
Comparative Genomics Analysis of Habitat Adaptation by Lactobacillus kefiranofaciens
Abstract
:1. Introduction
2. Material and Methods
2.1. Strains
2.2. DNA Extraction
2.3. DNA Quality Control and Genome Resequencing
2.4. Genome Splicing Assembly
2.5. Comparative Genomics Analysis
2.5.1. Average Nucleotide Identity (ANI) Analyses
2.5.2. Construction of Pan-Core Gene Sets
2.5.3. Construction of Phylogenetic Trees
2.5.4. Functional Gene Annotation
2.5.5. Carbohydrate Active Enzyme Analysis
2.5.6. Mapping and Data Analysis
3. Results
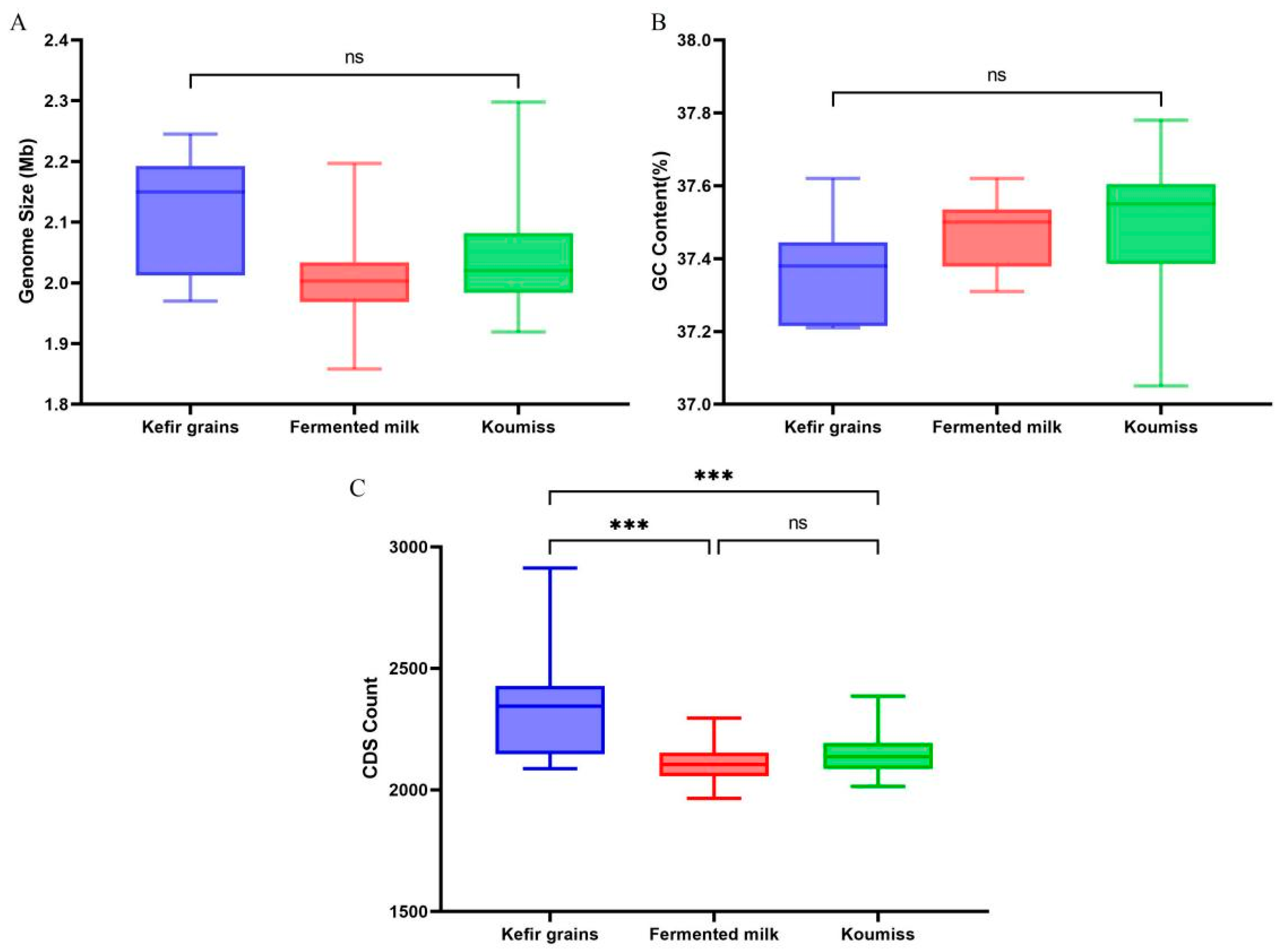
3.1. General Genomic Characteristics of L. kefiranofaciens
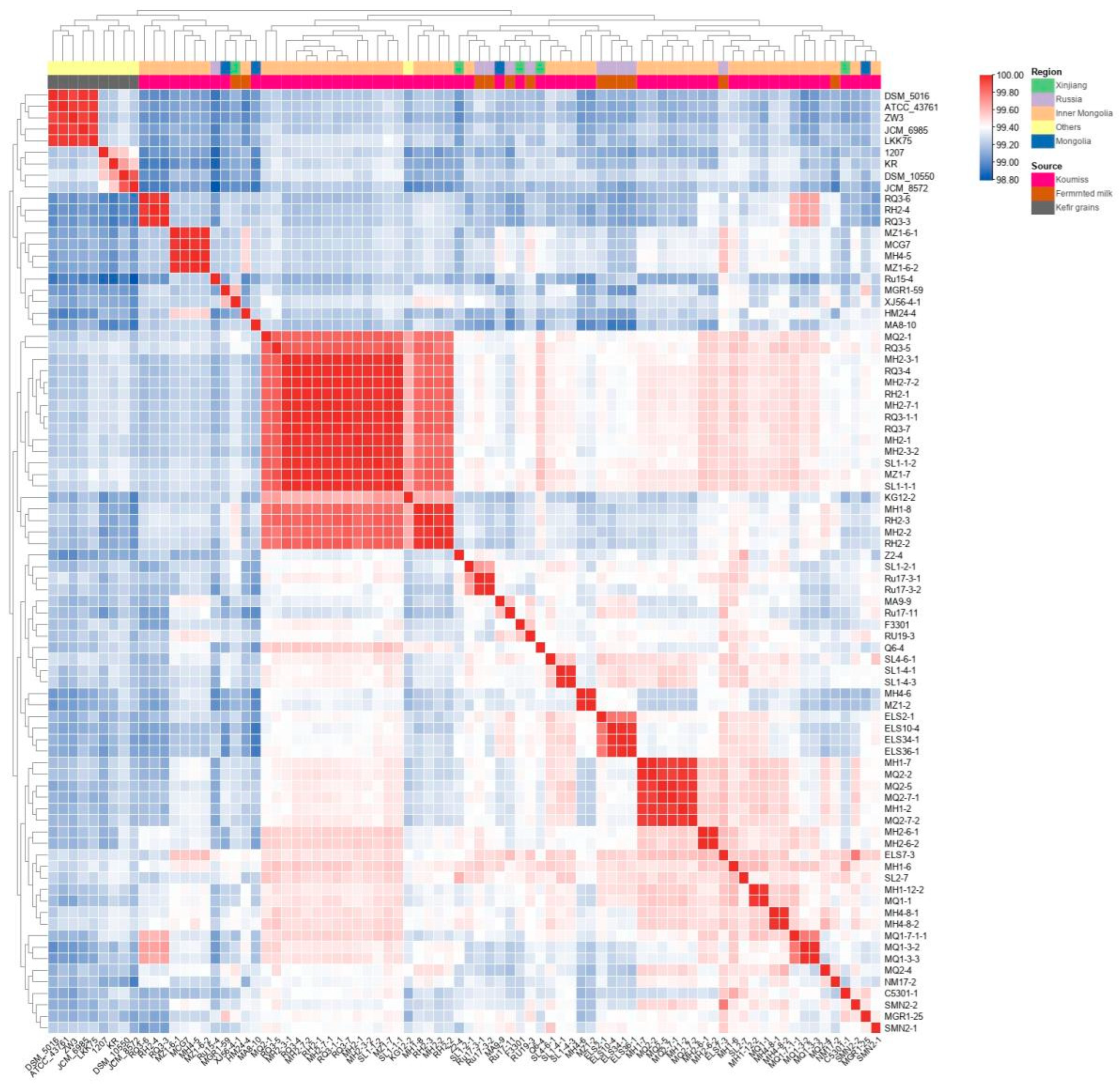
3.2. Average Nucleotide Identity (ANI) Analysis
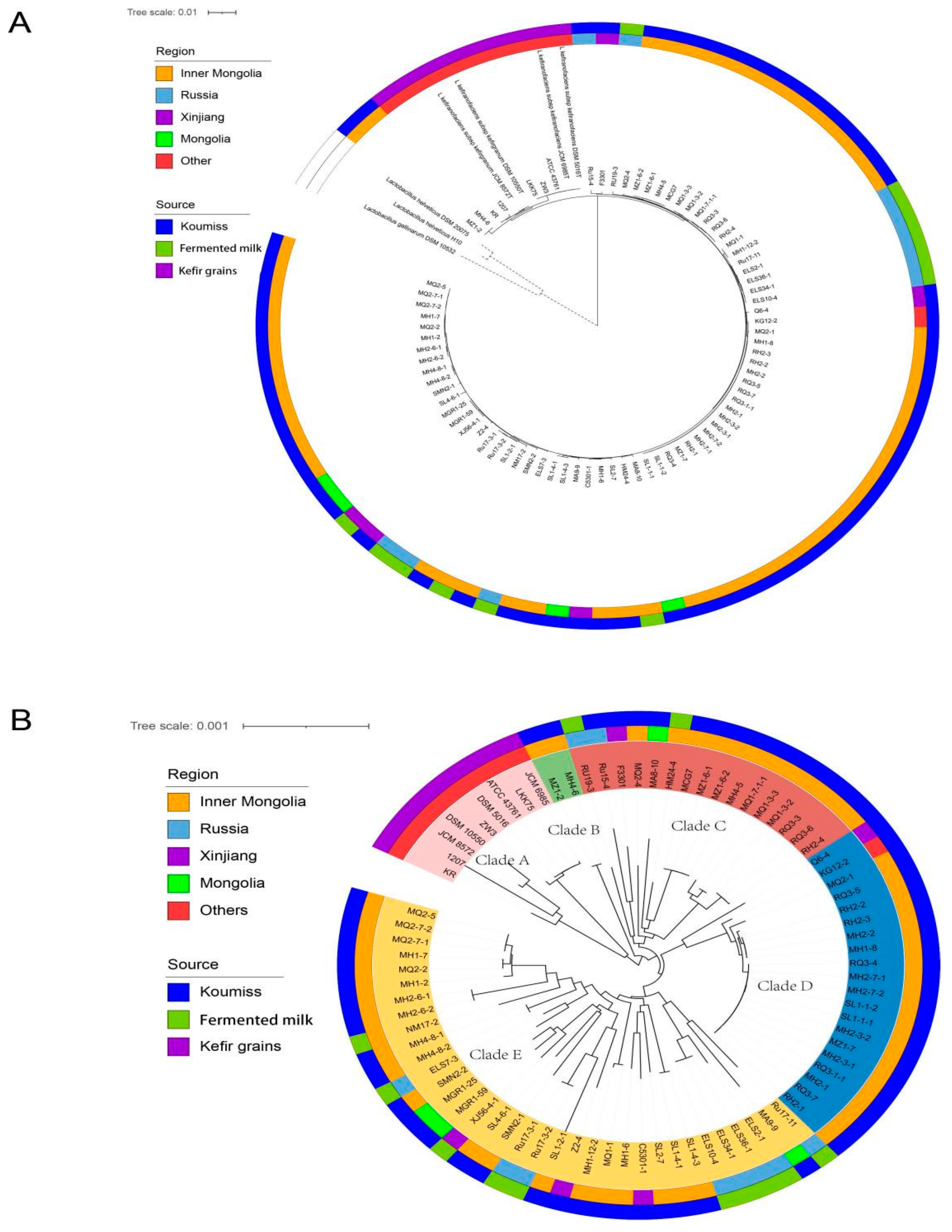
3.3. Phylogenetic Tree Construction Based on Core Genes
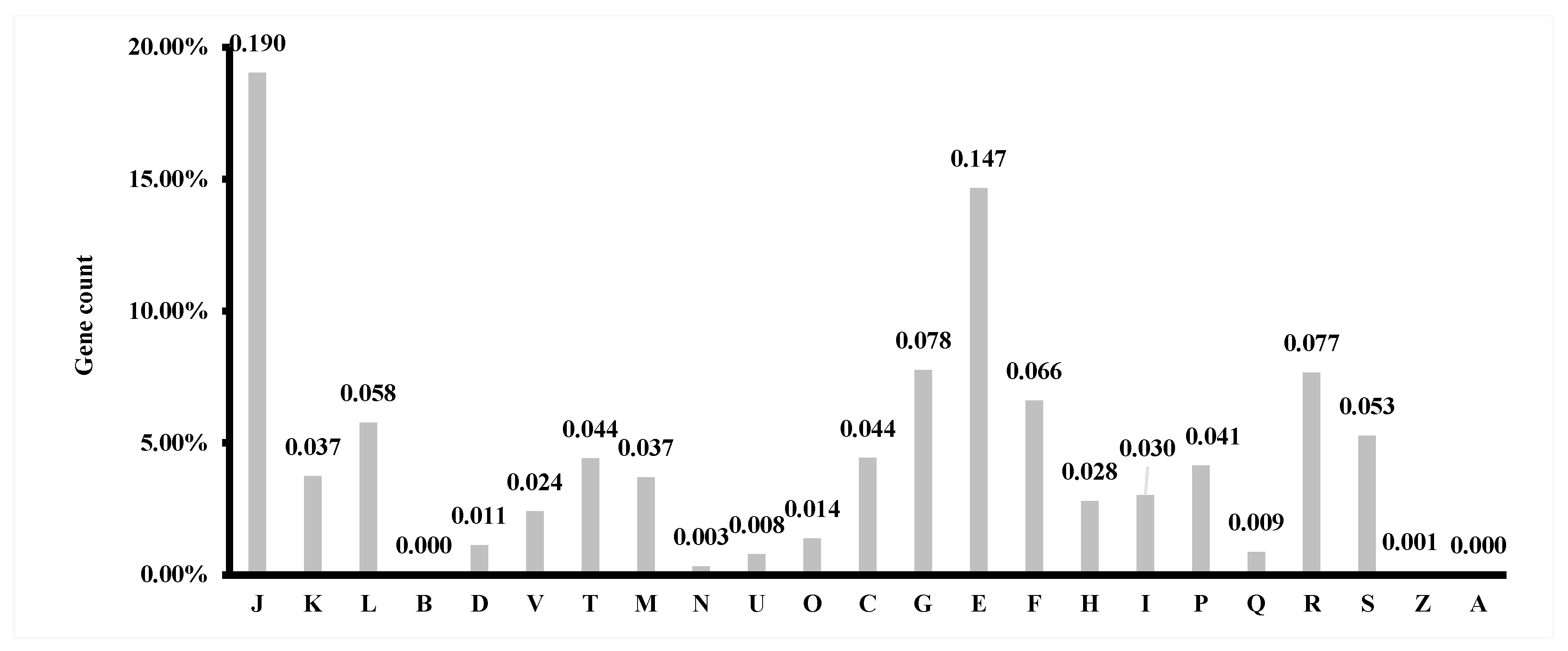
3.4. Gene Prediction and Annotation
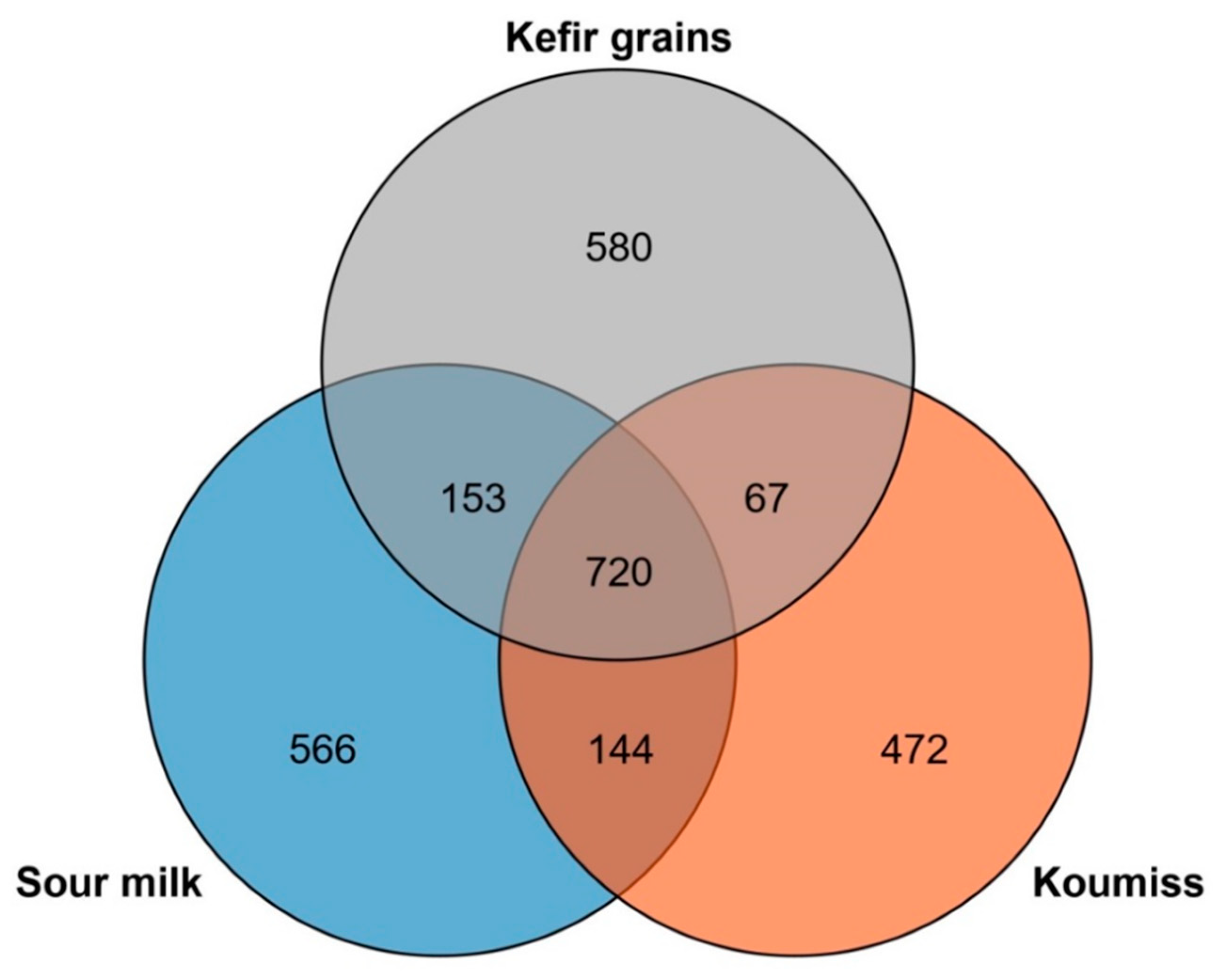
3.5. Core Genome and Pan-Genome Analysis
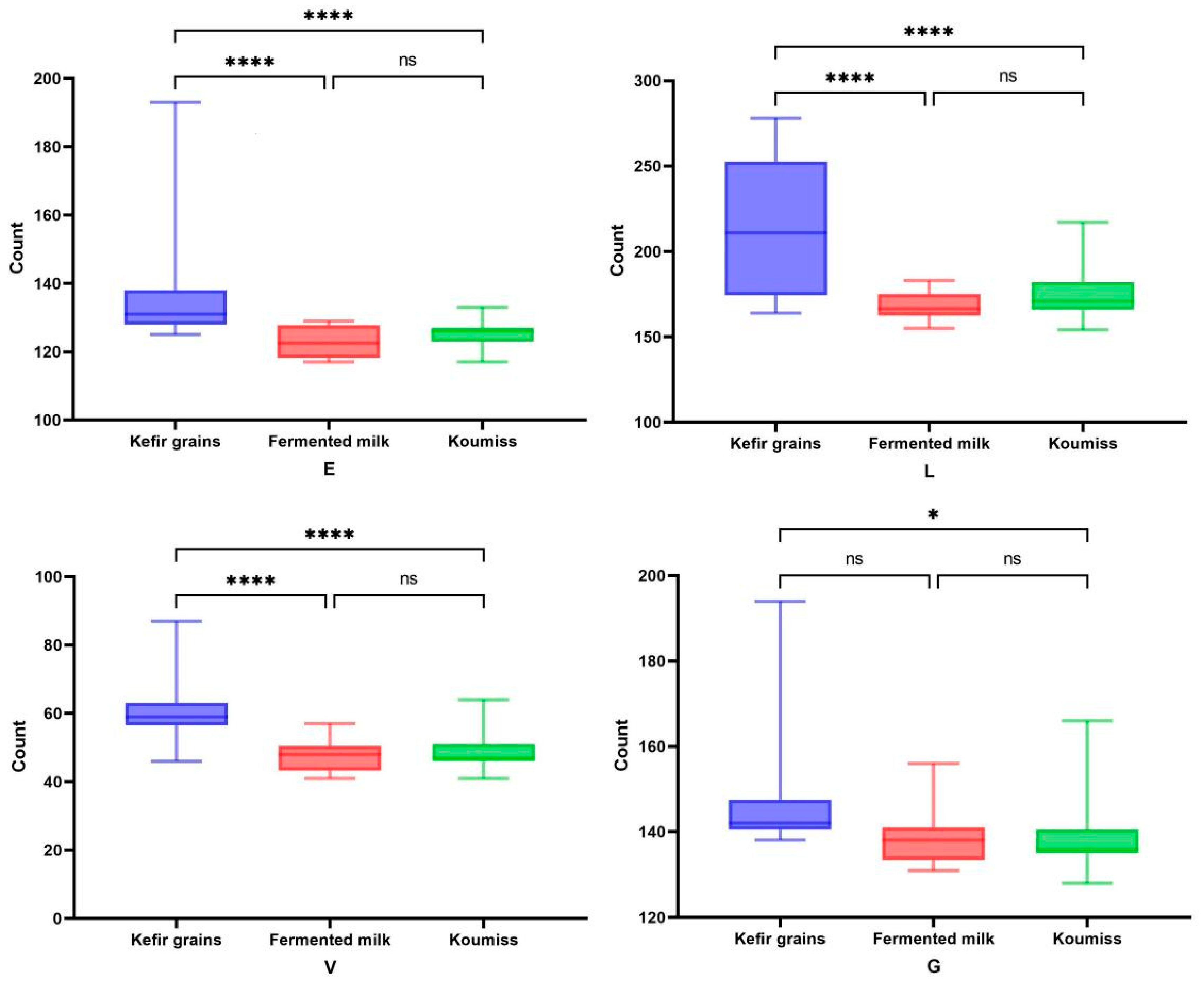
3.6. Analysis of Differences Amongst Isolates
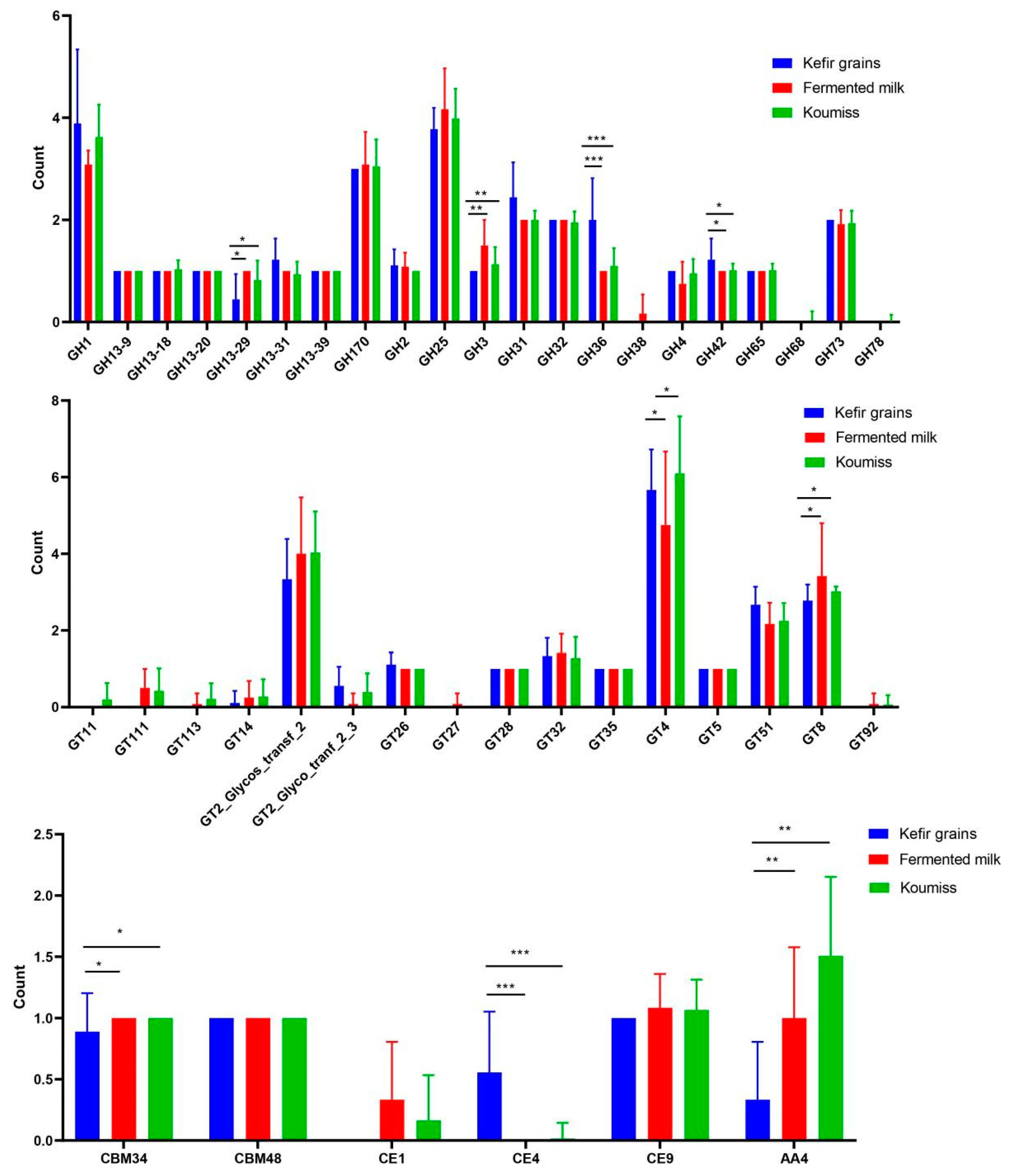
3.7. CAZy Analysis
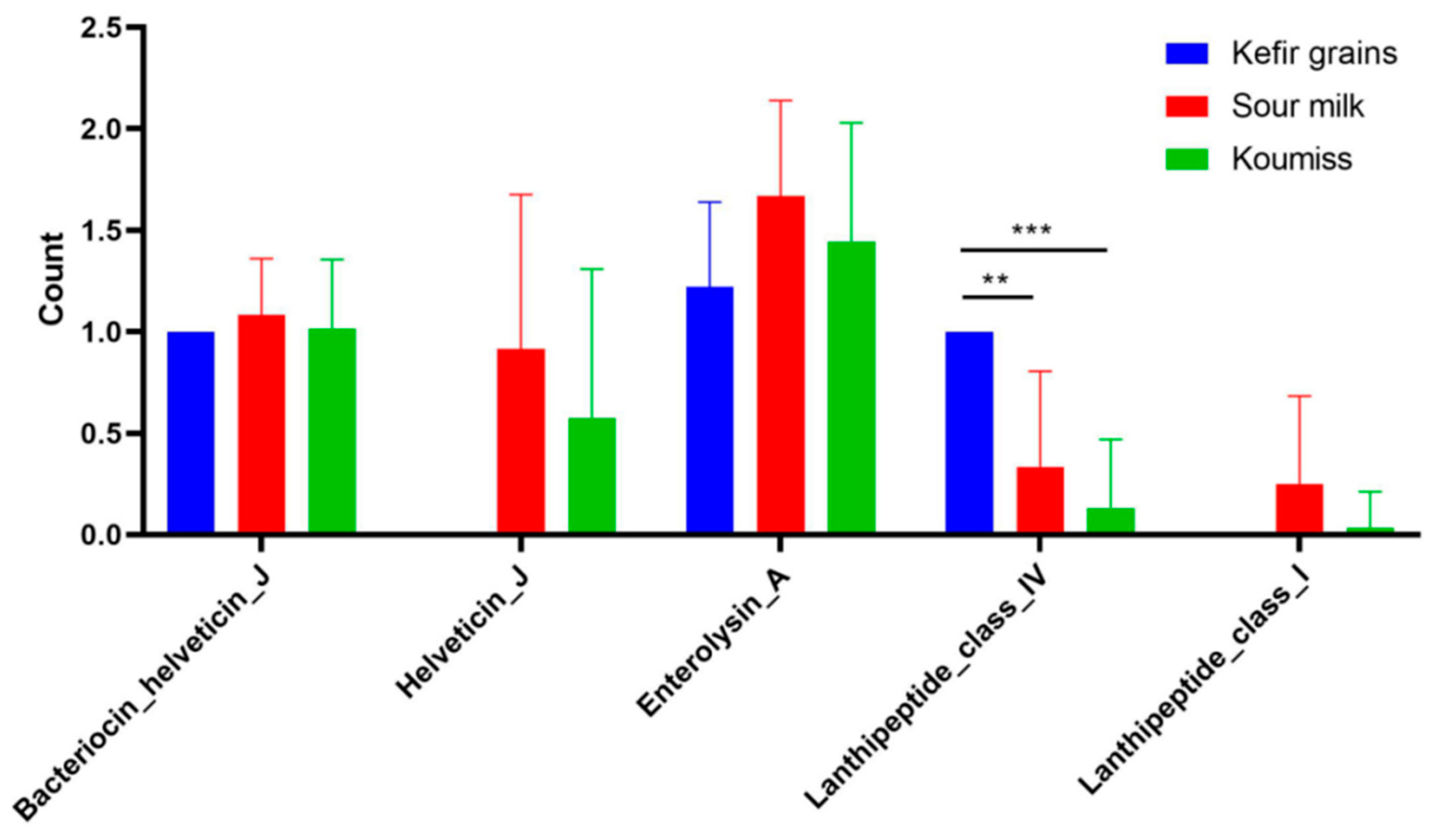
3.8. Bacteriocin Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Garrote, G.L.; Abraham, A.G.; De Antoni, G.L. Chemical and microbiological characterisation of kefir grains. J. Dairy Res. 2001, 68, 639–652. [Google Scholar] [CrossRef] [PubMed]
- McCue, P.P.; Shetty, K. Phenolic antioxidant mobilization during yogurt production from soymilk using Kefir cultures. Process Biochem. 2005, 40, 1791–1797. [Google Scholar] [CrossRef]
- Rodrigues, K.L.; Caputo, L.R.G.; Carvalho, J.C.T.; Evangelista, J.; Schneedorf, J.M. Antimicrobial and healing activity of kefir and kefiran extract. Int. J. Antimicrob. Agents 2005, 25, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Seydim, Z.B. Studies on Fermentative, Microbiological and Biochemical Properties of Kefir and Kefir Grains; Clemson University: America, NC, USA, 2021. [Google Scholar]
- Mukai, T.; Toba, T.; Itoh, T.; Adachi, S. Structural investigation of the capsular polysaccharide from Lactobacillus kefiranofaciens K1. Carbohydr. Res. 1990, 204, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, H.; Watanabe, T.; Fujii, Y.; Mukai, T.; Toba, T.; Adachi, S. Some taxonomical characteristics of encapsulated Lactobacillus sp. KPB-167B isolated from kefir grains and characterization of its extracellular polysaccharide. Int. J. Food Microbiol. 1991, 13, 257–264. [Google Scholar] [CrossRef]
- Toba, T.; Abe, S.; Arihara, K.; Adachi, S. A medium for the isolation of capsular bacteria from kefir grains. Agric. Biol. Chem. 1986, 50, 2673–2674. [Google Scholar]
- Fujisawa, T.; Adachi, S.; Toba, T.; Arihara, K.; Mitsuoka, T. Lactobacillus kefiranofaciens sp. nov. isolated from kefir grains. Int. J. Syst. Evol. Microbiol. 1988, 38, 12–14. [Google Scholar] [CrossRef]
- Vancanneyt, M.; Mengaud, J.; Cleenwerck, I.; Vanhonacker, K.; Hoste, B.; Dawyndt, P.; Degivry, M.; Ringuet, D.; Janssens, D.; Swings, J. Reclassification of Lactobacillus kefirgranum Takizawa et al. 1994 as Lactobacillus kefiranofaciens subsp. kefirgranum subsp. nov. and emended description of L. kefiranofaciens Fujisawa et al. 1988. Int. J. Syst. Evol. Microbiol. 2004, 54, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Hong, W.S.; Chen, Y.P.; Chen, M.J. The antiallergic effect of kefir Lactobacilli. J. Food Sci. 2010, 75, H244–H253. [Google Scholar] [CrossRef]
- Xing, Z.; Tang, W.; Geng, W.; Zheng, Y.; Wang, Y. In vitro and in vivo evaluation of the probiotic attributes of Lactobacillus kefiranofaciens XL10 isolated from Tibetan kefir grain. Appl. Microbiol. Biotechnol. 2017, 101, 2467–2477. [Google Scholar] [CrossRef]
- Hong, W.-S.; Chen, Y.-P.; Dai, T.-Y.; Huang, I.-N.; Chen, M.-J. Effect of heat-inactivated kefir-isolated Lactobacillus kefiranofaciens M1 on preventing an allergic airway response in mice. J. Agric. Food Chem. 2011, 59, 9022–9031. [Google Scholar] [CrossRef]
- Jeong, D.; Kim, D.-H.; Kang, I.-B.; Kim, H.; Song, K.-Y.; Kim, H.-S.; Seo, K.-H. Characterization and antibacterial activity of a novel exopolysaccharide produced by Lactobacillus kefiranofaciens DN1 isolated from kefir. Food Control 2017, 78, 436–442. [Google Scholar] [CrossRef]
- Hardison, R.C. Comparative genomics. PLoS Biol. 2003, 1, e58. [Google Scholar] [CrossRef]
- Chen, C.; Jing, R.; Fangfang, Z.; Zhenmin, L.; Benheng, G. Comparative genomics of Lact. Plantarum. Chin. J. Bioeng. 2013, 33, 35–44. [Google Scholar]
- Chao, Y. Population Genomics of Vibrio Parahaemolyticus; Academy of Military Sciences: Beijing, China, 2019. [Google Scholar]
- Xiaona, A.; Weicheng, L.; Jie, Y.; Lin, P.; Lanxin, M.; Caiqing, Y.; Heping, Z. Comparative genomics analysis of genetic diversity and habitat adaptation of L. reuteri from different sources. China J. Microbiol. 2020, 60, 875–886. [Google Scholar]
- Aktas, B.; Budinich, M.; Hoza, L.; Rankin, S.A.; Broadbent, J.R.; Steele, J.L. Shelf-life studies of putative probiotic Lacticaseibacillus casei strains in milk and model yogurt. Food Sci. Technol. Int. 2022, 5, 21–27. [Google Scholar] [CrossRef]
- Verce, M.; De Vuyst, L.; Weckx, S. Comparative genomics of Lactobacillus fermentum suggests a free-living lifestyle of this lactic acid bacterial species. Food Microbiol. 2020, 89, 103448. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Ahmed, Z.; Bai, X.; Wang, J. Complete genome sequence of Lactobacillus kefiranofaciens ZW3. Am. Soc. Microbiol. 2021, 193, 4280–4281. [Google Scholar] [CrossRef] [Green Version]
- Xing, Z.; Geng, W.; Li, C.; Sun, Y.; Wang, Y. Comparative genomics of Lactobacillus kefiranofaciens ZW3 and related members of Lactobacillus. spp reveal adaptations to dairy and gut environments. Sci. Rep. 2017, 7, 12827. [Google Scholar] [CrossRef] [Green Version]
- Francis, F.; Kim, J.; Ramaraj, T.; Farmer, A.; Rush, M.C.; Ham, J.H. Comparative genomic analysis of two Burkholderia glumae strains from different geographic origins reveals a high degree of plasticity in genome structure associated with genomic islands. Mol. Genet. Genom. 2013, 288, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. Gigascience 2012, 217X, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. dbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic. Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [Green Version]
- Yanping, W.; Ting, Y.; Yaqi, W.; Yuanwang, W.; Meng, W. Research progress of Lactobacillus kefiranofaciens subspecies and its research progress. Chin. J. Microbiol. 2022, 62, 414–420. [Google Scholar]
- Richa, H. Population Genetics and Functional Genomics of Lactobacillus helveticus in Naturally Fermented Dairy Products; Inner Mongolia Agricultural University: Hohhot, China, 2021. [Google Scholar]
- Hongli, W.; Yong, X.; Jian, L.; Lihui, G.; Minnan, L. Research progress of acetylxylanesterase. Chin. J. Bioeng. 2015, 36, 102–110. [Google Scholar]
- Ewing, T.A.; Gygli, G.; Fraaije, M.W.; van Berkel, W.J. Vanillyl alcohol oxidase. Enzymes 2020, 47, 87–116. [Google Scholar]
- Collins, F.W.; O’Connor, P.M.; O’Sullivan, O.; Gómez-Sala, B.; Rea, M.C.; Hill, C.; Ross, R.P. Bacteriocin Gene-Trait matching across the complete Lactobacillus Pan-genome. Sci. Rep. 2017, 7, 3481. [Google Scholar] [CrossRef] [Green Version]
- Joerger, M.; Klaenhammer, T. Cloning, expression, and nucleotide sequence of the Lactobacillus helveticus 481 gene encoding the bacteriocin helveticin J. J. Bacteriol. 1990, 172, 6339–6347. [Google Scholar] [CrossRef] [Green Version]
- Nilsen, T.; Nes, I.F.; Holo, H. Enterolysin A, a cell wall-degrading bacteriocin from Enterococcus faecalis LMG 2333. Appl. Environ. Microbiol. 2003, 69, 2975–2984. [Google Scholar] [CrossRef] [Green Version]
- Ji, Q.; Yang, M.; Yi, Z.; Chenyu, L.; Chun, L. Research Progress of Main Nutrients in Milk of Different Mammals; Dairy Science and Technology: Shanghai, China, 2021; Volume 44, p. 5. [Google Scholar]
- El Kafsi, H.; Binesse, J.; Loux, V.; Buratti, J.; Boudebbouze, S.; Dervyn, R.; Kennedy, S.; Galleron, N.; Quinquis, B.; Batto, J.-M. Lactobacillus delbrueckii ssp. lactis and ssp. bulgaricus: A chronicle of evolution in action. BMC Genom. 2014, 15, 407. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Pan, Y.; Li, B.; Ou, J.; Zhang, J.; Chen, Y.; Peng, X.; Chen, L. Molecular cloning and antimicrobial activity of enterolysin A and helveticin J of bacteriolysins from metagenome of Chinese traditional fermented foods. Food Control 2013, 31, 499–507. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Isolation Source | Region | Genome Size (Mbp) | GC Content (%) | CDS | tRNA | Accession No. |
|---|---|---|---|---|---|---|---|
| 1207 | Kefir grains | Unknown | 2.07 | 37.62 | 2156 | 63 | GCA_014656585.1 |
| ATCC 43761 | Kefir grains | Denmark | 2.18 | 37.21 | 2344 | 59 | GCA_900103655.1 |
| DSM 10550 | Kefir grains | Japan | 1.99 | 37.47 | 2087 | 56 | GCA_001434195.1 |
| DSM 5016 | Kefir grains | Denmark | 2.15 | 37.22 | 2310 | 46 | GCA_001435275.1 |
| JCM 6985 | Kefir grains | Denmark | 2.15 | 37.21 | 2435 | 50 | GCA_000615685.1 |
| JCM 8572 | Kefir grains | Japan | 1.97 | 37.42 | 2913 | 53 | GCA_001311335.1 |
| KR | Kefir grains | Germany | 2.03 | 37.4 | 2138 | 59 | GCA_002276565.1 |
| LKK75 | Kefir grains | Unknown | 2.21 | 37.38 | 2385 | 63 | GCA_009184665.1 |
| ZW3 | Kefir grains | Tibet | 2.25 | 37.36 | 2420 | 61 | GCA_000214785.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, R.; Liu, C.; Li, Y.; Liu, Q.; Su, X.; Peng, Q.; Lei, X.; Li, W.; Menghe, B.; Bao, Q.; et al. Comparative Genomics Analysis of Habitat Adaptation by Lactobacillus kefiranofaciens. Foods 2023, 12, 1606. https://doi.org/10.3390/foods12081606
Luo R, Liu C, Li Y, Liu Q, Su X, Peng Q, Lei X, Li W, Menghe B, Bao Q, et al. Comparative Genomics Analysis of Habitat Adaptation by Lactobacillus kefiranofaciens. Foods. 2023; 12(8):1606. https://doi.org/10.3390/foods12081606
Chicago/Turabian StyleLuo, Rui, Chen Liu, Yu Li, Qing Liu, Xin Su, Qingting Peng, Xueyan Lei, Weicheng Li, Bilige Menghe, Qiuhua Bao, and et al. 2023. "Comparative Genomics Analysis of Habitat Adaptation by Lactobacillus kefiranofaciens" Foods 12, no. 8: 1606. https://doi.org/10.3390/foods12081606
APA StyleLuo, R., Liu, C., Li, Y., Liu, Q., Su, X., Peng, Q., Lei, X., Li, W., Menghe, B., Bao, Q., & Liu, W. (2023). Comparative Genomics Analysis of Habitat Adaptation by Lactobacillus kefiranofaciens. Foods, 12(8), 1606. https://doi.org/10.3390/foods12081606