Abstract
Enterococcus faecium FUA027 transforms ellagic acid (EA) to urolithin A (UA), which makes it a potential application in the preparation of UA by industrial fermentation. Here, the genetic and probiotic characteristics of E. faecium FUA027 were evaluated through whole-genome sequence analysis and phenotypic assays. The chromosome size of this strain was 2,718,096 bp, with a GC content of 38.27%. The whole-genome analysis revealed that the genome contained 18 antibiotic resistance genes and seven putative virulence factor genes. E. faecium FUA027 does not contain plasmids and mobile genetic elements (MGEs), and so the transmissibility of antibiotic resistance genes or putative virulence factors should not occur. Phenotypic testing further indicated that E. faecium FUA027 is sensitive to clinically relevant antibiotics. In addition, this bacterium exhibited no hemolytic activity, no biogenic amine production, and could significantly inhibit the growth of the quality control strain. In vitro viability was >60% in all simulated gastrointestinal environments, with good antioxidant activity. The study results suggest that E. faecium FUA027 has the potential to be used in industrial fermentation for the production of urolithin A.
1. Introduction
Ellagitannins (ETs), the metabolic precursor of urolithins, can be hydrolyzed to ellagic acid (EA), which is subsequently metabolized by gut microorganisms to urolithins [1]. Among all types of those urolithins, urolithin A (UA) exhibited several potentially positive bioactivities, such as restoring muscle function [2], and antiobesity [3], antioxidant [4], anti-inflammation, and anticancer activities [5]. An increasing amount of the literature has recently focused on the impact of the natural compound UA on health, disease, and aging [6]. Numerous studies have shown that different urolithin metabotypes (UMs) produce significantly different amounts and types of urolithins [7]. The gut microflora in more than 40% of middle-aged and elderly people cannot metabolize EA to UA [8]. Cortés et al. found that the percentage of the UM-A population declines when the intestinal flora changes with age [9]. Given the influence of intestinal flora on UA formation [10], screening strains responsible for metabolizing EA to produce UA is of interest.
Currently, little is known about the species of gut bacteria involved in EA conversion to UA. Strains found to metabolize EA to produce UA include Bifidobacterium pseudocatenulatum INIA P815 [11], Streptococcus thermophilus FUA329 [12], Lactococcus garvieae FUA009 [13], and Enterococcus faecium FUA027 [14]. S. thermophilus FUA329 was isolated from human milk. L. garvieae FUA009 and E. faecium FUA027 were screened from fecal samples. These bacteria have the potential to be developed as probiotics for the in vitro biotransformation of EA to produce UA, or for industrial fermentation to produce UA [15].
Our previous studies have proven that E. faecium FUA027, which was isolated from human fecal samples, metabolizes EA to UA by detecting UA from the fermentation broth of the strain through high-performance liquid chromatography (HPLC) and liquid chromatography tandem mass spectrometry (LC-MS/MS). The highest yield of UA produced by E. faecium FUA027 was 10.80 μM, thereby making this strain a promising candidate for development as a probiotic [14].
The safety and probiotic properties of the strain to be used as probiotics must be evaluated [16]. In this study, whole-genome sequence information analysis and phenotypic assays were used in combination to assess antibiotic resistance, metabolite toxicity, and survival under simulated gastrointestinal conditions. The safety of E. faecium FUA027 and its potential for use in the preparation of UA by industrial fermentation were confirmed.
2. Materials and Methods
2.1. Bacterial Strain and Growth Conditions
E. faecium FUA027 was preserved in the China General Microbiological Culture Collection Center (CGMCC) under the accession number CGMCC No. 24964. All FUA027 strains, unless otherwise noted, were cultivated in Anaerobe Basal Broth (ABB) medium and incubated under anaerobic conditions consisting of N2/H2/CO2 (80:10:10, v:v:v) at 37 °C for 24 h. Staphylococcus aureus ATCC 12600, Escherichia coli ATCC 25922, Yeast ATCC 24060, Aspergillus niger ATCC 6273, and Lactobacillus plantarum ATCC 4008 strains were used partly for inhibition experiments and partly as control strains in the experiments. S. aureus and E. coli were cultured at 37 °C in Luria–Bertani broth for 24 h. Yeast and A. niger were cultured on potato dextrose agar medium at 37 °C for 48 h. L. plantarum and S. thermophilus were cultivated in Man Rogosa Sharpe broth at 37 °C for 48 h.
2.2. Whole-Genome Sequencing
The genomic DNA was extracted from the E. faecium FUA027 culture grown in ABB by using a bacterial DNA extraction kit from Sangon, Shanghai, Co. Ltd. (Shanghai, China). For the DNA sample preparations, 1 µg DNA per sample was used as the input material. Sequencing libraries were created using the NEBNext® Ultra™ DNA Library Prep Kit for Illumina (New England Biolabs, Ipswich, MA, USA) according to the manufacturer’s instructions. In brief, the DNA sample was sonicated to obtain 350-bp fragments. The DNA fragments were end-polished, A-tailed, and ligated with the full-length adaptor for Illumina sequencing with further PCR amplification. Finally, the AMPure XP system purified the PCR products, and the size distribution of the libraries was analyzed using the Agilent 2100 Bioanalyzer and quantified using real-time PCR. The whole genome of FUA027 was sequenced using the Nanopore PromethION platform and Illumina NovaSeq PE150 at the Beijing Novogene Bioinformatics Technology Co., Ltd. (Beijing, China).
2.3. Genome Assembly and Annotation
The trimmed data for the FUA027 genome were combined with PE150 and Nanopore data and assembled using SMRT Link v5.0.1 software (https://www.pacb.com/support/software-downloads/, accessed on 15 October 2022). The quality of the genome assembly was validated using QUAST ver. 5.0.2. The final assembly was annotated using the NCBI Prokaryotic Genome Annotation Pipeline (http://www.ncbi.nlm.nih.gov/genome/annotation_prok/, accessed on 15 October 2022) [17]. We used Gene Ontology (GO), the Kyoto Encyclopedia of Genes and Genomes (KEGG), Clusters of Orthologous Groups (COG), the Non-Redundant Protein Database, the Transporter Classification Database, and Swiss-Prot to predict gene function.
2.4. Strain Safety Assessment
2.4.1. Identifying Safety-Related Genes from the FUA027 Genome
Bacterial virulence factors were identified by referring to the virulence factor database updated in 2019 (VFDB, http://www.mgc.ac.cn/VFs/, accessed on 11 October 2022) [18]. Protein sequences with >50% similarity in the extraction comparison results were identified as virulence genes. Antimicrobial resistance determinant identification was performed using the ABRicate program (https://github.com/tseemann/abricate, accessed on 11 October 2022) based on the ResFinder database (http://genomicepidemiology.org/, accessed on 11 October 2022) [19]. Antibiotic resistance genes of E. faecium FUA027 were identified using the comprehensive antibiotic resistance database (CARD, https://card.mcmaster.ca, accessed on 11 October 2022) [20].
2.4.2. Antibiotic Susceptibility Testing
Susceptibility testing was performed through disk diffusion according to EUCAST recommendations [21]. The strain FUA027 was purified, inoculated into 20 mL of ABB liquid medium, and incubated anaerobically at 37 °C for 24 h. Bacterial colonies were counted, and the concentration of the bacterial solution was adjusted to 1.0 × 108 CFU/mL. The bacterial solution was then added dropwise to a 20 mm agar plate. The FUA329 bacterial solution was evenly coated on the plate. Under aseptic conditions, antibiotic susceptibility papers were gently pressed onto the agar plates using forceps. While doing so, the spacing of each drug-sensitive tablet could not be <20 mm and the distance from the edge of the plate could not be <17 mm. The plates were sealed and continuously incubated at 37 °C for 14 h. The size of the inhibition circle was noted to determine the sensitivity of antibiotics.
2.4.3. Hemolytic Activity Evaluation
The hemolytic activity was studied using the method described by Buxton. In short, E. faecium FUA027 was inoculated onto Columbia Blood Agar and incubated at 37 °C for 24 h [22]. S. aureus ATCC 12600 was used as a control strain.
2.4.4. Nitrate Reductase and Amino Acid Decarboxylase Activity
The nitrate broth assay kit and amino acid decarboxylase assay kit obtained from Beijing Land Bridge Technology Co., Ltd. (Beijing, China). were used in the metabolic toxicity test. The test was performed following the manufacturer’s instructions.
Detection of nitrate reductase activity: Under aseptic conditions, single colonies of the test strain and the quality control strain E. coli ATCC 25922 isolated from the plate were inoculated in a nitrate broth assay ampoule by using an inoculating needle. The plate was incubated at 37 °C for 24 h. After incubation, nitrate reduction reagents A and B were added dropwise at 5:2 (v:v), and the results were observed immediately. Three parallel experiments were conducted for each sample [23].
Detection of amino acid decarboxylase activity: Under aseptic conditions, a single colony of the test strain was picked from the plate by using an inoculating needle and inoculated into the amino acid decarboxylase series ampoule as well as the amino acid decarboxylase control tube. Sterile liquid paraffin was added to cover the surface of the medium, and lysine, ornithine, and arginine ampoules were incubated at 37 °C for 24 h. After the phenylalanine ampoule was incubated for 24 h, 4–5 drops of 10% FeCl3 aqueous solution were added to the ampoules, and the results were observed within 2 min. Following the incubation of the tryptophan ampoules for 24 h, 2–3 drops of the Kovacs reagent were added to the ampoules and the results were observed immediately. Three parallel experiments were conducted for each sample.
2.5. Assessment of Probiotic Properties
2.5.1. Probiotic-Associated Genes in the E. faecium FUA027 Genome
The Hidden Markov model (HMM) was used to find probiotic-associated genes in the genome as well as environmental tolerance-related genes [24]. Additionally, we searched for genes related to adhesion factors in the annotation results.
Putative genes involved in antimicrobial compound synthesis and secondary metabolism gene clusters in the E. faecium FUA027 genome were identified using AntiSMASH 6.0 (https://antismash.secondarymetabolites.org, accessed on 11 December 2022) [25] and BAGEL 4.0 (http://bagel4.molgenrug.nl/index.php, accessed on 11 December 2022) [26].
2.5.2. Evaluation of Acid and Bile Salt Tolerance In Vitro
Referring to Pieniz et al.’s study, the survival of strains in a simulated gastrointestinal environment was measured using the viable plate count method [27]. The strain FUA027 was grown in ABB liquid medium at 37 °C for 24 h. Then, the culture was adjusted to an optical density (OD600) of 1.0 ± 0.05.
Separate preparation of ABB liquid medium of different pH values and containing different bile salt concentrations: test tubes containing 9 mL of ABB liquid medium were adjusted with HCl to attain different pH values (i.e., 2.0, 2.5, 3.0, 3.5, and 4.0). The ABB liquid medium was supplemented with bovine bile salt, thereby achieving final concentrations of 0.1%, 0.2%, 0.3%, 0.4%, and 0.5% (w/v), respectively. Then, 1 mL of inoculum was added to each tube, and the normal ABB liquid medium was used as a control. Sampling was performed at 0, 1, 2, and 3 h. The samples were diluted with ABB medium and then coated and incubated on the plates for 24 h, and viable colonies on a plate were counted. The survival rate was calculated using the following formula:
where (log CFU/mL) represents the number of viable bacteria after t hours of treatment, and (log CFU/mL) refers to the number of viable bacteria of E. faecium FUA027 before treatment.
2.5.3. Evaluation of the Antioxidant Activity In Vitro
The FUA027 strain was cultured in ABB liquid medium at 37 °C for 18 h. The E. faecium FUA027 bacterial liquid was centrifuged (20 °C, 3000 rpm, 10 min), then discard supernatant intact cells of the strain were harvested. The cell pellet was washed twice with and suspended in 1 mL sterile distilled water [28]. The concentration of this suspension was adjusted to approximately 1.0 × 108 CFU/mL. This was considered as a sample in the antioxidant test. Using antioxidant kits from Jiancheng Bioengineering Institute (Nanjing, China), in vitro antioxidant activities were measured including the measurement of the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical, hydroxyl radical, and superoxide anion scavenging activities [29].
2.5.4. Hydrophobicity and Auto-Aggregation Tests
Hydrophobicity: The E. faecium FUA027 bacterial liquid was centrifuged (20 °C, 3000 rpm, 10 min), and the pellet was washed with and suspended in distilled water. The culture suspension was adjusted to an OD600 value of 0.5 ± 0.02 (). Then, an equal volume of xylene solution was added to the bacterial suspension and vortexed for 20 s at 37 °C for 1 h. The absorbance of the supernatant at 600 nm () was determined. Three parallel tests were conducted [30]. The hydrophobic rate was calculated using the following formula:
Auto-aggregation: The FUA027 bacterial liquid was centrifuged (20 °C, 3000 rpm, 10 min) and washed with distilled water. Its OD600 value was adjusted to 0.5 ± 0.02 (). The bacterial suspension was allowed to stand at 37 °C for 4 h, and the absorbance of the supernatant at 600 nm () was determined. Three parallel tests were conducted. The auto-aggregation rate was calculated using the following formula:
2.5.5. Evaluation of Antibacterial Activity
A single colony of E. faecium FUA027 was picked, inoculated into ABB liquid medium, and cultured anaerobically at 37 °C for 24 h. Then, 10 mL of bacterial solution was mixed thoroughly with an equal volume of ethyl acetate extract, vortex shaken for 30 s, and transferred to a separatory funnel. This mixture was allowed to stand at room temperature for 5 min. After the solution was stratified, the upper organic phase was collected and evaporated in a rotary evaporation flask. The rotary evaporator was used to rotary evaporate the organic phase at 60 °C for 10–15 min to ensure the absence of a smell of ethyl acetate. Then, 2 mL ethyl acetate was added to dissolve the residue in the rotary steaming bottle, fully mixed, and filtered through a nylon syringe filter (pore size: 20 μm). The liquid was collected as the antibacterial solution [31]. The experimental group was the upper organic phase of E. faecium FUA027 after extraction with ethyl acetate (concentrated five times) and the lower aqueous phase of E. faecium FUA027 after extraction with ethyl acetate. ABB medium extracts and ethyl acetate were used as blank controls.
The Kirby–Bauer test for antibacterial effects: 100 μL of bacterial solution of Staphylococcus aureus ATCC 12600, Escherichia coli ATCC 25922, Yeast ATCC 24060, and Aspergillus niger ATCC 6273 were evenly applied to the plate, respectively. Then, four sterile filter papers of diameter 5 ± 0.5 mm were placed in each plate. A total of 10 μL of sample was added dropwise to each filter paper sheet and incubated for 12 h at 37 °C. Then, a vernier caliper was used to measure and record the diameter of the suppression ring. The inhibitory effect was evaluated on the basis of the inhibition circle diameter. Three independent tests were repeated [32].
3. Results and Discussion
3.1. Genome Properties
The whole genome sequence of E. faecium FUA027 contained a single, circular 2,718,096-bp-long chromosome with an average GC content of 38.27% (Figure 1). The Glimmer program identified 2700 genes with an estimated coding ratio of 87.1%. Of them, 2617 were protein-coding genes and 83 were RNA genes. Among the 83 RNA genes, 17 genes coded for 5S, 16S, and 23S rRNAs; two genes coded for sRNAs; and 64 genes coded for tRNAs. (Table 1). The Plasmid Finder 2.0 tool did not find any plasmid sequences. The FUA027 genome sequence was submitted to NCBI under the accession number OM670243.
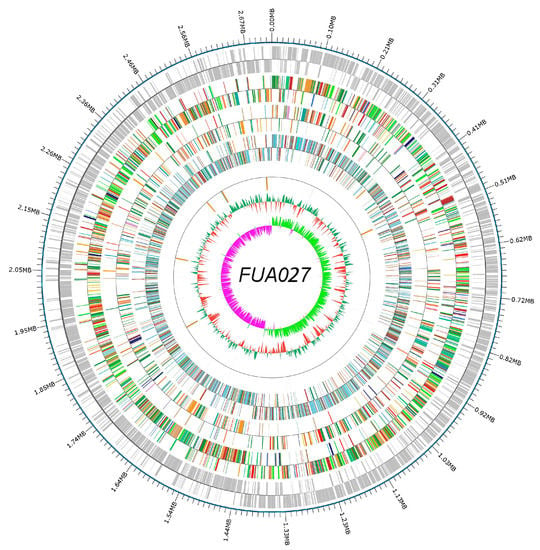
Figure 1.
Whole genome map. The outermost circle is the genome sequence position coordinates, and from outside to center are the coding gene, gene function annotation results (COG, KEGG, GO database annotation results information), ncRNA, and genome GC content: the inward red part indicates that the GC content of the region is lower than the average GC content of the whole genome, and the outward green part is the opposite; and genomic GC skew value: the inward pink part indicates that the GC content of G in the region is lower than that of C; the outward light green part is the opposite.

Table 1.
Characteristics of the E. faecium FUA027 genome.
3.2. Evaluation Safety of E. faecium FUA027
3.2.1. Identification of Antibiotic Resistance Gene
In the clinical setting, probiotic strains resistant to a particular antibiotic are typically associated with infection. Antibiotic resistance genes in the probiotic genome are not in themselves a safety issue, if the genes are not likely transferred to other strains. Instead, probiotics containing these genes could theoretically act as a source of antibiotic resistance genes for potentially pathogenic bacteria. Probiotics must also be tested for the presence of antibiotic resistance genes because studies have confirmed that these genes may be transferred in food and in the intestinal environment.
Enterococcus exhibits stronger natural resistance than other Gram-positive bacteria and acquires resistance genes through various mechanisms to produce multiple high-level drug-resistant strains [33]. Amino acid sequences of E. faecium FUA027 were compared with the drug resistance gene database CARD (https://card.mcmaster.ca/, accessed on 11 December 2022), and protein sequences with >50% similarity in the comparison results were extracted as antibiotic resistance genes. Eighteen antibiotic resistance genes were identified. A predictive analysis of drug resistance genes identified 10 types of aminoglycoside antibiotics, fluoroquinolones, lincosamides, and vancomycin. Probiotic E. faecium strain T-110 and non-pathogenic strain E. faecium NRRL B-2354 both contain a plasmid, according to Natarajan et al. [34]. Importantly, we used the MobileElementFinder tool to search for MGEs. As expected, the absence of MGEs was confirmed. Consequently, because E. faecium FUA027 has no plasmid and none of the antibiotic resistance genes associated with it are located on MGEs, these drug resistance properties cannot be transferred to other pathogenic bacteria through mobile elements, implying no occurrence of drug resistance transmission. Thus, this study from the genetic level confirms that E. faecium FUA027 is safe for the horizontal transfer of drug resistance. To corroborate the results of antibiotic resistance gene analyses, the antibiotic sensitivity test was conducted. Nevertheless, the presence of resistance genes did not exactly match the experimental results observed. According to the results, E. faecium FUA027 was resistant to nine antibiotic types (Table 2). In total, 27 antibiotics were detected. As shown in Table A1, E. faecium FUA027 was resistant to nine types of antibiotics. Combined with the results of the antibiotic susceptibility test in vitro, the antibiotic resistance genes in the genome were analyzed. E. faecium FUA027 was safe in terms of antibiotic resistance.
Table 2.
Putative antibiotic resistance genes identified in the genome of E. faecium FUA027.
3.2.2. Evaluation of Virulence Factor Genes and Toxin-Encoding Genes
According to the gene function classification, virulence genes carried by enterococci mainly encode for proteins related to adherence, exotoxin, exoenzyme, immunomodulation, and biofilm [35]. The VFDB was used to identify virulence factor genes in E. faecium FUA027; however, most putative virulence factor genes had <60% similarity with VFDB [36]. In total, seven potential virulence factor genes were identified (Table 3). These genes may encode for proteins involved in adhesion, immunomodulation, exoenzyme, and biofilm. Genes encoding enterococcal hemolysin A (hlyA), cytolysin (cyl), aggregation substance (as), enterococcal surface protein, sex pheromones (cob and ccf), and serum resistance-associated gene (sra), which are well-known potential virulence factors, were missing in E. faecium FUA027. According to Deng’s study, among 110 probiotic Enterococcus spp. 35 (31.8%) enterococcal strains exhibited β-hemolytic characteristics. However, in our study, FUA027 exhibited γ-hemolysis on blood plates and no genes encoding Hbl, Nhe, or cytotoxin K, which are associated with hemolysis and toxin production, were found in the genome (Figure 2). These results thus confirmed that E. faecium FUA027 would be used in the preparation of UA by industrial fermentation.
Table 3.
Putative virulence factors encoded in the E. faecium FUA027 genome.
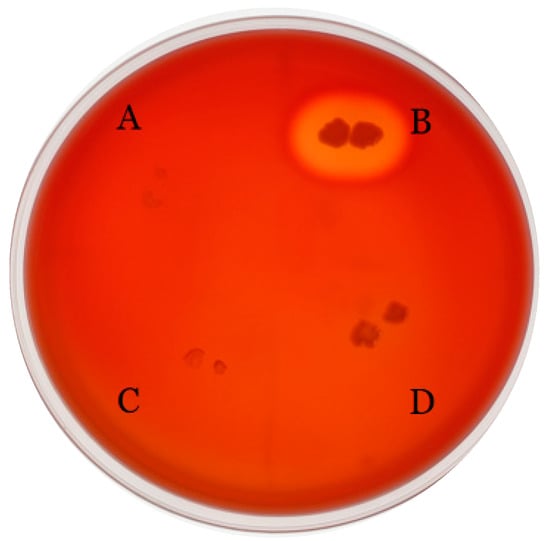
Figure 2.
Blood agar plate test results. S. thermophilus (A), S. aureus (B), S. thermophilus FUA329, screening from human milk sample (C), and E. faecium FUA027 (D).
3.2.3. Biogenic Amine Production
The results of nitrate reductase activity revealed that E. faecium FUA027 did not contain nitrate reductase. No color change was observed in the tubes containing the test strains, and the color was red after the addition of trace zinc powder, indicating that the test group was negative. The tube containing the quality control strain E. coli ATCC 25922 was red and positive.
The amino acid decarboxylase activity of E. faecium FUA027 was preliminarily detected on the basis of the color change in the amino acid decarboxylase medium. With E. faecium FUA027, the color of the amino acid decarboxylase medium remained unchanged and yellow, indicating that no biogenic amines (BA) were produced in the medium by the strain. The experimental results revealed that FUA027 did not possess lysine, ornithine, arginine, tryptophan, or phenylalanine decarboxylase activities. The main source of BA in food is the microbial decarboxylation of amino acids. For example, the decarboxylation of tyrosine, ornithine, and lysine produces tyramine, putrescine, and cadaverine, respectively. BA accumulation in food has serious implications for food safety and human health [37]. Of the 129 enterococci strains of three different origins (food, veterinary, and human) screened by Sarantinopoulos et al., none produced histamine, cadaverine, or putrescine [38]. However, >90% of E. faecium strains isolated from cheese have been identified as tyramine producers. Some E. faecium strains from humans also produced putrescine [39]. E. faecium FUA027 was found to not produce BA, and thus we believe that this strain may be used safely in industrial fermentation.
3.3. Assessment of Probiotic Properties
3.3.1. Acid and Bile Salt Tolerance In Vitro
Normal human gastric juice pH is approximately 1–3, and normal human intestinal pH is approximately 6.8–7.0. The pH in the stomach can rise to 4–5 after food is consumed. Probiotics can only exert their probiotic role if they resist the inhibitory effects of gastric acid and pepsin on the intestine [40]. A gene encoding conjugated bile acid hydrolase (cbh) and three genes encoding bile acid sodium symporter family proteins were discovered in E. faecium FUA027; these genes may have contributed to bile salt resistance. F0F1-ATPase is considered the main pH regulator inside cells. Eight genes coding for the F0F1-ATP synthase subunit were identified in the FUA027 genome. Furthermore, a cation transporter gene, two (Na+/H+) antiporter genes, and a sodium ion transporter gene linked to pH regulation and ion homeostasis were discovered (Table A2). The survival rates of E. faecium FUA027 in the in vitro acid tolerance test at different pH values are shown in Figure 3A. The survival rate declined steadily as the pH value decreased. Studies have shown that strains with a survival rate of >60% are acid-resistant strains. The survival rate of E. faecium FUA027 in the in vitro acid tolerance test at pH 3.0 was >60% and that at pH 2.0 was >50%. Compared to acid-tolerant strains, E. faecium FUA027 was less acid-tolerant.
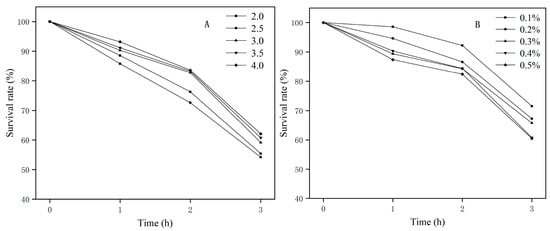
Figure 3.
(A) Acid resistance test results; (B) bile salt resistance test results.
Another crucial sign for assessing the qualities of possible probiotics is the tolerance of strains to high bile salt concentrations in the human gastrointestine. Studies have shown that the small intestine contains approximately 0.3% of bile salts. In our study, the survival rate of the strain was higher than 67% at bile salt concentrations of 0.1%–0.3%. The strain survival rate was still >60.00% at bile salt concentrations of 0.4% and 0.5% (Figure 3B), which indicates that the strain has excellent bile salt resistance.
We identified a gene coding for conjugated bile acid hydrolase (cbh), two conjugated bile acid hydrolase genes (namely nhaC and napA), and ABC transporter genes potentially contributing to bile salt resistance in E. faecium FUA027. Eight genes coding for the F0F1-ATP synthase subunit (namely atpB, atpE, atpF, atpH, atpA, atpG, atpD, and atpC) were identified in the FUA027 genome. Therefore, we suggest that the in vitro results of acid and bile salt tolerance in E. faecium FUA027 are explained by these related genes in its genome.
3.3.2. Antioxidant Ability In Vitro
Some probiotic metabolites can lessen the oxidative damage that causes aging and chronic diseases [41]. The results of the in vitro antioxidant ability of E. faecium FUA027 are presented in Table 4. The DPPH scavenging activity of the fermentation supernatant was as high as 57.62%, the superoxide anion scavenging capacity was 36.23%, and the clearance rate of hydroxyl radical was 30.12%. Polysaccharides, phosphonic acid, and peptidase, which are fundamental cell wall building blocks, are crucial for antioxidation. The extracellular metabolite structure is closely related to the antioxidant activity of the fermentation supernatant. In addition, the antioxidant activities of L. plantarum and E. faecalis were studied. The DPPH scavenging activity of L. plantarum was 62.78%, which was close to that of E. faecium FUA027. By contrast, the activity of E. faecalis was lower than that of E. faecium FUA027.
Table 4.
Antioxidant activity of E. faecium FUA027 in vitro.
Ten genes associated with the oxidative stress response were found in the FUA027 genome; these genes could help the strain avoid damage by O2− and H2O2−, such as peroxide-responsive repressor (perR), NADH peroxidase (npr), alkyl hydroperoxide reductase (ahpC/F), glutathione peroxidase (gpx), superoxide dismutase (sodA), thioredoxin reductase (trxB), and glutathione reductase (gor). Among them, perR regulates H2O2− induced oxidative stress. In the presence of H2O2− or with iron and manganese ion deficiencies, perR upregulates antioxidant enzymes such as catA and ahpC/F to scavenge H2O2− and alkyl hydroperoxides (Table A2). The presence of these antioxidant genes indicated that E. faecium FUA027 has high antioxidant activity. Based on the results of genomic and phenotypic experimental analyses, we speculate that this may be due to the expression of antioxidant genes in the E. faecium FUA027 genome, such as catalase, glutathione peroxidase, and superoxide dismutation, which make FUA027 possess a good antioxidant capacity.
3.3.3. Evaluation of Adhesion-Related Genes
Probiotics play a beneficial role by adhering to intestinal mucosa and epithelial cells. We searched for gene annotation data related to adhesion, colonization, mucin binding, flagella hook, and fibrinogen/fibronectin binding. Adhesion lipoprotein, s-ribosylhomocysteine lyasef (luxS), segregation and condensation protein B (scpB) were found in the E. faecium FUA027 genome (Table 5) [42].
Table 5.
Cell adhesion-associated proteins in the E. faecium FUA027 genome.
Biofilms of lactic acid bacteria can colonize the intestine, thereby protecting strains in gastrointestinal transit, producing certain antimicrobial compounds, and stimulating the immune response. Auto-aggregation is a crucial property of biofilm formation, and hydrophobicity may assist in adhesion. Auto-aggregation and hydrophobicity are vital indicators of the ability of microbes to respond to bacterial gut colonization. FUA027 exhibited higher hydrophobicity and auto-aggregation than the commercial probiotic strain Bifidobacterium longum BB536). This demonstrates that E. faecium FUA027 can better colonize the intestinal tract, and thus exert its probiotic properties.
3.3.4. Antibacterial Test of E. faecium FUA027 against Quality Control Strains
In the in vitro experiment, the inhibitory ability of E. faecium FUA027 against four test strains was investigated. As shown in Figure 4, FUA027 exhibited significant inhibitory effects on E. coli ATCC 25922 and S. aureus ATCC 12600, with inhibition circle sizes of 26.24 ± 0.34 mm and 22.12 ± 0.26 mm, respectively. The inhibition circle sizes were 9.2 ± 0.52 mm and 8.74 ± 0.38 mm for Yeast ATCC 24060 and A. niger ATCC 6273, respectively. E. faecium FUA027 had a significantly better inhibitory effect on bacteria than on fungi. Antimicrobial activity is a crucial property of probiotics against gastrointestinal infections. E. faecium mainly exerts its bacteriostatic effect by secreting organic acids. Furthermore, bacteriocins, bacteriocin-like, and hydrogen peroxide secreted by E. faecium can inhibit intestinal pathogenic microorganisms to some extent. Many bacteriocin-producing E. faecalis strains have been reported. Rahmeh et al. explored how E. faecium S6 exerts its antimicrobial effect by producing enterotoxins and organic acids [43]. Valenzuela et al. isolated an E. faecium PE 2-2 strain from seafood that inhibited S. aureus and demonstrated that this strain carried the enterocin A structural gene [44]. Basanta et al. reported that E. faecium L50 isolated from a Spanish dry fermented sausage produces enterocin L50 (EntL50, EntL50A, and EntL50B), enterocin P, and enterocin Q and exhibits a broad antimicrobial spectrum [45]. Enterococins are often used as a preservative for meat and dairy products. The most widely used enterococins are enterocin A and enterocin B, belonging to class II bacteriocin. In our study, four biosynthetic gene clusters associated with T3PKS, a cyclic lactone autoinducer, were identified using AntiSMASH 5.0, and BAGEL 4.0 predicted a bacteriocin from the class sactipeptide in the E. faecium FUA027 genome. Sactipeptides (sulfur-to-alpha carbon thioether cross-linked peptides) are ribosomally synthesized and post-translationally modified peptides that exhibit antibacterial activity [46]. In conclusion, in vitro experiments supported the presence and activity of extracellularly secreted bacteriocins, as they significantly inhibit the growth of E. coli ATCC 25922 and S. aureus ATCC 12600.
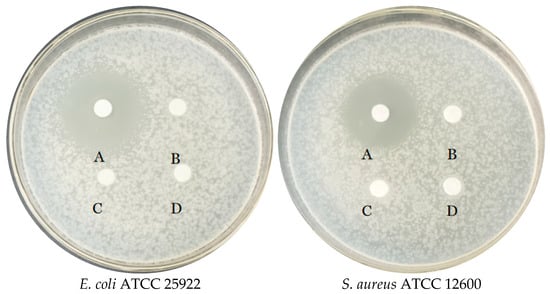

Figure 4.
Antimicrobial activity of E. faecium FUA027 against quality control strains. The upper organic phase of E. faecium FUA027 fermentation liquid after extraction with ethyl acetate (concentrated five times) (A), the lower aqueous phase of E. faecium FUA027 after extraction with ethyl acetate (B), ABB medium extracts (C), and ethyl acetate (D).
4. Conclusions
In summary, we here described the whole-genome sequence of E. faecium FUA027. FUA027 has a 2,718,096-bp-long chromosome with an average GC content of 38.27%. Genomic screening revealed that FUA027 lacked key virulence factor genes and toxin-coding genes. Although 18 antibiotic resistance genes were screened from the strain, the strain has no plasmids or mobile elements and is therefore unlikely to undergo the acquisition and transfer of resistance genes. The safety of this strain was further confirmed through hemolysis tests, metabolic toxicity tests, and antibiotic resistance tests. The detection of antimicrobial gene clusters and adhesion- and stress-associated genes in the genome, along with the results of tolerance tests such as tolerance to acid and bile salt and in vitro antioxidant activity-related genes, revealed the probiotic properties of the strain. Genomic analysis combined with phenotypic studies confirmed the safety and probiotic properties of this strain as a potential probiotic candidate.
Author Contributions
Conceptualization, S.L. and Y.F.; methodology, S.L.; software, M.X. and W.Z.; validation, S.L. and G.Y.; formal analysis, S.M., F.H. and Y.H.; investigation, X.H.; resources, S.L.; data curation, X.Z. and Y.Z.; writing—original draft preparation, M.X.; writing—review and editing, S.L. and Y.F.; visualization, J.S.; supervision, S.L.; project administration, S.L.; funding acquisition, S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Priority Academic Program Development of Jiangsu Higher Education Institutions, the Key Natural Science Foundation of the Jiangsu Higher Education Institutions of China (20KJA550001), the National Natural Science Foundation of Jiangsu Ocean University (KQ20041), Project “333” of JiangsuProvince, Open-end Funds of Jiangsu Key Laboratory of Marine Bioresources and Environment (SH20191204), Jangsu Province College Students’ Innovation and Entrepreneurship Training Program (SZ202111641638001).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
E. faecium FUA027 antibiotic resistance judgment criteria and test results.
Table A1.
E. faecium FUA027 antibiotic resistance judgment criteria and test results.
| Antibiotic | Standard for Judging Diameter of Inhibition Zone Diam (mm) | Drug Contents (µg) | Zone Diam (mm) | Antibacterial Effect | ||
|---|---|---|---|---|---|---|
| R | I | S | ||||
| Amikacin | ≤14 | 15~16 | ≥17 | 30 | 10 ± 0.2 | R |
| Norfloxacin | ≤12 | 13~16 | ≥17 | 10 | 19 ± 0.3 | S |
| Ofloxacin | ≤12 | 13~15 | ≥16 | 5 | 18 ± 0.4 | S |
| Ciprofloxacin | ≤15 | 16~20 | ≥21 | 5 | 15 ± 0.1 | I |
| Levofloxacin | ≤12 | 13~16 | ≥17 | 5 | 25 ± 0.3 | S |
| Erythromycin | ≤13 | 14~22 | ≥23 | 15 | 22 ± 0.1 | I |
| Tetracycline | ≤14 | 15~18 | ≥19 | 30 | 21 ± 0.2 | S |
| Cefuroxime | ≤14 | 15~17 | ≥18 | 30 | 30 ± 0.2 | S |
| Cefazolin | ≤14 | - | ≥15 | 30 | 22 ± 0.3 | S |
| Cephalothin | ≤14 | 15~17 | ≥18 | 30 | 27 ± 0.4 | S |
| Cefotaxime | ≤22 | 23~25 | ≥26 | 30 | 30 ± 0.1 | S |
| Cefatriaxone | ≤13 | 14~20 | ≥21 | 30 | 24 ± 0.4 | S |
| Ceftazidime | ≤14 | 15~17 | ≥18 | 30 | 18 ± 0.3 | S |
| Piperacillin | ≤28 | - | ≥29 | 100 | 21 ± 0.3 | S |
| Ampicillin | ≤16 | 18~24 | ≥25 | 10 | 22 ± 0.2 | S |
| Oxacillin | ≤17 | - | ≥25 | 1 | 14 ± 0.4 | R |
| Penicillin G | ≤28 | - | ≥29 | 10 | 22 ± 0.6 | S |
| Aztreonam | ≤15 | 16~21 | ≥22 | 30 | 0 | R |
| Compound sulfamethoxazole | ≤10 | 11~15 | ≥16 | 23.75 | 0 | R |
| Nitrofurantoin | ≤14 | 15~16 | ≥17 | 300 | 15 ± 0.6 | I |
| Chloramphenicol | ≤12 | 13~17 | ≥18 | 30 | 19 ± 0.3 | S |
| Polymyxin B | ≤11 | 12~14 | ≥15 | 300 | 0 | R |
| Clindamycin | ≤13 | 14~17 | ≥18 | 2 | 8 ± 0.4 | R |
| Kanamycin | ≤12 | 13~14 | ≥15 | 30 | 6 ± 0.4 | R |
| Gentamicin | ≤12 | 13~14 | ≥15 | 10 | 9 ± 0.6 | R |
| Streptomycin | ≤11 | 12~14 | ≥15 | 10 | 7 ± 0.7 | R |
| Vancomycin | ≤14 | 15~16 | ≥17 | 30 | 19 ± 0.3 | S |
Table A2.
Stress-responsive proteins in the whole genome of E. faecium FUA027.
Table A2.
Stress-responsive proteins in the whole genome of E. faecium FUA027.
| Type of Stress Response | Protein | Gene Locus |
|---|---|---|
| Acid stress response | F0F1-ATPase | GM_000855, GM_000856 |
| GM_000857, GM_000858 | ||
| GM_000859, GM_000860 | ||
| GM_000861, GM_000862 | ||
| Na+/H+ antiporter | GM_000704, GM_001361 | |
| Bile salts stress response | Conjugated bile acid hydrolase | GM_001069 |
| ABC transporter | GM_002271, GM_002452 | |
| GM_000767, GM_002209 | ||
| GM_002135, GM_002210 | ||
| GM_002486, GM_002485 | ||
| GM_002270, GM_000917 | ||
| GM_000916, GM_002136 | ||
| GM_001223, GM_001224 | ||
| Temperature stress response | Cold shock protein | GM_001318 |
| Metal stress response | Divalent metal cation transporter | GM_000913 |
| Cadmium, zinc and cobalt-transporting ATPase | GM_000987 | |
| Potassium/sodium uptake protein | GM_000822 | |
| Oxidative stress response | Alkyl hydroperoxide reductase | GM_001365, GM_001366 |
| Glutathione peroxidase | GM_000501 | |
| Thioredoxin reductases | GM_000926 | |
| Glutathione reductase | GM_002670 | |
| Superoxide dismutase | GM_001908 | |
| Peroxide-responsive repressor | GM_001511 | |
| NADH peroxidase | GM_000518 | |
| Superoxide dismutase | GM_001908 |
References
- García-Villalba, R.; Beltrán, D.; Espín, J.C.; Selma, M.V.; Tomás-Barberán, F.A. Time Course Production of Urolithins from Ellagic Acid by Human Gut Microbiota. J. Agric. Food Chem. 2013, 61, 8797–8806. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.; Green, A.E.; Kim, Y.A.; Bae, S.-J.; Ha, K.-T.; Gariani, K.; Lee, M.-R.; Menzies, K.J.; Ryu, D. Sarcopenia and Muscle Aging: A Brief Overview. Endocrinol. Metab. 2020, 35, 716–732. [Google Scholar] [CrossRef] [PubMed]
- Selma, M.V.; Romo-Vaquero, M.; García-Villalba, R.; González-Sarrías, A.; Tomás-Barberán, F.A.; Espín, J.C. The human gut microbial ecology associated with overweight and obesity determines ellagic acid metabolism. Food Funct. 2016, 7, 1769–1774. [Google Scholar] [CrossRef]
- Alfei, S.; Marengo, B.; Zuccari, G. Oxidative Stress, Antioxidant Capabilities, and Bioavailability: Ellagic Acid or Urolithins? Antioxidants 2020, 9, 707. [Google Scholar] [CrossRef] [PubMed]
- Al-Harbi, S.A.; Abdulrahman, A.O.; Zamzami, M.A.; Khan, M.I. Urolithins: The Gut Based Polyphenol Metabolites of Ellagitannins in Cancer Prevention, a Review. Front. Nutr. 2021, 8, 647582. [Google Scholar] [CrossRef]
- García-Villalba, R.; Giménez-Bastida, J.A.; Cortés-Martín, A.; Ávila-Gálvez, M.; Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C.; González-Sarrías, A. Urolithins: A Comprehensive Update on their Metabolism, Bioactivity, and Associated Gut Microbiota. Mol. Nutr. Food Res. 2022, 66, 2101019. [Google Scholar] [CrossRef] [PubMed]
- Xian, W.; Yang, S.; Deng, Y.; Yang, Y.; Chen, C.; Li, W.; Yang, R. Distribution of Urolithins Metabotypes in Healthy Chinese Youth: Difference in Gut Microbiota and Predicted Metabolic Pathways. J. Agric. Food Chem. 2021, 69, 13055–13065. [Google Scholar] [CrossRef]
- D’Amico, D.; Andreux, P.A.; Valdés, P.; Singh, A.; Rinsch, C.; Auwerx, J. Impact of the Natural Compound Urolithin A on Health, Disease, and Aging. Trends Mol. Med. 2021, 27, 687–699. [Google Scholar] [CrossRef]
- Cortés-Martín, A.; García-Villalba, R.; González-Sarrías, A.; Romo-Vaquero, M.; Loria-Kohen, V.; Ramírez-De-Molina, A.; Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. The gut microbiota urolithin metabotypes revisited: The human metabolism of ellagic acid is mainly determined by aging. Food Funct. 2018, 9, 4100–4106. [Google Scholar] [CrossRef]
- Kang, I.; Buckner, T.; Shay, N.F.; Gu, L.; Chung, S. Improvements in Metabolic Health with Consumption of Ellagic Acid and Subsequent Conversion into Urolithins: Evidence and Mechanisms. Adv. Nutr. 2016, 7, 961–972. [Google Scholar] [CrossRef]
- Gaya, P.; Peirotén, Á; Medina, M.; Alvaréz, I.; Landete, J.M. Bifidobacterium pseudocatenulatum INIA P815: The first bacterium able to produce urolithins a and b from ellagic acid. J. Funct. Foods 2018, 45, 95–99. [Google Scholar]
- Liu, Q.; Liu, S.; Ye, Q.; Hou, X.; Yang, G.; Lu, J.; Hai, Y.; Shen, J.; Fang, Y. A Novel Streptococcus thermophilus FUA329 Isolated from Human Breast Milk Capable of Producing Urolithin A from Ellagic Acid. Foods 2022, 11, 3280. [Google Scholar] [CrossRef]
- Mi, H.; Liu, S.; Hai, Y.; Yang, G.; Lu, J.; He, F.; Zhao, Y.; Xia, M.; Hou, X.; Fang, Y. Lactococcus garvieae FUA009, a Novel Intestinal Bacterium Capable of Producing the Bioactive Metabolite Urolithin A from Ellagic Acid. Foods 2022, 11, 2621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Fang, Y.; Yang, G.; Hou, X.; Hai, Y.; Xia, M.; He, F.; Zhao, Y.; Liu, S. Isolation and characterization of a novel human intestinal Enterococcus faecium FUA027 capable of producing urolithin A from ellagic acid. Front. Nutr. 2022, 9, 1039697. [Google Scholar] [CrossRef] [PubMed]
- Koirala, S.; Anal, A.K. Probiotics-based foods and beverages as future foods and their overall safety and regulatory claims. Future Foods 2021, 3, 100013. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, K.; Nie, K.; Deng, M.; Luo, W.; Wu, X.; Huang, Y.; Wang, X. Assessment of the safety and probiotic properties of Roseburia intestinalis: A potential “Next Generation Probiotic”. Front. Microbiol. 2022, 13, 973046. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Hombach, M.; Maurer, F.; Pfiffner, T.; Böttger, E.C.; Furrer, R. Standardization of Operator-Dependent Variables Affecting Precision and Accuracy of the Disk Diffusion Method for Antibiotic Susceptibility Testing. J. Clin. Microbiol. 2015, 53, 3864–3869. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Choi, S.I.; Jeong, Y.; Kang, C.H. Evaluation of Safety and Probiotic Potential of Enterococcus faecalis MG5206 and Enterococcus faecium MG5232 Isolated from Kimchi, a Korean Fermented Cabbage. Microorganisms 2022, 10, 2070. [Google Scholar] [CrossRef] [PubMed]
- De Aguero, N.L.; Frizzo, L.S.; Ouwehand, A.C.; Aleu, G.; Rosmini, M.R. Technological Characterisation of Probiotic Lactic Acid Bacteria as Starter Cultures for Dry Fermented Sausages. Foods 2020, 9, 596. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-P.; Liu, D.-M.; Zhao, S.; Huang, Y.-Y.; Yu, J.-J.; Zhou, Q.-Y. Assessing the safety and probiotic characteristics of Bacillus coagulans 13002 based on complete genome and phenotype analysis. LWT 2022, 155, 112847. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed]
- Pieniz, S.; Andreazza, R.; Anghinoni, T.; Camargo, F.; Brandelli, A. Probiotic potential, antimicrobial and antioxidant activities of Enterococcus durans strain LAB18s. Food Control 2014, 37, 251–256. [Google Scholar] [CrossRef]
- Wu, Y.; Li, S.; Tao, Y.; Li, D.; Han, Y.; Show, P.L.; Wen, G.; Zhou, J. Fermentation of blueberry and blackberry juices using Lactobacillus plantarum, Streptococcus thermophilus and Bifidobacterium bifidum: Growth of probiotics, metabolism of phenolics, antioxidant capacity in vitro and sensory evaluation. Food Chem. 2021, 348, 129083. [Google Scholar] [CrossRef]
- Mu, G.; Gao, Y.; Tuo, Y.; Li, H.; Zhang, Y.; Qian, F.; Jiang, S. Assessing and comparing antioxidant activities of lactobacilli strains by using different chemical and cellular antioxidant methods. J. Dairy Sci. 2018, 101, 10792–10806. [Google Scholar] [CrossRef]
- Azat, R.; Liu, Y.; Li, W.; Kayir, A.; Lin, D.-B.; Zhou, W.-W.; Zheng, X.-D. Probiotic properties of lactic acid bacteria isolated from traditionally fermented Xinjiang cheese. J. Zhejiang Univ. Sci. B 2016, 17, 597–609. [Google Scholar] [CrossRef]
- Maragkoudakis, P.A.; Zoumpopoulou, G.; Miaris, C.; Kalantzopoulos, G.; Pot, B.; Tsakalidou, E. Probiotic potential of Lactobacillus strains isolated from dairy products. Int. Dairy J. 2006, 16, 189–199. [Google Scholar] [CrossRef]
- Humphries, R.; Bobenchik, A.M.; Hindler, J.A.; Schuetz, A.N. Overview of Changes to the Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Susceptibility Testing, M100, 31st Edition. J. Clin. Microbiol. 2021, 59, e00213-21. [Google Scholar] [CrossRef] [PubMed]
- Klare, I.; Konstabel, C.; Badstubner, D.; Werner, G.; Witte, W. Occurrence and spread of antibiotic resistances in Enterococcus faecium. Int. J. Food Microbiol. 2003, 88, 269–290. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, P.; Parani, M. First Complete Genome Sequence of a Probiotic Enterococcus faecium Strain T-110 and Its Comparative Genome Analysis with Pathogenic and Non-pathogenic Enterococcus faecium Genomes. J. Genet. Genom. 2015, 42, 43–46. [Google Scholar] [CrossRef]
- Graham, K.; Stack, H.; Rea, R. Safety, beneficial and technological properties of enterococci for use in functional food applications—A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 3836–3861. [Google Scholar] [CrossRef] [PubMed]
- Terai, T.; Kato, K.; Ishikawa, E.; Nakao, M.; Ito, M.; Miyazaki, K.; Kushiro, A.; Imai, S.; Nomura, Y.; Hanada, N.; et al. Safety assessment of the candidate oral probiotic Lactobacillus crispatus YIT 12319: Analysis of antibiotic resistance and virulence-associated genes. Food Chem. Toxicol. 2020, 140, 111278. [Google Scholar] [CrossRef]
- Linares, D.M.; Martín, M.; Ladero, V.; Alvarez, M.A.; Fernández, M. Biogenic Amines in Dairy Products. Crit. Rev. Food Sci. Nutr. 2011, 51, 691–703. [Google Scholar] [CrossRef]
- Sarantinopoulos, P.; Andrighetto, C.; Georgalaki, M.D.; Rea, M.; Lombardi, A.; Cogan, T.M.; Kalantzopoulos, G.; Tsakalidou, E. Biochemical properties of enterococci relevant to their technological performance. Int. Dairy J. 2001, 11, 621–647. [Google Scholar] [CrossRef]
- Ladero, V.; Fernandez, M.; Calles-Enriquez, M.; Sanchez-Llana, E.; Canedo, E.; Martin, M.C.; Alvarez, M.A. Is the production of the biogenic amines tyramine and putrescine a species-level trait in enterococci? Food Microbiol. 2012, 30, 132–138. [Google Scholar] [CrossRef]
- Wang, J.; Da, R.; Tuo, X.; Cheng, Y.; Wei, J.; Jiang, K.; Lv, J.; Adediji, O.M.; Han, B. Probiotic and Safety Properties Screening of Enterococcus faecalis from Healthy Chinese Infants. Probiotics Antimicrob. Proteins 2020, 12, 1115–1125. [Google Scholar] [CrossRef]
- Succi, M.; Tremonte, P.; Reale, A.; Sorrentino, E.; Grazia, L.; Pacifico, S.; Coppola, R. Bile salt and acid tolerance of Lactobacillus rhamnosus strains isolated from Parmigiano Reggiano cheese. FEMS Microbiol. Lett. 2005, 244, 129–137. [Google Scholar] [CrossRef]
- Zago, M.; Fornasari, M.E.; Carminati, D.; Burns, P.; Suàrez, V.; Vinderola, G.; Reinheimer, J.; Giraffa, G. Characterization and probiotic potential of Lactobacillus plantarum strains isolated from cheeses. Food Microbiol. 2011, 28, 1033–1040. [Google Scholar] [CrossRef]
- Rahmeh, R.; Akbar, A.; Alonaizi, T.; Kishk, M.; Shajan, A.; Akbar, B. Characterization and application of antimicrobials produced by Enterococcus faecium S6 isolated from raw camel milk. J. Dairy Sci. 2020, 103, 11106–11115. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, A.S.; Benomar, N.; Abriouel, H.; Cañamero, M.M.; Gálvez, A. Isolation and identification of Enterococcus faecium from seafoods: Antimicrobial resistance and production of bacteriocin-like substances. Food Microbiol. 2010, 27, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Basanta, A.; Sánchez, J.; Gómez-Sala, B.; Herranz, C.; Hernández, P.E.; Cintas, L.M. Antimicrobial activity of Enterococcus faecium L50, a strain producing enterocins L50 (L50A and L50B), P and Q, against beer-spoilage lactic acid bacteria in broth, wort (hopped and unhopped), and alcoholic and non-alcoholic lager beers. Int. J. Food Microbiol. 2008, 125, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; van der Donk, W. Macrocyclization and Backbone Modification in RiPP Biosynthesis. Annu. Rev. Biochem. 2022, 91, 269–294. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).