
Characterization of the Technofunctional Properties and Three-Dimensional Structure Prediction of 11S Globulins from Amaranth (Amaranthus hypochondriacus L.) Seeds

 , ,
, ,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and methods
2.1. Materials and Chemical Reactants
2.2. Purification of 11S Globulins from Amaranth Seeds
2.3. Protein Quantification
2.4. Protein in-Gel Digestion, Mass Spectrometry Analysis, and Protein Identification
2.5. Protein Structure Prediction of Amaranth 11S Globulins
2.6. Dynamic Light Scattering (DLS) Analysis
2.7. Surface Hydrophobicity
2.8. Solubility
2.9. Emulsifying Properties
2.10. Statistical Analysis
3. Results and Discussions
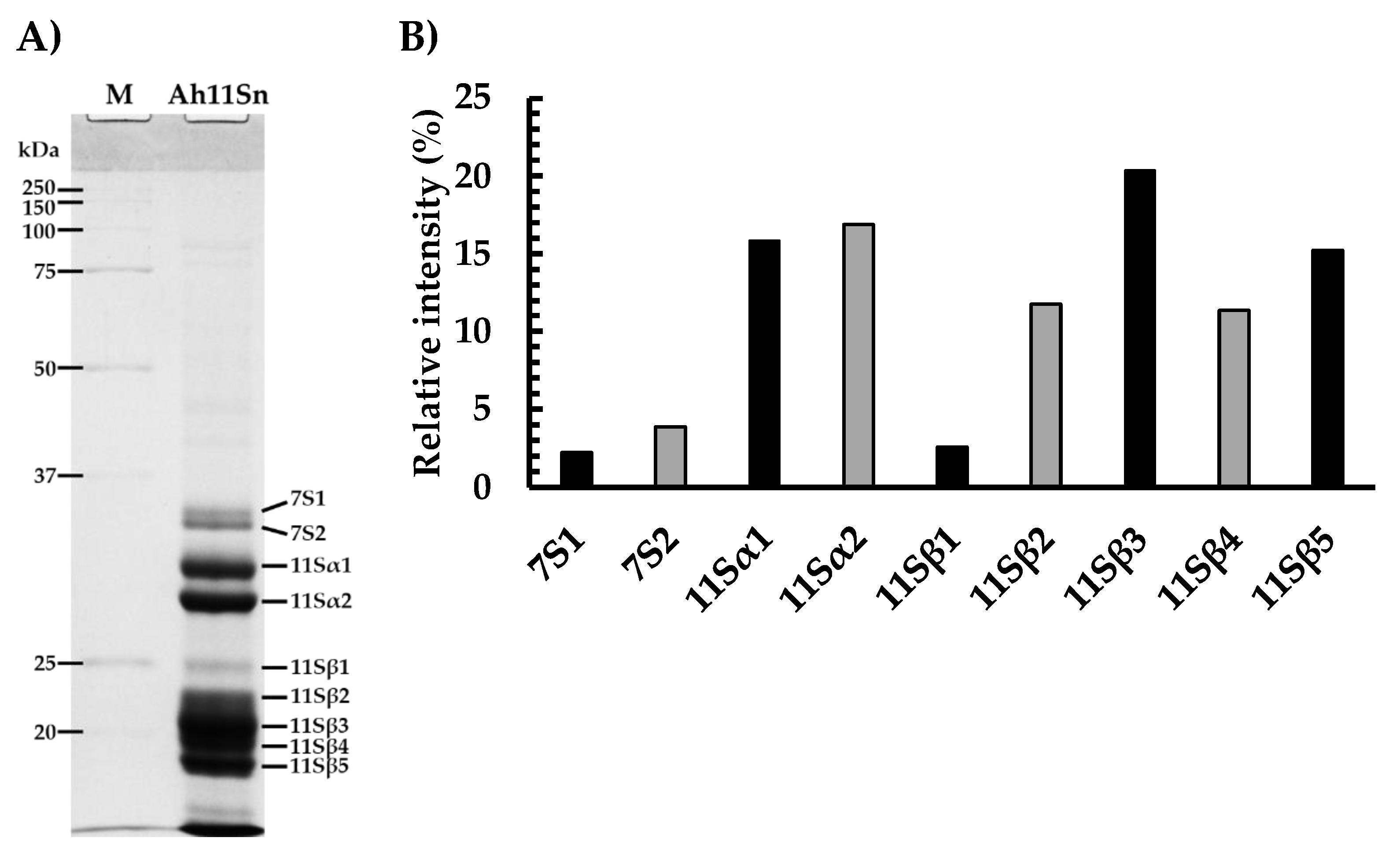
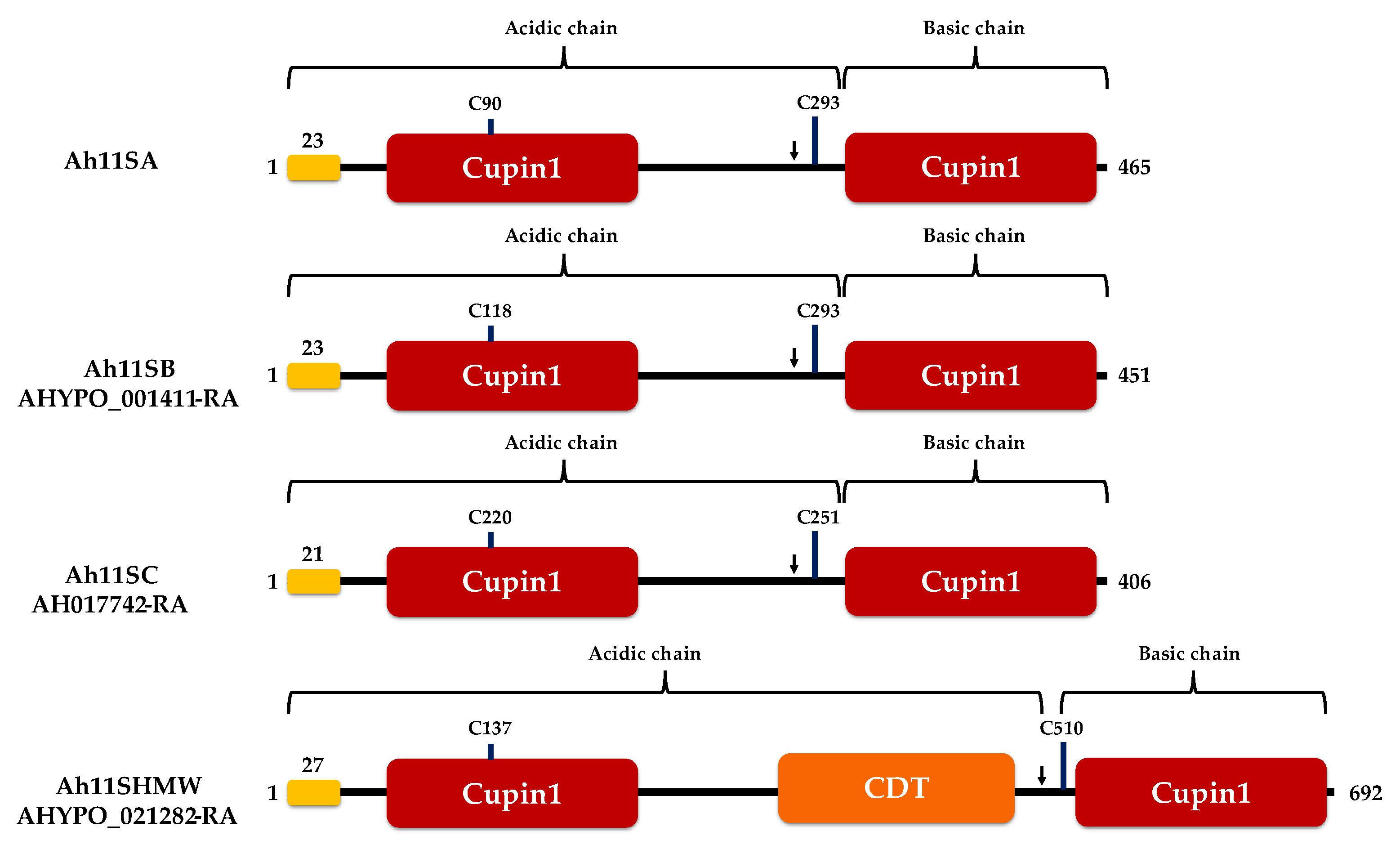
3.1. Purification and LC–MS Identification of Amaranth 11S Globulins
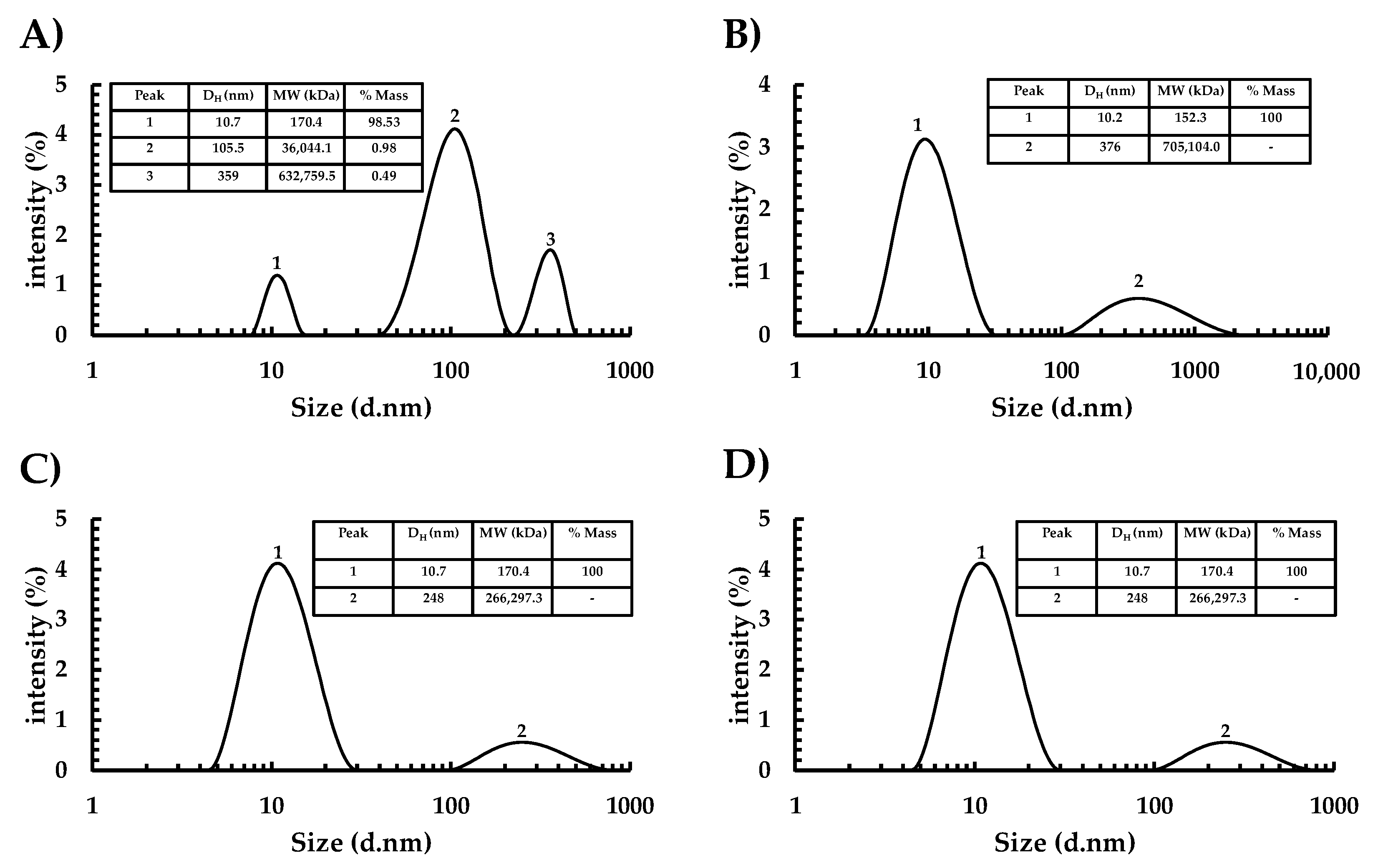
3.2. Assembly Capacity
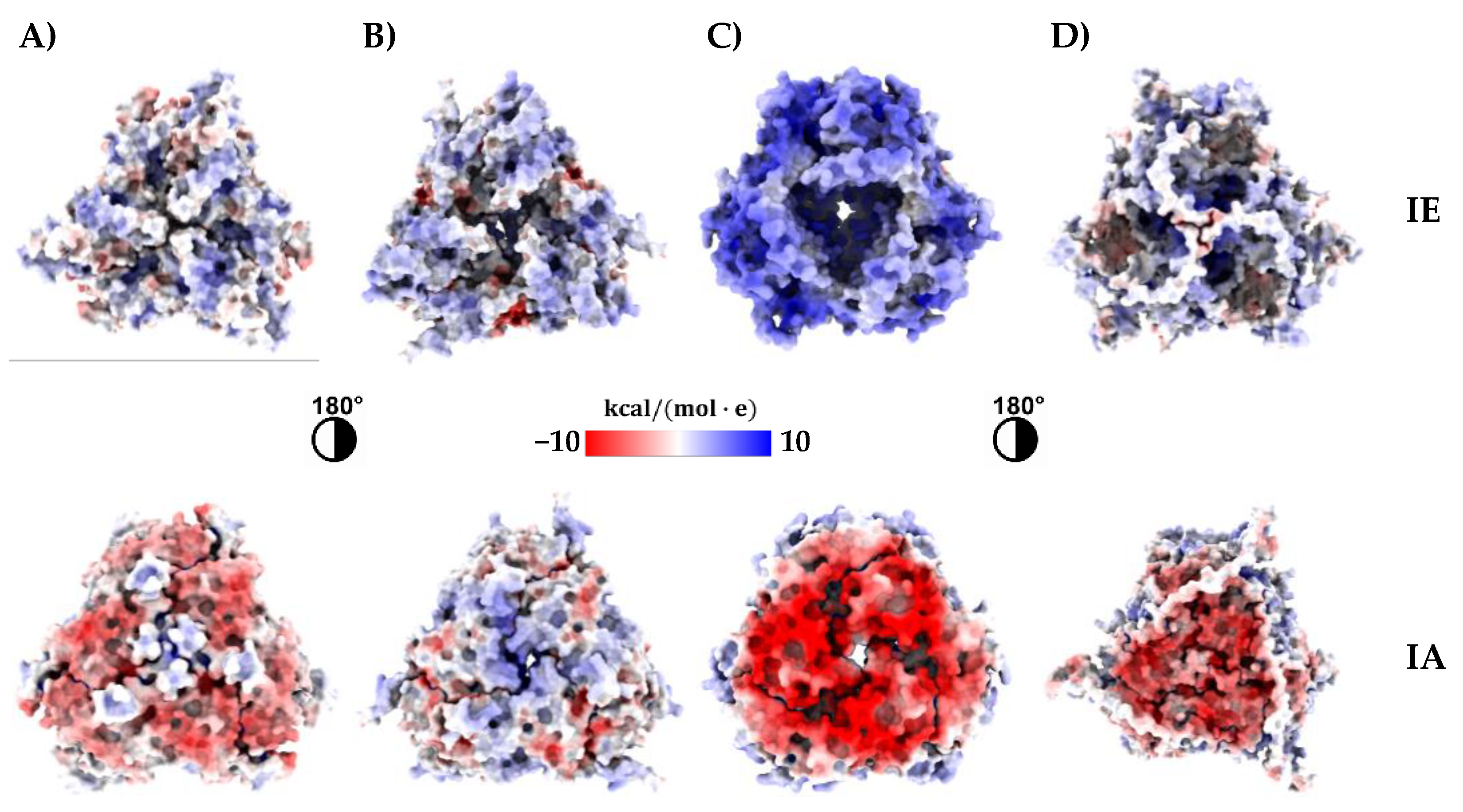
3.3. Three-Dimensional Structure Prediction of Amaranth 11S Globulins
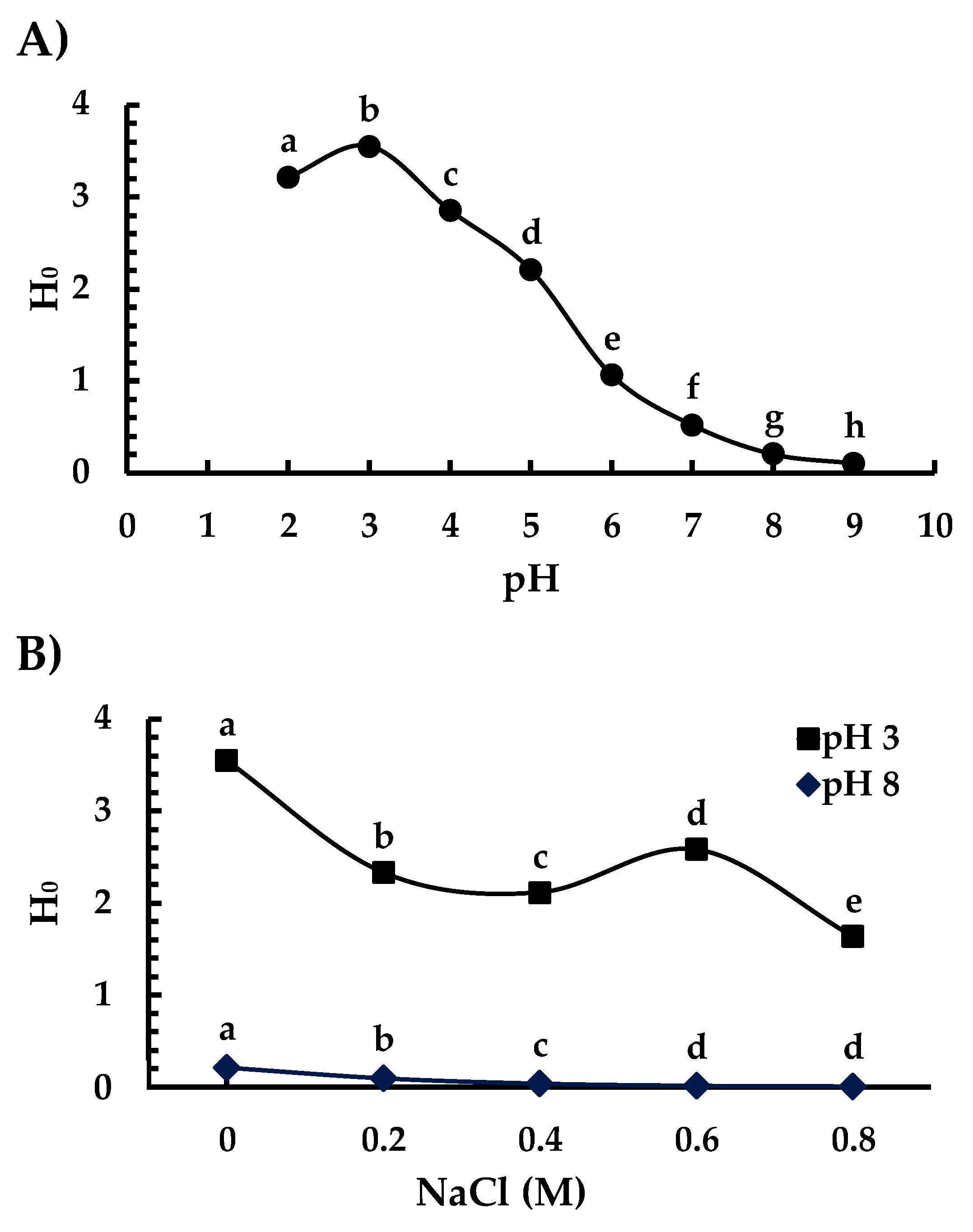
3.4. Surface Hydrophobicity
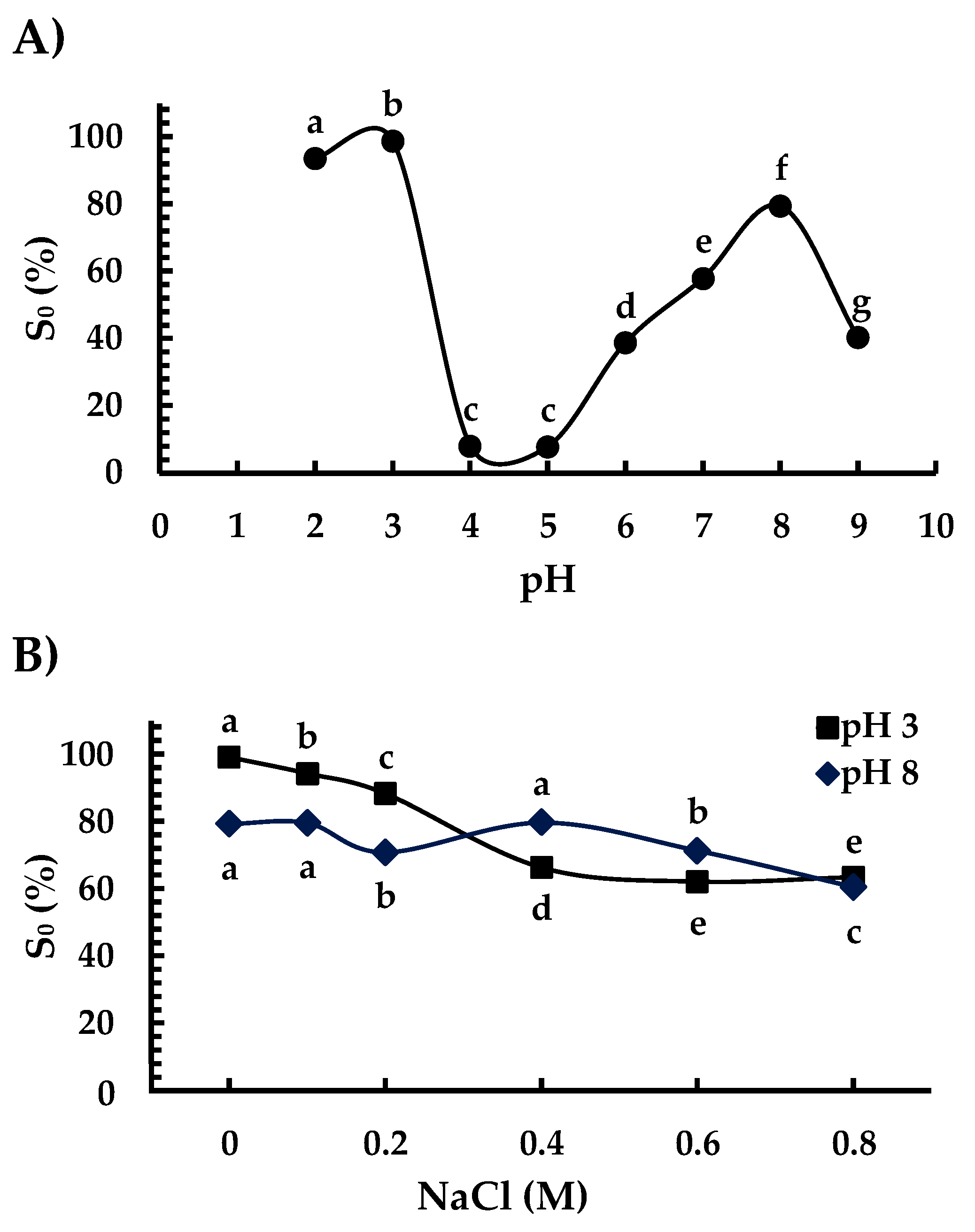
3.5. Solubility
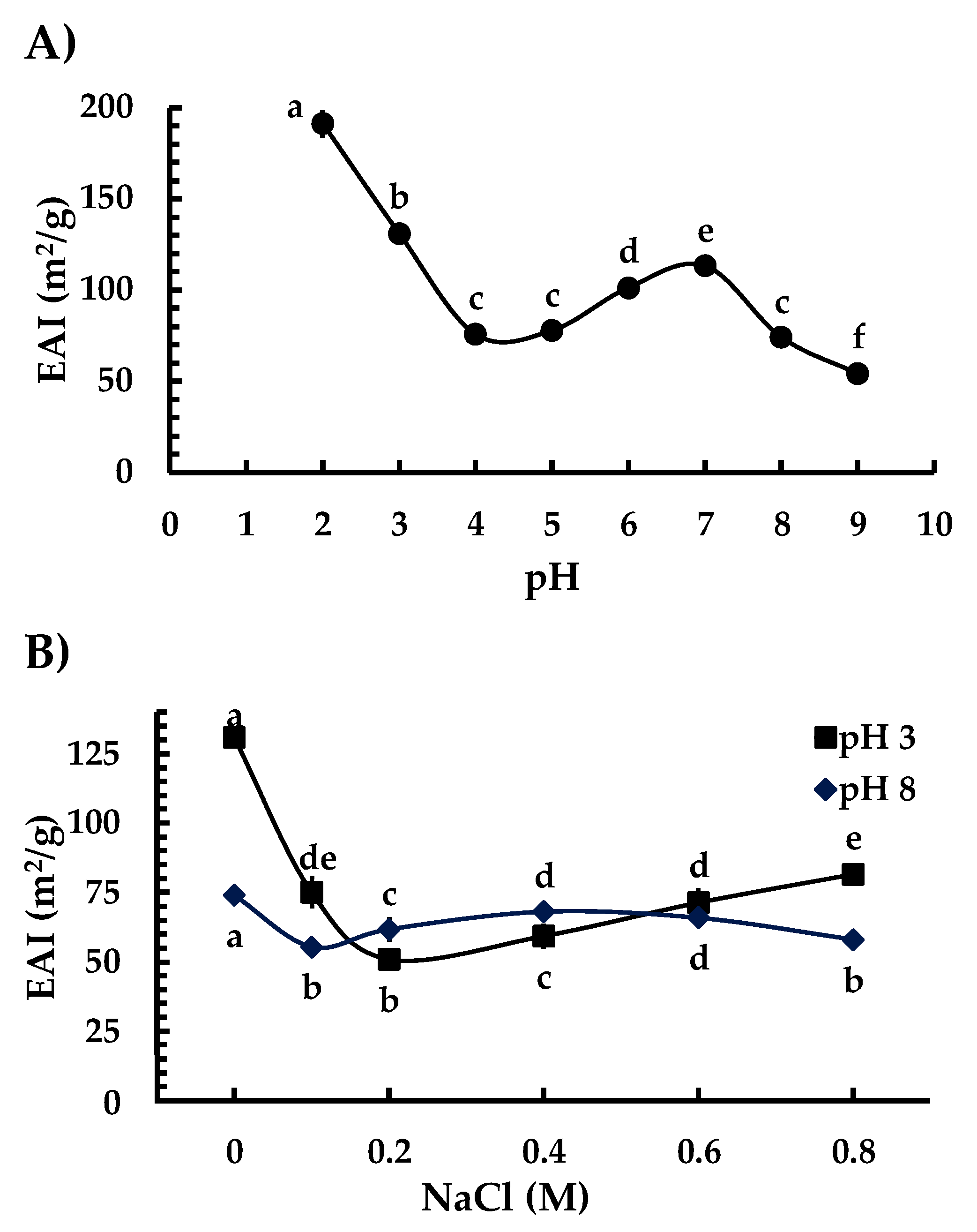
3.6. Emulsifying Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barba de la Rosa, A.P.; Paredes-López, O.; Gueguen, J.; Viroben, G. Fractionation procedures, electrophoretic characterization, and amino acid composition of amaranth seed proteins. J. Agric. Food Chem. 1992, 40, 931–936. [Google Scholar] [CrossRef]
- Barba de la Rosa, A.P.; Barba Montoya, A.; Martínez-Cuevas, P.; Hernández-Ledesma, B.; León-Galván, M.F.; de León-Rodríguez, A.; González, C. Tryptic amaranth glutelin digests induce endothelial nitric oxide production through inhibition of ACE: Antihypertensive role of amaranth peptides. Nitric Oxide 2010, 23, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Cervantes, E.; Jeong, H.J.; León-Galván, F.; Barrera-Pacheco, A.; de León-Rodríguez, A.; González De Mejia, E.; de Lumen, B.O.; Barba De La Rosa, A.P. Amaranth lunasin-like peptide internalizes into the cell nucleus and inhibits chemical carcinogen-induced transformation of NIH-3T3 cells. Peptides 2010, 31, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
- Silva-Sánchez, C.; Barba De La Rosa, A.P.; León-Galván, M.F.; de Lumen, B.O.; de León-Rodríguez, A.; González De Mejía, E. Bioactive peptides in amaranth (Amaranthus hypochondriacus) seed. J. Agric. Food Chem. 2008, 56, 1233–1240. [Google Scholar] [CrossRef]
- Aguilar-Hernández, H.S.; Santos, L.; León-Galván, F.; Barrera-Pacheco, A.; Espitia-Rangel, E.; de León-Rodríguez, A.; Guevara-González, R.G.; Barba de la Rosa, A.P. Identification of calcium stress induced genes in amaranth leaves through suppression subtractive hybridization. J. Plant Physiol. 2011, 168, 2102–2109. [Google Scholar] [CrossRef]
- Huerta-Ocampo, J.A.; León-Galván, M.F.; Ortega-Cruz, L.B.; Barrera-Pacheco, A.; de León-Rodríguez, A.; Mendoza-Hernández, G.; Barba de la Rosa, A.P. Water stress induces up-regulation of DOF1 and MIF1 transcription factors and down-regulation of proteins involved in secondary metabolism in amaranth roots (Amaranthus hypochondriacus L.). Plant Biol. 2011, 13, 472–482. [Google Scholar] [CrossRef]
- Omamt, E.N.; Hammes, P.S.; Robbertse, P.J. Differences in salinity tolerance for growth and water-use efficiency in some amaranth (Amaranthus spp.) genotypes. N. Z. J. Crop Hortic. Sci. 2010, 34, 11–22. [Google Scholar] [CrossRef]
- Kesari, P.; Neetu BS, P.; Sharma, A.; Katiki, M.; Kumar, P.; Gurjar, B.; Tomar, S.; Sharma, A.; Kumar, P. Structural, functional and evolutionary aspects of seed globulins. Protein Pept. Lett. 2017, 24, 267–277. [Google Scholar] [CrossRef]
- Tandang-Silvas MR, G.; Tecson-Mendoza, E.M.; Mikami, B.; Utsumi, S.; Maruyama, N. Molecular design of seed storage proteins for enhanced food physicochemical properties. Annu. Rev. Food Sci. Technol. 2011, 2, 59–73. [Google Scholar] [CrossRef]
- Kinsella, J.E.; Melachouris, N. Functional properties of proteins in foods: A survey. CRC Crit. Rev. Food Sci. Nutr. 1976, 7, 219–280. [Google Scholar] [CrossRef]
- Carrazco-Peña, L.; Osuna-Castro, J.A.; de León-Rodríguez, A.; Maruyama, N.; Toro-Vazquez, J.F.; Morales-Rueda, J.A.; Barba De La Rosa, A.P. Modification of solubility and heat-induced gelation of Amaranth 11S Globulin by protein engineering. J. Agric. Food Chem. 2013, 61, 3509–3516. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bojórquez-Velázquez, E.; Elizalde-Contreras, J.M.; Zamora-Briseño, J.A.; Ruiz-May, E. Efficient Protein Extraction Protocols for NanoLC-MS/MS Proteomics Analysis of Plant Tissues with High Proteolytic Activity: A Case Study with Pineapple Pulp. In Plant Comparative Genomics. Methods in Molecular Biology; Pereira-Santana, A., Gamboa-Tuz, S.D., Rodríguez-Zapata, L.C., Eds.; Humana: New York, NY, USA, 2022; Volume 2512. [Google Scholar] [CrossRef]
- Clouse, J.W.; Adhikary, D.; Page, J.T.; Ramaraj, T.; Deyholos, M.K.; Udall, J.A.; Fairbanks, D.J.; Jellen, E.N.; Maughan, P.J. The Amaranth Genome: Genome, transcriptome, and physical map assembly. Plant Genome 2016, 9. [Google Scholar] [CrossRef] [Green Version]
- Lightfoot, D.J.; Jarvis, D.E.; Ramaraj, T.; Lee, R.; Jellen, E.N.; Maughan, P.J. Single-molecule sequencing and Hi-C-based proximity-guided assembly of amaranth (Amaranthus hypochondriacus) chromosomes provide insights into genome evolution. BMC Biol. 2017, 15, 74. [Google Scholar] [CrossRef] [Green Version]
- Krieger, E.; Joo, K.; Lee, J.; Lee, J.; Raman, S.; Thompson, J.; Tyka, M.; Baker, D.; Karplus, K. Improving physical realism, stereochemistry, and side-chain accuracy in homology modeling: Four approaches that performed well in CASP8. Proteins Struct. Funct. Bioinform. 2009, 77 (Suppl. 9), 114–122. [Google Scholar] [CrossRef] [Green Version]
- Baek, M.; Park, T.; Heo, L.; Park, C.; Seok, C. GalaxyHomomer: A web server for protein homo-oligomer structure prediction from a monomer sequence or structure. Nucleic Acids Res. 2017, 45, W320–W324. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenberg, D.; Lüthy, R.; Bowie, J.U. [20] VERIFY3D: Assessment of protein models with three-dimensional profiles. Methods Enzymol. 1997, 277, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Fiser, A.; Sali, A. ModLoop: Automated modeling of loops in protein structures. Bioinformatics 2003, 19, 2500–2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS biomolecular solvation software suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef] [Green Version]
- De la Cruz-Torres, L.F.; Rodríguez-Celestino, V.; Centeno-Leija, S.; Serrano-Posada, H.; Ceballos-Magaña, S.G.; Aguilar-Padilla, J.; Mancilla-Margalli, N.A.; Osuna-Castro, J.A. Development of a rapid, high-sensitivity, low-cost fluorescence method for protein surface hydrophobicity determination using a Nanodrop fluorospectrometer. Food Chem. 2022, 396, 133681. [Google Scholar] [CrossRef]
- Kato, A.; Nakai, S. Hydrophobicity determined by a fluorescence probe method and its correlation with surface properties of proteins. Biochim. Biophys. Acta (BBA) Protein Struct. 1980, 624, 13–20. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- De la Cruz-Torres, L.F.; Pérez-Martínez, J.D.; Sánchez-Becerril, M.; Toro-Vázquez, J.F.; Mancilla-Margalli, N.A.; Osuna-Castro, J.A.; VillaVelázquez-Mendoza, C.I. Physicochemical and functional properties of 11S globulin from chan (Hyptis suaveolens L. poit) seeds. J. Cereal Sci. 2017, 77, 66–72. [Google Scholar] [CrossRef]
- Pearce, K.N.; Kinsella, J.E. Emulsifying properties of proteins: Evaluation of a turbidimetric technique. J. Agric. Food Chem. 2002, 26, 716–723. [Google Scholar] [CrossRef]
- Bojórquez-Velázquez, E.; Lino-López, G.J.; Huerta-Ocampo, J.A.; Barrera-Pacheco, A.; Barba De La Rosa, A.P.; Moreno, A.; Mancilla-Margalli, N.A.; Osuna-Castro, J.A. Purification and biochemical characterization of 11S globulin from chan (Hyptis suaveolens L. Poit) seeds. Food Chem. 2016, 192, 203–211. [Google Scholar] [CrossRef]
- Tang, C.H.; Sun, X. Physicochemical and structural properties of 8S and/or 11S globulins from mungbean (Vigna radiata (L.) Wilczek] with Various Polypeptide Constituents. J. Agric. Food Chem. 2010, 58, 6395–6402. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Takako, F.; Meili, Z.; Shiori, M.; Maruyama, N.; Utsumi, S. Comparison of physicochemical properties of 7S and 11S globulins from pea, fava bean, cowpea, and french bean with those of soybean—French bean 7S globulin exhibits excellent properties. J. Agric. Food Chem. 2008, 56, 10273–10279. [Google Scholar] [CrossRef] [PubMed]
- Tandang-Silvas, M.R.; Cabanos, C.S.; Carrazco Peña, L.D.; de La Rosa AP, B.; Osuna-Castro, J.A.; Utsumi, S.; Mikami, B.; Maruyama, N. Crystal structure of a major seed storage protein, 11S proglobulin, from Amaranthus hypochondriacus: Insight into its physico-chemical properties. Food Chem. 2012, 135, 819–826. [Google Scholar] [CrossRef]
- Bojórquez-Velázquez, E.; Barrera-Pacheco, A.; Espitia-Rangel, E.; Herrera-Estrella, A.; de La Rosa AP, B. Protein analysis reveals differential accumulation of late embryogenesis abundant and storage proteins in seeds of wild and cultivated amaranth species. BMC Plant Biol. 2019, 19, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan-Wilson, A.L.; Wilson, K.A. Mobilization of Seed Protein Reserves. Physiol. Plant. 2012, 145, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Víctor, M.; Ruiz-Henestrosa, P.; Martinez, M.J.; Patino, J.M.R.; Pilosof, A.M.R. A Dynamic light scattering study on the complex assembly of glycinin soy globulin in aqueous solutions. J. Am. Oil Chem. Soc. 2012, 89, 1183–1191. [Google Scholar] [CrossRef]
- Zhao, Y.; Mine, Y.; Ma, C.Y. Study of thermal aggregation of oat globulin by laser light scattering. J. Agric. Food Chem. 2004, 52, 3089–3096. [Google Scholar] [CrossRef]
- Adachi, M.; Kanamori, J.; Masuda, T.; Yagasaki, K.; Kitamura, K.; Mikami, B.; Utsumi, S. Crystal structure of soybean 11S globulin: Glycinin A3B4 homohexamer. Proc. Natl. Acad. Sci. USA 2003, 100, 7395–7400. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Zhou, A.; Zhang, F. Crystallization and crystallographic studies of a novel chickpea 11S globulin. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2022, 78, 324–329. [Google Scholar] [CrossRef]
- Withana-Gamage, T.S.; Hegedus, D.D.; McIntosh, T.C.; Coutu, C.; Qiu, X.; Wanasundara, J.P.D. Subunit composition affects formation and stabilization of o/w emulsions by 11S seed storage protein cruciferin. Food Res. Int. 2020, 137, 109387. [Google Scholar] [CrossRef]
- Deng, Q.; Wang, L.; Wei, F.; Xie, B.; Huang, F.H.; Huang, W.; Shi, J.; Huang, Q.; Tian, B.; Xue, S. Functional properties of protein isolates, globulin and albumin extracted from Ginkgo biloba seeds. Food Chem. 2011, 124, 1458–1465. [Google Scholar] [CrossRef]
- González-González, R.M.; Peraza-Campos, A.L.; Ceballos-Magaña, S.G.; Rodríguez-Pérez, M.A.; Sañudo-Barajas, J.A.; Parra-Delgado, H.; Osuna-Castro, J.A. Biochemical and functional characterization of albumins and globulins of Brosimum alicastrum seeds. J. Food Process. Preserv. 2020, 44, e14679. [Google Scholar] [CrossRef]
- De la Cruz-Torres, L.F.; Mancilla-Margalli, N.A.; VillaVelázquez-Mendoza, C.I.; Carrazco-Peña, L.D.; Chan-Cupul, W.; Osuna-Castro, J.A.; Toro-Vázquez, J.F.; Pérez-Martínez, J.D. Thermal and emulsifying properties of globulins from chan (Hyptis suaveolens L. Poit) seeds. J. Food Process. Preserv. 2020, 44, e14652. [Google Scholar] [CrossRef]
- Tandang MR, G.; Atsuta, N.; Maruyama, N.; Adachi, M.; Utsumi, S. Evaluation of the solubility and emulsifying property of soybean proglycinin and rapeseed procruciferin in relation to structure modified by protein engineering. J. Agric. Food Chem. 2005, 53, 8736–8744. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar-Padilla, J.; Centeno-Leija, S.; Bojórquez-Velázquez, E.; Elizalde-Contreras, J.M.; Ruiz-May, E.; Serrano-Posada, H.; Osuna-Castro, J.A. Characterization of the Technofunctional Properties and Three-Dimensional Structure Prediction of 11S Globulins from Amaranth (Amaranthus hypochondriacus L.) Seeds. Foods 2023, 12, 461. https://doi.org/10.3390/foods12030461
Aguilar-Padilla J, Centeno-Leija S, Bojórquez-Velázquez E, Elizalde-Contreras JM, Ruiz-May E, Serrano-Posada H, Osuna-Castro JA. Characterization of the Technofunctional Properties and Three-Dimensional Structure Prediction of 11S Globulins from Amaranth (Amaranthus hypochondriacus L.) Seeds. Foods. 2023; 12(3):461. https://doi.org/10.3390/foods12030461
Chicago/Turabian StyleAguilar-Padilla, Jorge, Sara Centeno-Leija, Esaú Bojórquez-Velázquez, José M. Elizalde-Contreras, Eliel Ruiz-May, Hugo Serrano-Posada, and Juan Alberto Osuna-Castro. 2023. "Characterization of the Technofunctional Properties and Three-Dimensional Structure Prediction of 11S Globulins from Amaranth (Amaranthus hypochondriacus L.) Seeds" Foods 12, no. 3: 461. https://doi.org/10.3390/foods12030461