

Ex Vivo Study of Colon Health, Contractility and Innervation in Male and Female Rats after Regular Exposure to Instant Cascara Beverage

 , , ,
, , ,  ,
,  and
and 
Abstract

1. Introduction
2. Resources and Procedures
2.1. Ethical Declaration
2.2. Instant Cascara Beverage
2.3. Experimental Groups and Study Animals
2.4. Methodological Approach
2.5. Colon Wall Health: Histological Analysis
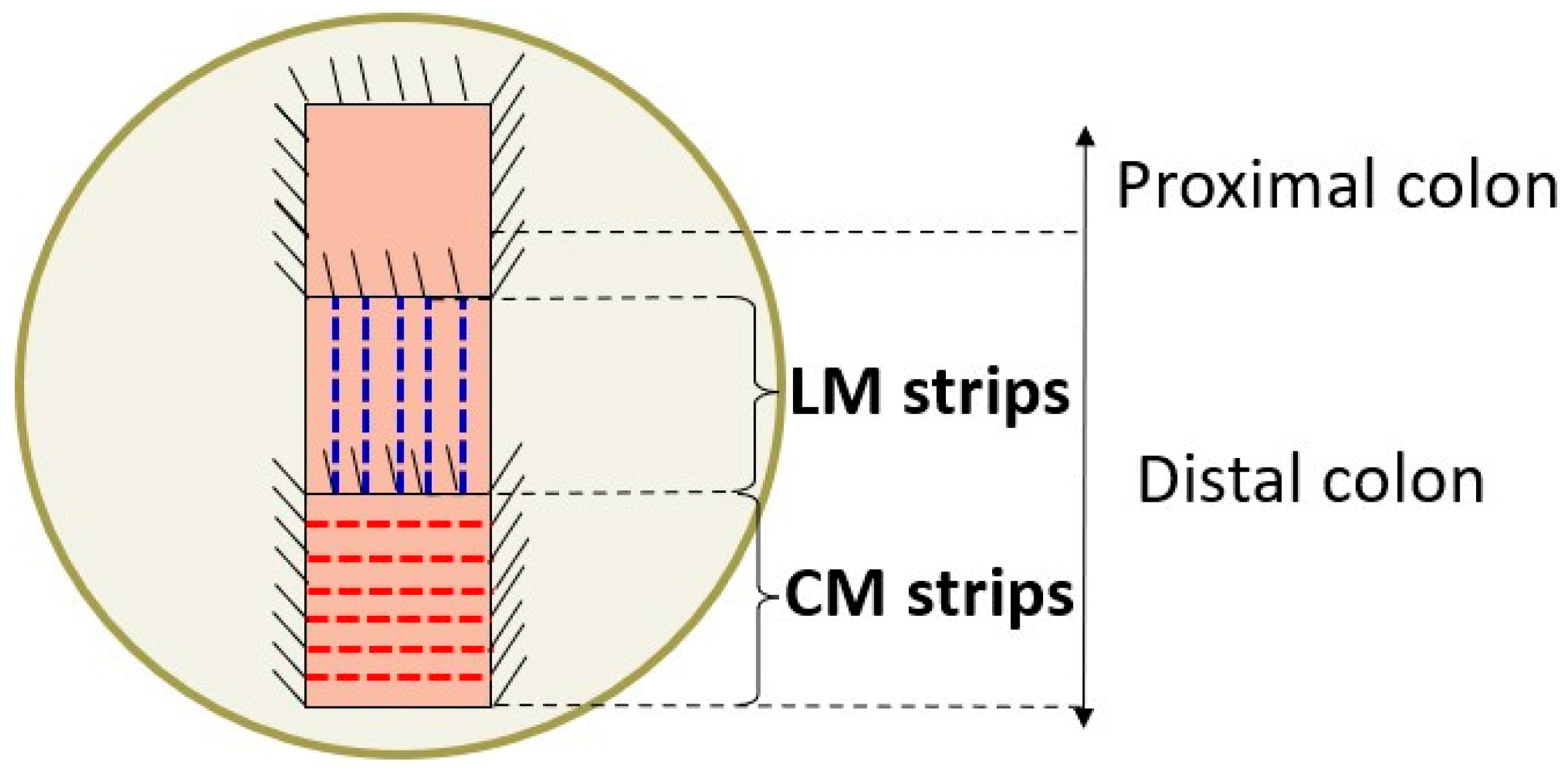
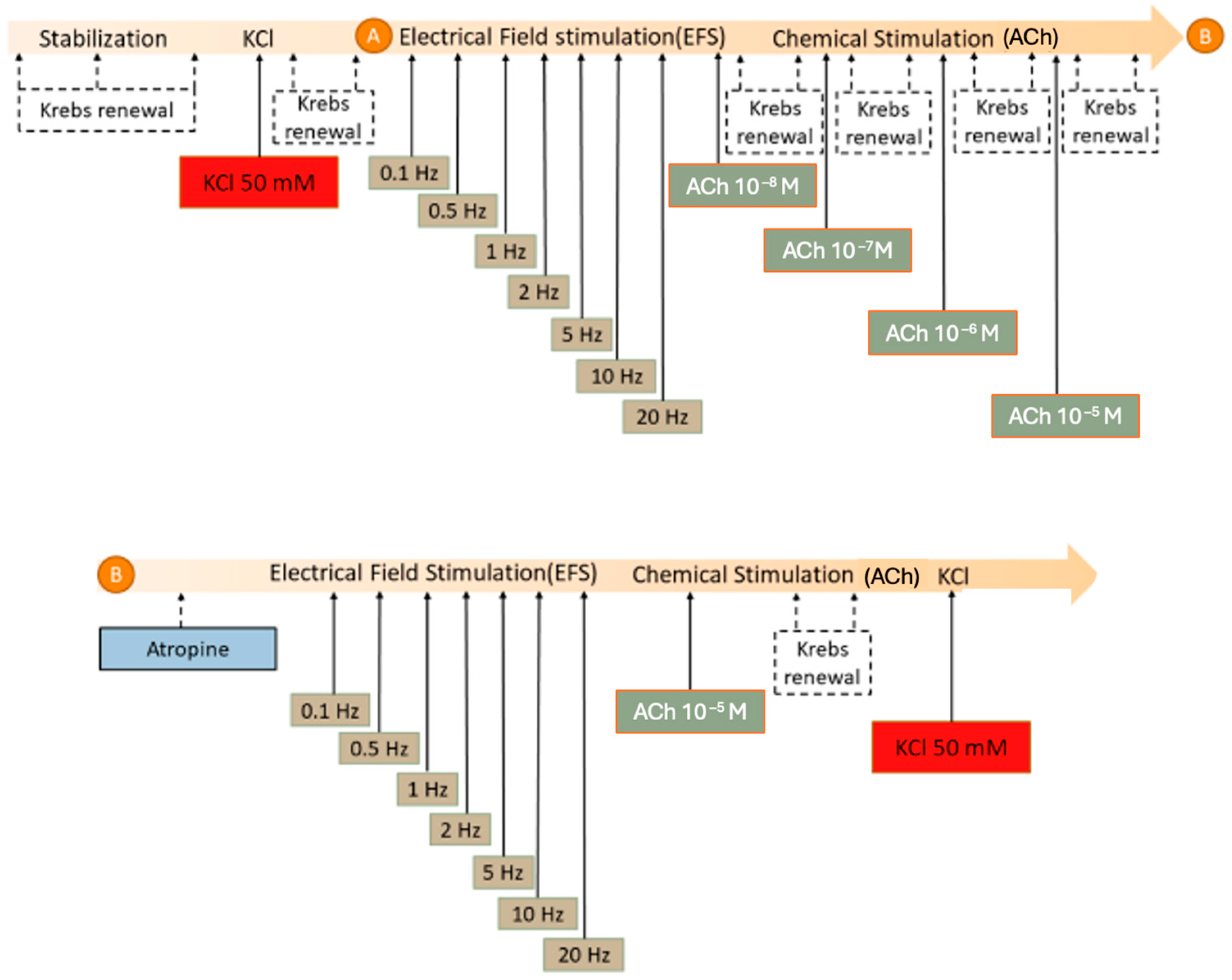
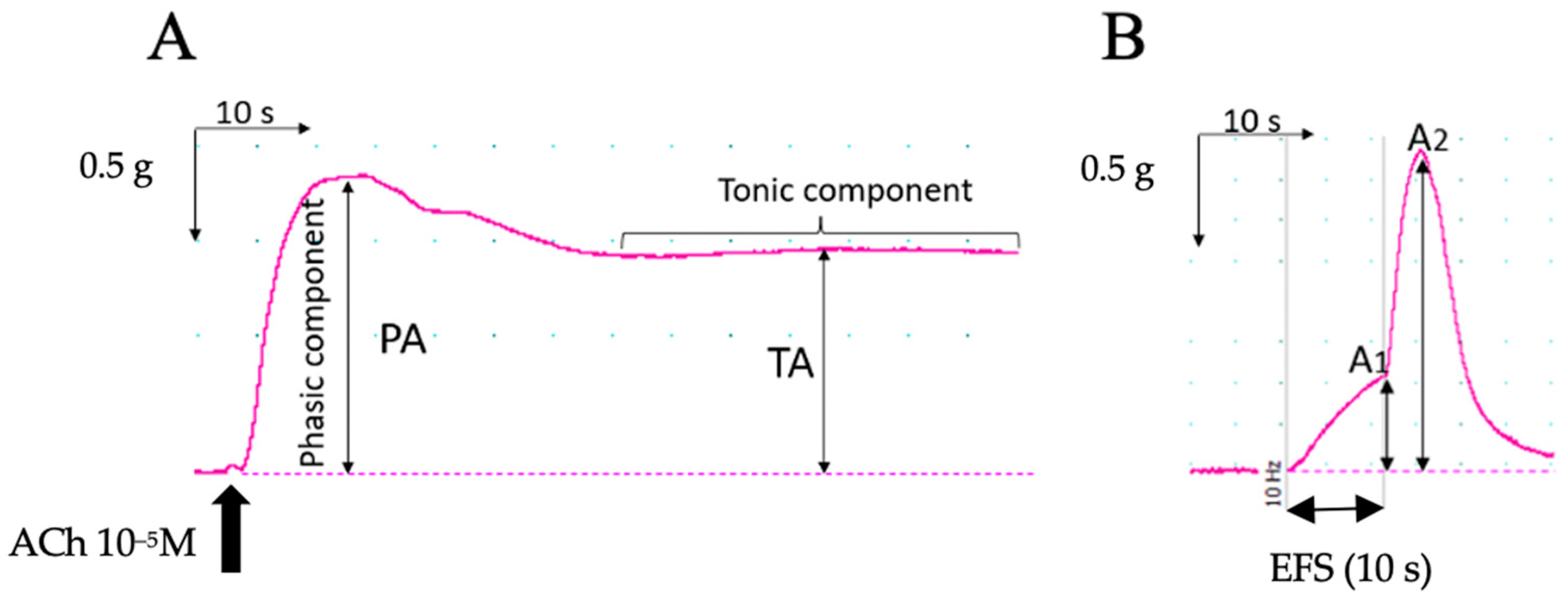
2.6. Colon Contractility: Organ Bath Study
2.7. Colonic Innervation: Immunohistochemistry Analysis
2.7.1. Colonic Sections
2.7.2. Whole-Mount Longitudinal Muscle–Myenteric Plexus Preparations
2.8. Drugs and Reagents
2.9. Data Evaluation
3. Results
3.1. General Health
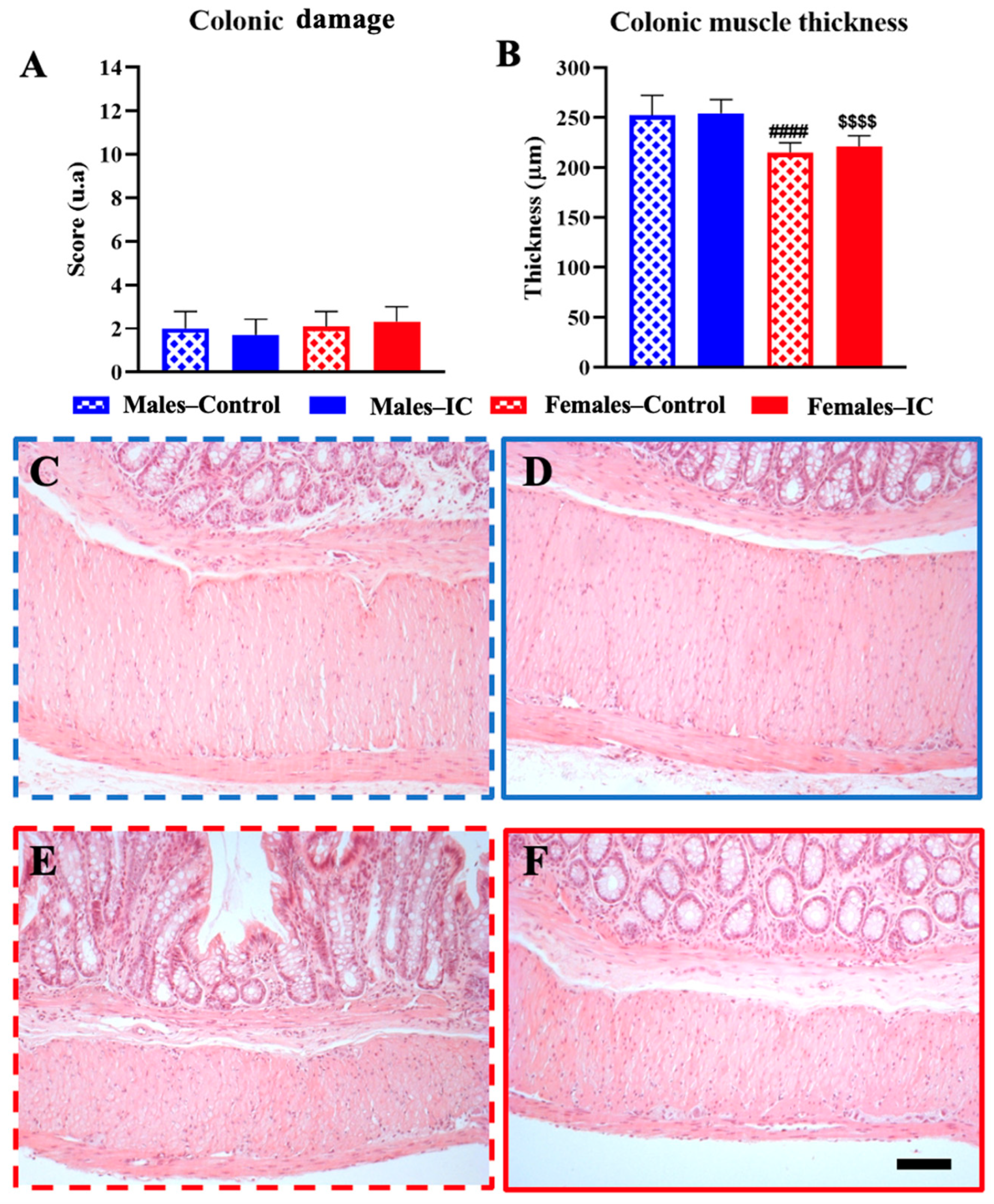
3.2. Colon Wall Health: Histological Analysis
3.3. Colon Contractility: Organ Bath Study
3.3.1. Responses to KCl Stimulation
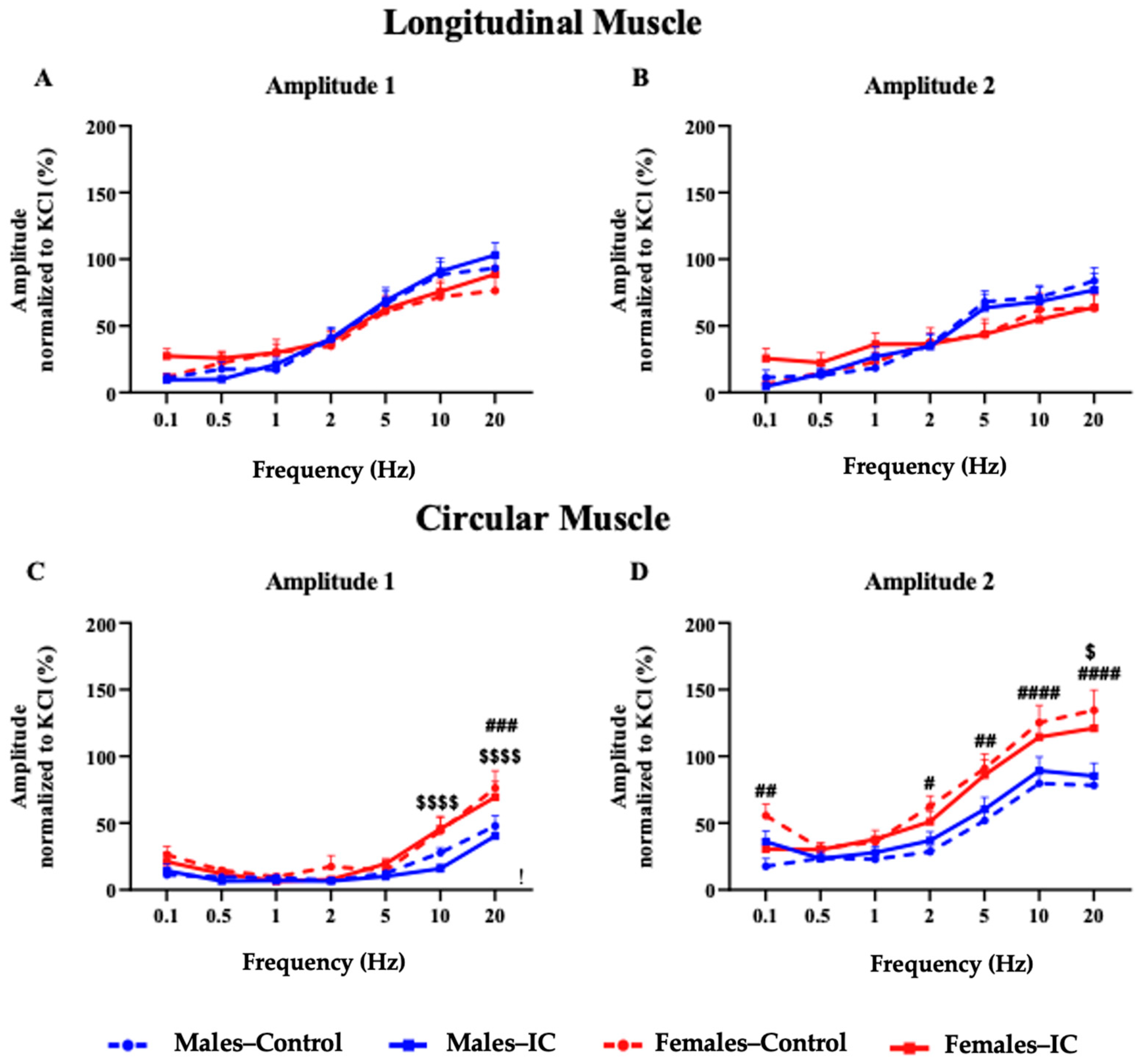
3.3.2. Electrical Field Stimulation
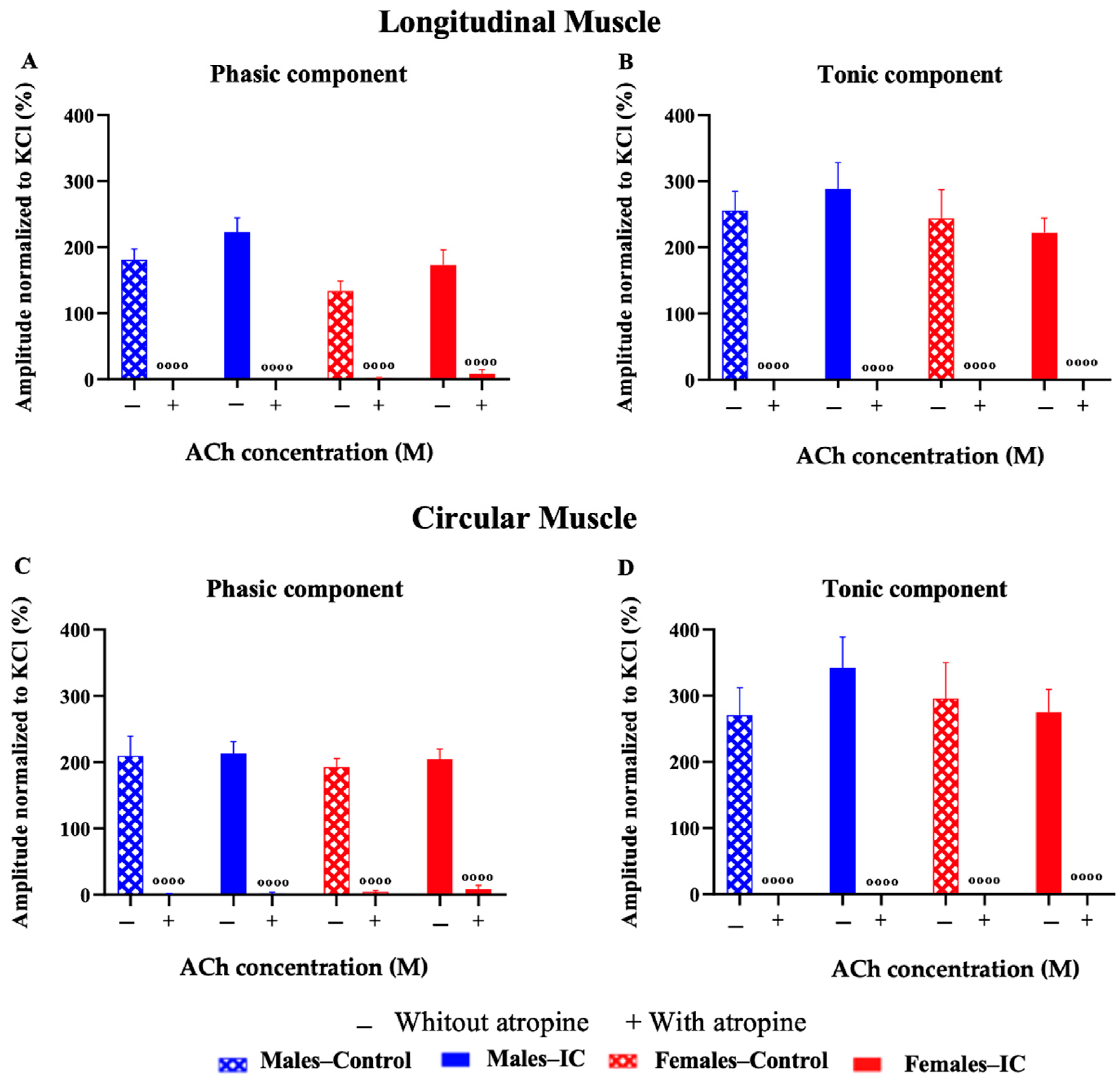
3.3.3. Chemical Stimulation with Acetylcholine
3.4. Colonic Innervation: Immunohistochemistry Analysis
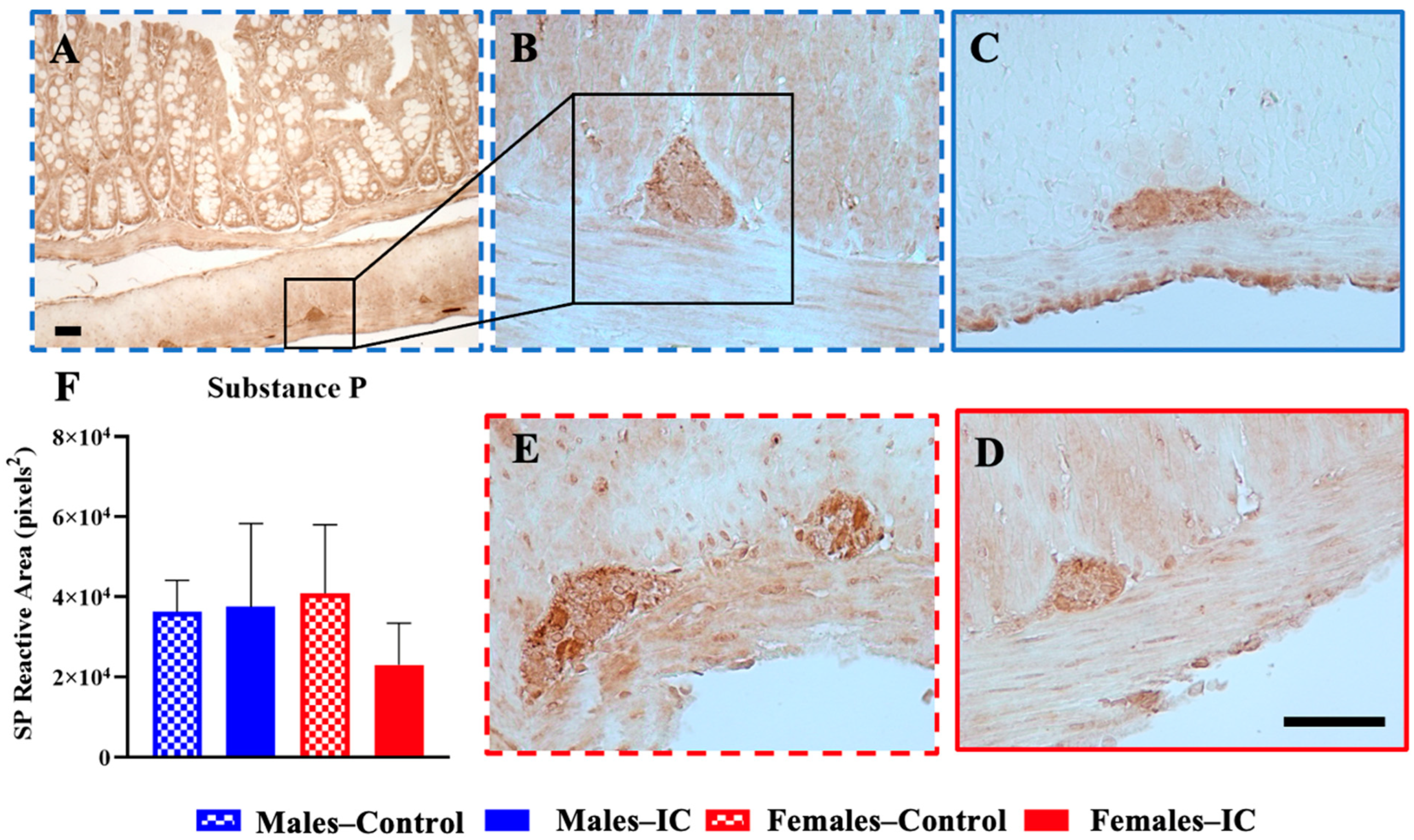
3.4.1. Substance P
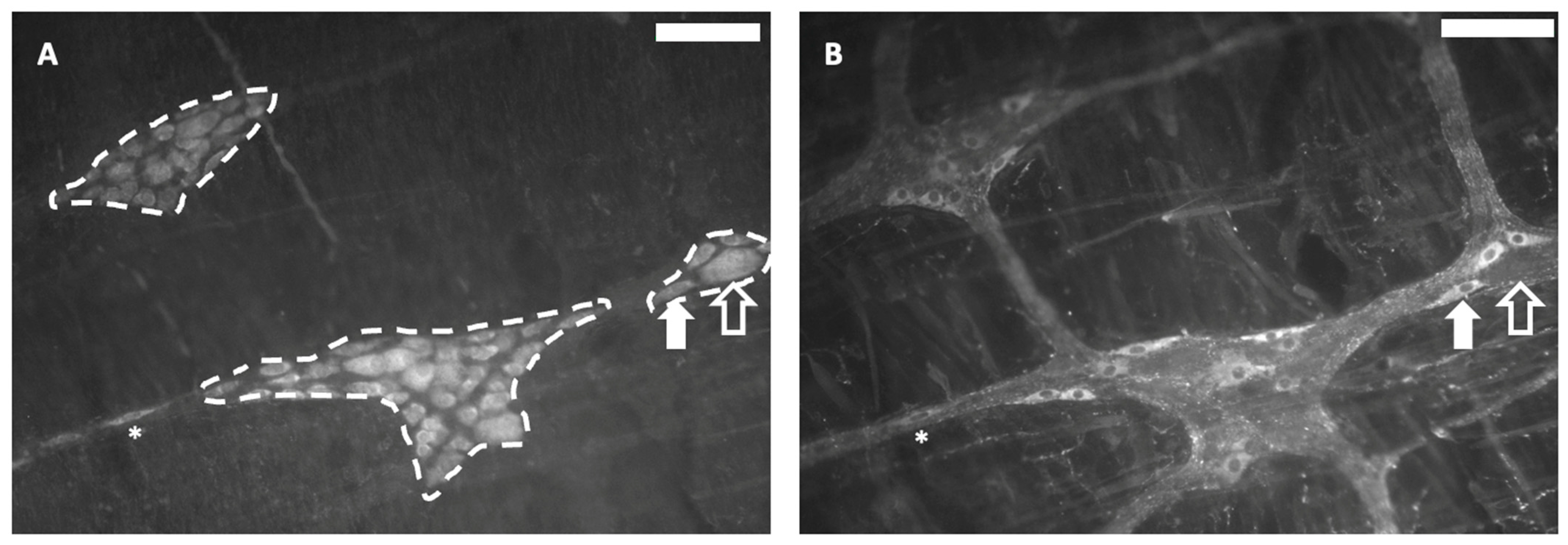
3.4.2. Myenteric Plexus Study in Whole-Mount LMMP Preparations
4. Discussion
4.1. Colon Wall Health
4.2. Colon Contractility
4.3. Colonic Innervation
4.4. Significance, Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sensoy, I. A Review on the Food Digestion in the Digestive Tract and the Used in Vitro Models. Curr. Res. Food Sci. 2021, 4, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Suganya, K.; Koo, B.S. Gut–Brain Axis: Role of Gut Microbiota on Neurological Disorders and How Probiotics/Prebiotics Beneficially Modulate Microbial and Immune Pathways to Improve Brain Functions. Int. J. Mol. Sci. 2020, 21, 7551. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The Enteric Nervous System: Normal Functions and Enteric Neuropathies. Neurogastroenterol. Motil. 2008, 20, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Spencer, N.J.; Hu, H. Enteric Nervous System: Sensory Transduction, Neural Circuits and Gastrointestinal Motility. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 338. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Baumann, P.; Tüscher, O.; Schick, S.; Endres, K. The Aging Enteric Nervous System. Int. J. Mol. Sci. 2023, 24, 9471. [Google Scholar] [CrossRef] [PubMed]
- Holland, A.M.; Bon-Frauches, A.C.; Keszthelyi, D.; Melotte, V.; Boesmans, W. The Enteric Nervous System in Gastrointestinal Disease Etiology. Cell. Mol. Life Sci. 2021, 78, 4713. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems. Ann. Gastroenterol. Q. Publ. Hell. Soc. Gastroenterol. 2015, 28, 203. [Google Scholar]
- Liu, L.; Huh, J.R.; Shah, K. Microbiota and the Gut-Brain-Axis: Implications for New Therapeutic Design in the CNS. EBioMedicine 2022, 77, 103908. [Google Scholar] [CrossRef]
- Karakan, T.; Ozkul, C.; Akkol, E.K.; Bilici, S.; Sobarzo-Sánchez, E.; Capasso, R. Gut-Brain-Microbiota Axis: Antibiotics and Functional Gastrointestinal Disorders. Nutrients 2021, 13, 389. [Google Scholar] [CrossRef]
- Rao, M.; Gershon, M.D. The Bowel and beyond: The Enteric Nervous System in Neurological Disorders. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 517. [Google Scholar] [CrossRef]
- Mittal, R.; Debs, L.H.; Patel, A.P.; Nguyen, D.; Patel, K.; O’Connor, G.; Grati, M.; Mittal, J.; Yan, D.; Eshraghi, A.A.; et al. Neurotransmitters: The Critical Modulators Regulating Gut-Brain Axis. J. Cell Physiol. 2017, 232, 2359. [Google Scholar] [CrossRef] [PubMed]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486. [Google Scholar] [CrossRef]
- Iriondo-DeHond, A.; Elizondo, A.S.; Iriondo-DeHond, M.; Ríos, M.B.; Mufari, R.; Mendiola, J.A.; Ibañez, E.; del Castillo, M.D. Assessment of Healthy and Harmful Maillard Reaction Products in a Novel Coffee Cascara Beverage: Melanoidins and Acrylamide. Foods 2020, 9, 620. [Google Scholar] [CrossRef] [PubMed]
- López-Parra, M.B.; Gómez-Domínguez, I.; Iriondo-DeHond, M.; Villamediana Merino, E.; Sánchez-Martín, V.; Mendiola, J.A.; Iriondo-DeHond, A.; del Castillo, M.D. The Impact of the Drying Process on the Antioxidant and Anti-Inflammatory Potential of Dried Ripe Coffee Cherry Pulp Soluble Powder. Foods 2024, 13, 1114. [Google Scholar] [CrossRef] [PubMed]
- Lachenmeier, W.; Sánchez-Martín, V.; López-Parra, M.B.; Iriondo-Dehond, A.; Haza, A.I.; Morales, P.; Dolores Del Castillo, M. Instant Cascara: A Potential Sustainable Promoter of Gastrointestinal Health. Proceedings 2023, 89, 21. [Google Scholar] [CrossRef]
- Iriondo-Dehond, A.; Uranga, J.A.; Del Castillo, M.D.; Abalo, R. Effects of Coffee and Its Components on the Gastrointestinal Tract and the Brain–Gut Axis. Nutrients 2020, 13, 88. [Google Scholar] [CrossRef] [PubMed]
- Romaní-Pérez, M.; Bullich-Vilarrubias, C.; López-Almela, I.; Liébana-García, R.; Olivares, M.; Sanz, Y. The Microbiota and the Gut–Brain Axis in Controlling Food Intake and Energy Homeostasis. Int. J. Mol. Sci. 2021, 22, 5830. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Meckling, K.A.; Marcone, M.F.; Kakuda, Y.; Tsao, R. Synergistic, Additive, and Antagonistic Effects of Food Mixtures on Total Antioxidant Capacities. J. Agric. Food Chem. 2011, 59, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.W.; Wang, C.C.; Sung, W.W.; Ting, W.C.; Lin, C.C.; Tsai, M.C. The Effect of Coffee/Caffeine on Postoperative Ileus Following Elective Colorectal Surgery: A Meta-Analysis of Randomized Controlled Trials. Int. J. Color. Dis. 2022, 37, 623–630. [Google Scholar] [CrossRef]
- Nasi, M.; De Gaetano, A.; Carnevale, G.; Bertoni, L.; Selleri, V.; Zanini, G.; Pisciotta, A.; Caramaschi, S.; Reggiani Bonetti, L.; Farinetti, A.; et al. Effects of Energy Drink Acute Assumption in Gastrointestinal Tract of Rats. Nutrients 2022, 14, 1928. [Google Scholar] [CrossRef]
- Gallego-Barceló, P.; Bagues, A.; Benítez-Álvarez, D.; López-Tofiño, Y.; Gálvez-Robleño, C.; López-Gómez, L.; Dolores, M.; Castillo, D.; Abalo, R. Evaluation of the Effects of Instant Cascara Beverage on the Brain-Gut Axis of Healthy Male and Female Rats. Nutrients 2023, 16, 65. [Google Scholar] [CrossRef]
- Del Castillo, M.D.; Ibáñez, E.; Amigo-Benavent, M.; Herrero, M.; Plaza, M.; Ullate, M. Application of Products of Coffee Silverskin in Anti-Ageing Cosmetics and Functional Food. WO2013004873A1, 10 January 2013. [Google Scholar]
- Iriondo-Dehond, A.; Iriondo-Dehond, M.; Del Castillo, M.D. Applications of Compounds from Coffee Processing By-Products. Biomolecules 2020, 10, 1219. [Google Scholar] [CrossRef]
- López-Gómez, L.; López-Tofiño, Y.; Abalo, R. Dependency on Sex and Stimulus Quality of Nociceptive Behavior in a Conscious Visceral Pain Rat Model. Neurosci. Lett. 2021, 746, 135667. [Google Scholar] [CrossRef]
- Cora, M.C.; Kooistra, L.; Travlos, G. Vaginal Cytology of the Laboratory Rat and Mouse:Review and Criteria for the Staging of the Estrous Cycle Using Stained Vaginal Smears. Toxicol. Pathol. 2015, 43, 776–793. [Google Scholar] [CrossRef]
- Ranganathan, P.; Jayakumar, C.; Manicassamy, S.; Ramesh, G. CXCR2 Knockout Mice Are Protected against DSS-Colitis-Induced Acute Kidney Injury and Inflammation. Am. J. Physiol. Ren. Physiol. 2013, 305, 1422–1427. [Google Scholar] [CrossRef]
- Uranga, J.A.; García-Martínez, J.M.; García-Jiménez, C.; Vera, G.; Martín-Fontelles, M.I.; Abalo, R. Alterations in the Small Intestinal Wall and Motor Function after Repeated Cisplatin in Rat. Neurogastroenterol. Motil. 2017, 29, e13047. [Google Scholar] [CrossRef]
- López-Tofiño, Y.; Barragán del Caz, L.F.; Benítez-Álvarez, D.; Molero-Mateo, P.; Nurgali, K.; Vera, G.; Bagües, A.; Abalo, R. Contractility of Isolated Colonic Smooth Muscle Strips from Rats Treated with Cancer Chemotherapy: Differential Effects of Cisplatin and Vincristine. Front. Neurosci. 2023, 17, 1304609. [Google Scholar] [CrossRef] [PubMed]
- Straface, M.; Makwana, R.; Palmer, A.; Rombolà, L.; Aleong, J.C.; Morrone, L.A.; Sanger, G.J. Inhibition of Neuromuscular Contractions of Human and Rat Colon by Bergamot Essential Oil and Linalool: Evidence to Support a Therapeutic Action. Nutrients 2020, 12, 1381. [Google Scholar] [CrossRef] [PubMed]
- Vera, G.; Castillo, M.; Cabezos, P.A.; Chiarlone, A.; Martín, M.I.; Gori, A.; Pasquinelli, G.; Barbara, G.; Stanghellini, V.; Corinaldesi, R.; et al. Enteric Neuropathy Evoked by Repeated Cisplatin in the Rat. Neurogastroenterol. Motil. 2011, 23, 370-e163. [Google Scholar] [CrossRef] [PubMed]
- Abalo, R.; Rivera, A.J.; Vera, G.; Martín, M.I. Ileal Myenteric Plexus in Aged Guinea-Pigs: Loss of Structure and Calretinin-Immunoreactive Neurones. Neurogastroenterol. Motil. 2005, 17, 123–132. [Google Scholar] [CrossRef]
- Chen, Y.; Kim, M.; Paye, S.; Benayoun, B.A. Sex as a Biological Variable in Nutrition Research: From Human Studies to Animal Models. Annu. Rev. Nutr. 2022, 42, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Thein, W.; Po, W.W.; Kim, D.M.; Sohn, U.D. The Altered Signaling on EFS-Induced Colon Contractility in Diabetic Rats. Biomol. Ther. 2020, 28, 328. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.; Suárez, L.; Andrés, M.T.; Flórez, B.H.; Bordallo, J.; Riestra, S.; Cantabrana, B. Modulatory Effect of Intestinal Polyamines and Trace Amines on the Spontaneous Phasic Contractions of the Isolated Ileum and Colon Rings of Mice. Food Nutr. Res. 2017, 61, 1321948. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Qiao, X.; Falone, A.E.; Reslan, O.M.; Sheppard, S.J.; Khalil, R.A. Gender-Specific Reduction in Contraction Is Associated with Increased Estrogen Receptor Expression in Single Vascular Smooth Muscle Cells of Female Rat. Cell. Physiol. Biochem. 2010, 26, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Al-Shboul, O.A.; Al-Dwairi, A.N.; Alqudah, M.A.; Mustafa, A.G. Gender Differences in the Regulation of MLC20 Phosphorylation and Smooth Muscle Contraction in Rat Stomach. Biomed. Rep. 2018, 8, 283. [Google Scholar] [CrossRef] [PubMed]
- Tokutomi, Y.; Tokutomi, N.; Nishi, K. The Properties of Ryanodine-Sensitive Ca2+ Release in Mouse Gastric Smooth Muscle Cells. Br. J. Pharmacol. 2001, 133, 125. [Google Scholar] [CrossRef] [PubMed]
- Tazzeo, T.; Bates, G.; Roman, H.N.; Lauzon, A.M.; Khasnis, M.D.; Eto, M.; Janssen, L.J. Caffeine Relaxes Smooth Muscle through Actin Depolymerization. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 303, L334. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.T.; Hennig, G.W.; Fleming, N.W.; Keef, K.D.; Spencer, N.J.; Ward, S.M.; Sanders, K.M.; Smith, T.K. The Mechanism and Spread of Pacemaker Activity through Myenteric Interstitial Cells of Cajal in Human Small Intestine. Gastroenterology 2007, 132, 1852–1865. [Google Scholar] [CrossRef]
- Ratz, P.H.; Berg, K.M.; Urban, N.H.; Miner, A.S. Regulation of Smooth Muscle Calcium Sensitivity: KCl as a Calcium-Sensitizing Stimulus. Am. J. Physiol. Cell Physiol. 2005, 288, C769–C783. [Google Scholar] [CrossRef]
- Montgomery, L.E.A.; Tansey, E.A.; Johnson, C.D.; Roe, S.M.; Quinn, J.G. Autonomic Modification of Intestinal Smooth Muscle Contractility. Adv. Physiol. Educ. 2016, 40, 104–109. [Google Scholar] [CrossRef]
- Zhou, H.; Kong, D.H.; Pan, Q.W.; Wang, H.H. Sources of Calcium in Agonist-Induced Contraction of Rat Distal Colon Smooth Muscle in Vitro. World J. Gastroenterol. 2008, 14, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Broad, J.; Kung, V.W.S.; Palmer, A.; Elahi, S.; Karami, A.; Darreh-Shori, T.; Ahmed, S.; Thaha, M.A.; Carroll, R.; Chin-Aleong, J.; et al. Changes in Neuromuscular Structure and Functions of Human Colon during Ageing Are Region-Dependent. Gut 2019, 68, 1210–1223. [Google Scholar] [CrossRef] [PubMed]
- Hegde, S.; Shi, D.W.; Johnson, J.C.; Geesala, R.; Zhang, K.; Lin, Y.M.; Shi, X.Z. Mechanistic Study of Coffee Effects on Gut Microbiota and Motility in Rats. Nutrients 2022, 14, 4877. [Google Scholar] [CrossRef] [PubMed]
- Aulí, M.; Martínez, E.; Gallego, D.; Opazo, A.; Espín, F.; Martí-Gallostra, M.; Jiménez, M.; Clavé, P. Effects of Excitatory and Inhibitory Neurotransmission on Motor Patterns of Human Sigmoid Colon in Vitro. Br. J. Pharmacol. 2008, 155, 1043. [Google Scholar] [CrossRef] [PubMed]
- Sanger, G.J.; Broad, J.; Kung, V.; Knowles, C.H. Translational Neuropharmacology: The Use of Human Isolated Gastrointestinal Tissues. Br. J. Pharmacol. 2013, 168, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Yamato, S.; Kurematsu, A.; Amano, T.; Ariga, H.; Ando, T.; Komaki, G.; Wada, K. Urocortin 1: A Putative Excitatory Neurotransmitter in the Enteric Nervous System. Neurogastroenterol. Motil. 2020, 32, e13842. [Google Scholar] [CrossRef] [PubMed]
- Hinds, N.M.; Ullrich, K.; Smid, S.D. Cannabinoid 1 (CB1) Receptors Coupled to Cholinergic Motorneurones Inhibit Neurogenic Circular Muscle Contractility in the Human Colon. Br. J. Pharmacol. 2006, 148, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Broad, J.; Mukherjee, S.; Samadi, M.; Martin, J.E.; Dukes, G.E.; Sanger, G.J. Regional- and Agonist-Dependent Facilitation of Human Neurogastrointestinal Functions by Motilin Receptor Agonists. Br. J. Pharmacol. 2012, 167, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Micci, M.A.; Murthy, K.S.; Pasricha, P.J. Substance P Is Essential for Maintaining Gut Muscle Contractility: A Novel Role for Coneurotransmission Revealed by Botulinum Toxin. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G839. [Google Scholar] [CrossRef]
- Drimousis, S.; Markus, I.; Murphy, T.v.; Shevy Perera, D.; Phan-Thien, K.C.; Zhang, L.; Liu, L. Gender-Related Differences of Tachykinin NK 2 Receptor Expression and Activity in Human Colonic Smooth Muscle. J. Pharmacol. Exp. Ther. 2020, 375, 28–39. [Google Scholar] [CrossRef]
- Di Natale, M.R.; Hunne, B.; Liew, J.J.M.; Fothergill, L.J.; Stebbing, M.J.; Furness, J.B. Morphologies, Dimensions and Targets of Gastric Nitric Oxide Synthase Neurons. Cell Tissue Res. 2022, 388, 19. [Google Scholar] [CrossRef]
- Idrizaj, E.; Traini, C.; Vannucchi, M.G.; Baccari, M.C. Nitric Oxide: From Gastric Motility to Gastric Dysmotility. Int. J. Mol. Sci. 2021, 22, 9990. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, B.; Li, W.; He, Y.; Tang, X.; Zhang, T.; Zhong, Z.; Pan, Q.; Zhang, Y. Effects of repeated drug administration on behaviors in normal mice and fluoxetine efficacy in chronic unpredictable mild stress mice. Biochem. Biophys. Res. Commun. 2022, 615, 36–42. [Google Scholar] [CrossRef]
- Hoggatt, A.F.; Hoggatt, J.; Honerlaw, M.; Pelus, L.M. A Spoonful of Sugar Helps the Medicine Go Down: A Novel Technique to Improve Oral Gavage in Mice. J. Am. Assoc. Lab. Anim. Sci. 2010, 49, 329–334. [Google Scholar]
- McDonnell-Dowling, K.; Kleefeld, S.; Kelly, J.P. Consequences of Oral Gavage during Gestation and Lactation on Rat Dams and the Neurodevelopment and Behavior of Their Offspring. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 79. [Google Scholar]
- Gil, M.C.; Aguirre, J.A.; Lemoine, A.P.; Segura, E.T.; Barontini, M.; Armando, I. Influence of Age on Stress Responses to Metabolic Cage Housing in Rats. Cell. Mol. Neurobiol. 1999, 19, 625–633. [Google Scholar] [CrossRef]
- Labanski, A.; Langhorst, J.; Engler, H.; Elsenbruch, S. Stress and the Brain-Gut Axis in Functional and Chronic-Inflammatory Gastrointestinal Diseases: A Transdisciplinary Challenge. Psychoneuroendocrinology 2020, 111, 104501. [Google Scholar] [CrossRef]
- Bagues, A.; Lopez-Tofiño, Y.; Galvez-Robleño, C.; Abalo, R. Effects of Two Different Acute and Subchronic Stressors on Gastrointestinal Transit in the Rat: A Radiographic Analysis. Neurogastroenterol. Motil. 2021, 33, e14232. [Google Scholar] [CrossRef]
- Gálvez-Robleño, C.; López-Tofiño, Y.; López-Gómez, L.; Bagüés, A.; Soto-Montenegro, M.L.; Abalo, R. Radiographic Assessment of the Impact of Sex and the Circadian Rhythm-Dependent Behaviour on Gastrointestinal Transit in the Rat. Lab. Anim. 2023, 57, 270–282. [Google Scholar] [CrossRef]
- Chey, W.D.; Hashash, J.G.; Manning, L.; Chang, L. AGA Clinical Practice Update on the Role of Diet in Irritable Bowel Syndrome: Expert Review. Gastroenterology 2022, 162, 1737–1745.e5. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Instant Cascara Beverage (10 mg/mL) |
|---|---|
| Carbohydrates (mg/mL) | 4.748 |
| Total fiber (mg/mL) | 1.832 |
| Lipids (mg/mL) | 0.058 |
| Protein (mg/mL) | 0.625 |
| Magnesium (mg/mL) | 2.084 × 10−3 |
| Sodium (mg/mL) | 26.658 × 10−3 |
| Potassium (mg/mL) | 228.4 × 10−3 |
| Calcium (mg/mL) | 5.478 × 10−3 |
| Caffeine (mg/mL) | 0.139 |
| Phenolic content (mg eq. Cga/mL) | 0.89 |
| Chlorogenic acids (mg/mL) | 1.07–1.26 |
| Melanoidins (mg/mL) | 1.5 |
| Males—Control | Males—IC | Females—Control | Females—IC | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Initial (g) | Final (g) | Initial (g) | Final (g) | Initial (g) | Final (g) | Initial (g) | Final (g) | ||
| LM | PA | 0.40 ± 0.07 | 0.35 ± 0.06 | 0.38 ± 0.06 | 0.31 ± 0.05 | 0.41 ± 0.05 | 0.33 ± 0.05 | 0.44 ± 0.05 | 0.36 ± 0.05 |
| TA | 0.24 ± 0.04 | 0.21 ± 0.05 | 0.27 ± 0.05 | 0.18 ± 0.03 | 0.18 ± 0.03 | 0.19 ± 0.05 | 0.22 ± 0.03 | 0.19 ± 0.03 | |
| CM | PA | 1.73 ± 0.21 | 1.53 ± 0.19 | 1.47 ± 0.18 | 1.48 ± 0.18 | 0.96 ± 0.13 ## | 0.96 ± 0.10 # | 1.14 ± 0.09 | 1.31 ± 0.08 |
| TA | 0.75 ± 0.11 | 0.70 ± 0.12 | 0.80 ± 0.11 | 0.63 ± 0.11 | 0.55 ± 0.09 # | 0.37 ± 0.05 | 0.74 ± 0.09 | 0.54 ± 0.06 | |
| Antibody | Parameter (Unit) | Males—Control | Males—IC | Females—Control | Females—IC |
|---|---|---|---|---|---|
| HuC/D | Ganglionic area (%) | 10.90 ± 1.21 | 10.59 ± 1.28 | 10.95 ± 0.59 | 11.05 ± 1.28 |
| Ganglionic size (μm2) | 25,716 ± 2690 | 22,345 ± 2750 | 20,332 ± 3277 | 25,126 ± 3199 | |
| Neurons/ganglion | 54.06 ± 4.933 | 47 ± 4.77 | 41.91 ± 9.12 | 49.77 ± 6.431 | |
| Total neuronal density (neurons/mm2) | 231.40 ±19.46 | 212 ± 17.64 | 217.40 ± 23.49 | 219.90 ± 28.79 | |
| Packing density (intraganglionic neurons/ganglionic area) | 2156 ± 226.1 | 2147 ± 134.7 | 2000 ± 143.1 | 1976 ± 45.15 | |
| Extraganglionic neuronal density (extraganglionic neurons/serosal area) | 3.75 ± 0.78 | 4.28 ± 0.89 | 4.81 ± 0.35 | 3.03 ± 1.026 | |
| nNOS | Inhibitory neurons (%) | 18.67 ± 2.24 | 22.13 ± 2.36 | 18.93 ± 2.02 | 22.25 ± 4.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallego-Barceló, P.; Benítez-Álvarez, D.; Bagues, A.; Silván-Ros, B.; Montalbán-Rodríguez, A.; López-Gómez, L.; Vera, G.; del Castillo, M.D.; Uranga, J.A.; Abalo, R. Ex Vivo Study of Colon Health, Contractility and Innervation in Male and Female Rats after Regular Exposure to Instant Cascara Beverage. Foods 2024, 13, 2474. https://doi.org/10.3390/foods13162474
Gallego-Barceló P, Benítez-Álvarez D, Bagues A, Silván-Ros B, Montalbán-Rodríguez A, López-Gómez L, Vera G, del Castillo MD, Uranga JA, Abalo R. Ex Vivo Study of Colon Health, Contractility and Innervation in Male and Female Rats after Regular Exposure to Instant Cascara Beverage. Foods. 2024; 13(16):2474. https://doi.org/10.3390/foods13162474
Chicago/Turabian StyleGallego-Barceló, Paula, David Benítez-Álvarez, Ana Bagues, Blanca Silván-Ros, Alba Montalbán-Rodríguez, Laura López-Gómez, Gema Vera, María Dolores del Castillo, José A. Uranga, and Raquel Abalo. 2024. "Ex Vivo Study of Colon Health, Contractility and Innervation in Male and Female Rats after Regular Exposure to Instant Cascara Beverage" Foods 13, no. 16: 2474. https://doi.org/10.3390/foods13162474
APA StyleGallego-Barceló, P., Benítez-Álvarez, D., Bagues, A., Silván-Ros, B., Montalbán-Rodríguez, A., López-Gómez, L., Vera, G., del Castillo, M. D., Uranga, J. A., & Abalo, R. (2024). Ex Vivo Study of Colon Health, Contractility and Innervation in Male and Female Rats after Regular Exposure to Instant Cascara Beverage. Foods, 13(16), 2474. https://doi.org/10.3390/foods13162474