. Food Chem. 2001, 49, 1529–1533. [Google Scholar] [CrossRef]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1767, 1073–1101. [Google Scholar] [CrossRef] [PubMed]
- Pace, C.N.; Fu, H.; Fryar, K.L.; Landua, J.; Trevino, S.R.; Shirley, B.A.; Hendricks, M.M.; Iimura, S.; Gajiwala, K.; Scholtz, J.M.; et al. Contribution of Hydrophobic Interactions to Protein Stability. J. Mol. Biol. 2011, 408, 514–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gravel, A.; Doyen, A. The use of edible insect proteins in food: Challenges and issues related to their functional properties. Innov. Food Sci. Emerg. Technol. 2020, 59, 102272. [Google Scholar] [CrossRef]
- Foo, C.W.P.; Bini, E.; Hensman, J.; Knight, D.; Lewis, R.; Kaplan, D. Role of pH and charge on silk protein assembly in insects and spiders. Appl. Phys. A 2006, 82, 223–233. [Google Scholar] [CrossRef]
- Trevino, S.R.; Scholtz, J.M.; Pace, C.N. Amino Acid Contribution to Protein Solubility: Asp, Glu, and Ser Contribute more Favorably than the other Hydrophilic Amino Acids in RNase Sa. J. Mol. Biol. 2007, 366, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Fennema, O.R. Food Chemistry; CRC Press: Boca Raton, FL, USA, 1996; Volume 76. [Google Scholar]
- Sathe, S.K.; Deshpande, S.S.; Salunkhe, D.K. Functional Properties of Winged Bean [Psophocarpus tetragonolobus (L.) DC] Proteins. J. Food Sci. 1982, 47, 503–509. [Google Scholar] [CrossRef]
- El Nasri, N.A.; El Tinay, A. Functional properties of fenugreek (Trigonella foenum graecum) protein concentrate. Food Chem. 2007, 103, 582–589. [Google Scholar] [CrossRef]
- Zielińska, E.; Karaś, M.; Baraniak, B. Comparison of functional properties of edible insects and protein preparations thereof. LWT 2018, 91, 168–174. [Google Scholar] [CrossRef]
- Omotoso, O.T. An Evaluation of the Nutrients and Some Anti-Nutrients in Silkworm, Bombyxmori L. (Bombycidae: Lepidoptera). Jordan J. Biol. Sci. 2015, 8, 45–50. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, J.; Murray, B.; Flynn, C.; Norton, I. The effect of ultrasound treatment on the structural, physical and emulsifying properties of animal and vegetable proteins. Food Hydrocoll. 2016, 53, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Mishyna, M.; Martinez, J.-J.I.; Chen, J.; Davidovich-Pinhas, M.; Benjamin, O. Heat-induced aggregation and gelation of proteins from edible honey bee brood (Apis mellifera) as a function of temperature and pH. Food Hydrocoll. 2019, 91, 117–126. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Huhu Larvae Protein Extract | Huhu Pupae Protein Extract | T. molitor Larvae | T. molitor Pupae | Beef | Chickpea | Adult Daily Requirement 1985 FAO/WHO/UN |
|---|---|---|---|---|---|---|---|
| Essential | |||||||
| Histidine (His) | 20.8 ± 6.4 | 19.1 ±2.3 | 22.4 | 19.0 | 29.4 | 27.1 | 15 |
| Isoleucine (Ile) | 50.9 ± 1.8 | 55.6 ± 3.6 | 19.7 | 16.1 | 38.4 | 78.9 | 30 |
| Phenylalanine (Phe) | 42.7 ± 1.5 | 47.7 ± 3.4 | 18.8 | 15.3 | 30.9 | 60.8 | 30 |
| Threonine (Thr) | 42.2 ± 1.9 | 40.9 ± 2.5 | 21.5 | 20.3 | 34.3 | 38.5 | 23 |
| Lysine (Lys) | 76.9 ± 2.4 | 75.5 ± 4.3 | 32.3 | 31.5 | 66.6 | 73.7 | 45 |
| Leucine (Leu) | 81.7 ± 1.9 | 91.3 ± 6.1 | 29.7 | 22.1 | 61.8 | 78.9 | 59 |
| Valine (Val) | 53.9 b ± 2.3 | 64.5 a ± 4.4 | 34.4 | 32.1 | 44.8 | 46.8 | 39 |
| Methionine (Met) | 17.5 ± 1.2 | 17.0 ± 1.1 | 5.9 | 4.4 | 23.7 | 15.7 | 22 (Met + Cys) |
| Total essential amino acid (TEAA) | 386.7 ± 9.1 | 411.7 ± 26.8 | 184.7 | 160.8 | 329.9 | 420.4 | 263 |
| EAAI | 3.3 ± 0.02 | 3.4 ± 0.03 | 1.6 | 1.3 | 3.0 | 3.5 | |
| Non-essential | |||||||
| Aspartic acid (Asp) | 96.2 ± 4.4 | 102.2 ± 7.6 | 46.4 | 44.3 | - | 59.4 | |
| Cysteine (Cys) | 12.3 ± 2.7 | 15.2 ± 0.9 | 5.5 | 4.6 | 10.1 | 34.9 | |
| Glycine (Gly) | 41.8 ± 1.2 | 39.4 ± 2.9 | 26.1 | 24.2 | 31.0 | 41.4 | |
| Arginine (Arg) | 60.5 ± 1.6 | 57.7 ± 3.1 | 33.2 | 29.3 | 47.9 | 86.3 | |
| Alanine (Ala) | 54.9 a ± 1.3 | 43.9 b ± 2.8 | 46.4 | 44.3 | 42.2 | 43.5 | |
| Serine (Ser) | 45.1 ± 1.4 | 45.3 ± 3.6 | 23.4 | 20.9 | 32.0 | 52.0 | |
| Tyrosine (Tyr) | 58.3 b ± 2.9 | 81.8 a ± 4.6 | 35.0 | 30.6 | 27.1 | 11.8 | |
| Glutamine (Glu) | 94.0 ± 23.2 | 87.7 ± 7.0 | 65.7 | 60.4 | 46.8 | 83.3 | |
| Proline (Pro) | 50.8 ± 3.4 | 48.9 ± 4.2 | 40.4 | 35.1 | 30.0 | 42.9 | |
| Taurine (Tau) | 7.1 ± 0.2 | 6.9 ± 0.3 | - | - | - | - | |
| Asparagine (Asn) | 2.5 ± 0.2 | 2.5 ± 0.2 | - | - | - | - | |
| Sum of total amino acids | 910.0 ± 23.4 | 943.1 ± 62.3 | - | - | - | - |
| Huhu Grub Extract Source | True Protein (%, DW) | Ntotal | Nprotein | kA | kP | Nitrogen-to-Protein Conversion Factor, k * | Crude Protein Content (%) | |
|---|---|---|---|---|---|---|---|---|
| New k | kP (6.25) | |||||||
| Larvae | 70.1 a ± 3.6 | 11.9 b ± 0.0 | 11.3 a ± 0.5 | 6.2 a ± 0.1 | 5.9 a ± 0.0 | 6.1 a ± 0.2 | 72.1 b ± 1.7 | 74.3 b ± 0.1 |
| Pupae | 75.7 a ± 4.6 | 12.1 a ± 0.0 | 11.8 a ± 1.5 | 6.4 a ± 0.4 | 6.3 a ± 0.8 | 6.4 a ± 0.4 | 76.5 a ± 4.8 | 75.3 a ± 0.1 |
| HLPE | HPPE | ||
|---|---|---|---|
| Colour | L* | 38.4 a ± 1.4 | 32.8 b ± 0.6 |
| a* | 4.0 a ± 0.4 | 4.3 a ± 0.1 | |
| b* | 15.8 a ± 0.4 | 14.5 b ± 0.2 | |
| BI (browning index) | 60.0 a ± 3.3 | 66.4 a ± 2.8 | |
| Water-holding capacity (WHC) (g/mL) | 9.1 a ± 0.5 | 7.3 b ± 0.2 | |
| Oil-holding capacity (OHC) (g/mL) | 7.9 a ± 0.8 | 5.1 b ± 0.9 | |
| Surface hydrophobicity | 35.5 a ± 1.7 | 36.3 a ± 1.4 | |
| Polydispersity index (PDI) | 0.5 a ± 0.0 | 0.6 a ± 0.1 | |
| Hydrodynamic diameter (dh, nm) | 255.6 b ± 8.1 | 1078.7 a ± 22.2 | |
| Emulsion capacity (%) | 82.2 a ± 1.9 | 77.8 a ± 4.8 | |
| Heat coagulation at pH 5 (%) | 4.5 b ± 0.6 | 7.5 a ± 0.7 | |
| Heat coagulation at pH 7 (%) | 17.6 a ± 3.6 | 10.5 b ± 2.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kavle, R.R.; Nolan, P.J.; Bekhit, A.E.-D.A.; Carne, A.; Morton, J.D.; Agyei, D. Physicochemical Characteristics, Techno-Functionalities, and Amino Acid Profile of Prionoplus reticularis (Huhu) Larvae and Pupae Protein Extracts. Foods 2023, 12, 417. https://doi.org/10.3390/foods12020417
Kavle RR, Nolan PJ, Bekhit AE-DA, Carne A, Morton JD, Agyei D. Physicochemical Characteristics, Techno-Functionalities, and Amino Acid Profile of Prionoplus reticularis (Huhu) Larvae and Pupae Protein Extracts. Foods. 2023; 12(2):417. https://doi.org/10.3390/foods12020417
Chicago/Turabian StyleKavle, Ruchita Rao, Patrick James Nolan, Alaa El-Din Ahmed Bekhit, Alan Carne, James David Morton, and Dominic Agyei. 2023. "Physicochemical Characteristics, Techno-Functionalities, and Amino Acid Profile of Prionoplus reticularis (Huhu) Larvae and Pupae Protein Extracts" Foods 12, no. 2: 417. https://doi.org/10.3390/foods12020417