
Sunflower Meal Valorization through Enzyme-Aided Fractionation and the Production of Emerging Prebiotics
 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Compositional Analysis of Sunflower Meal
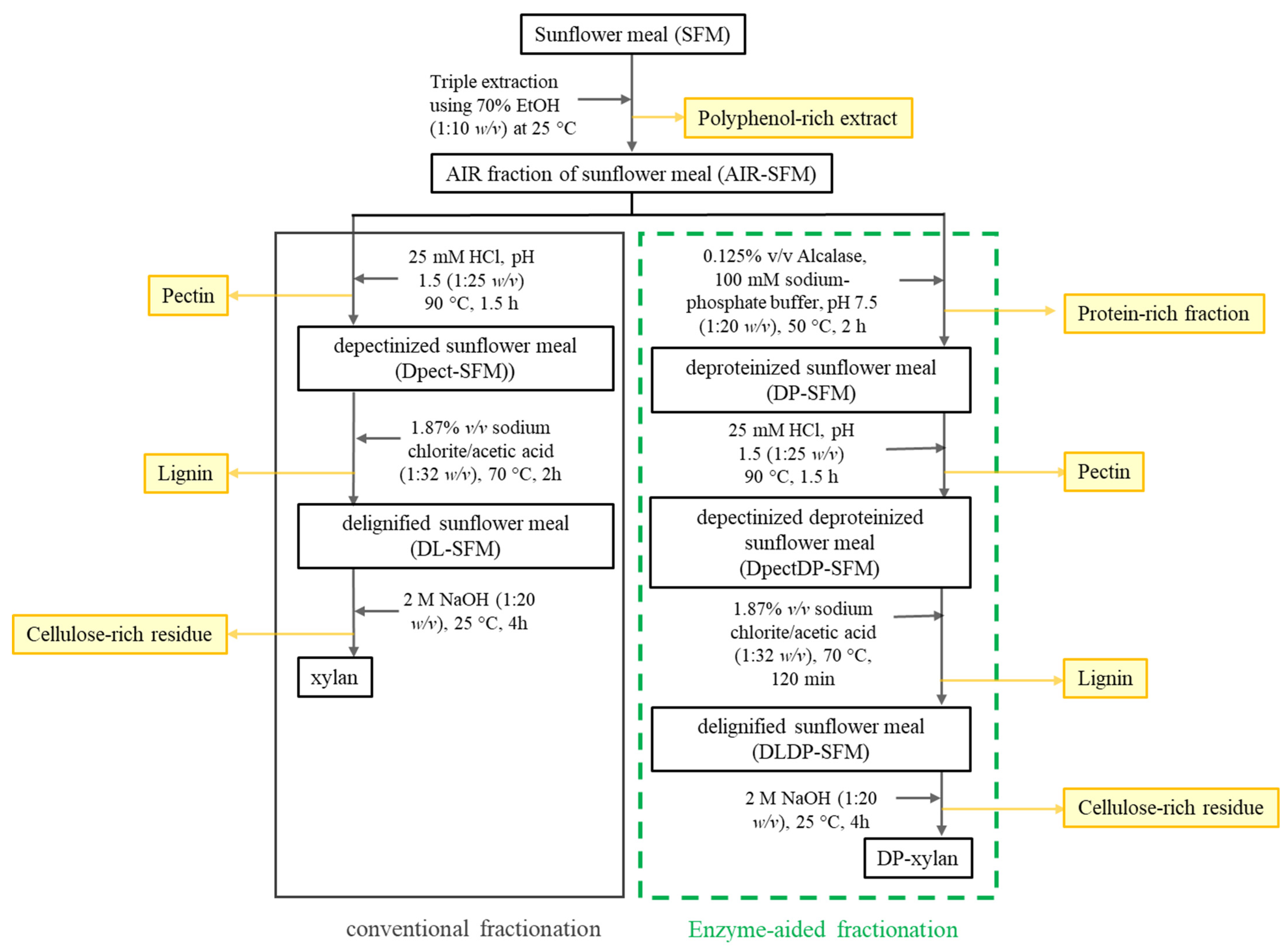
2.3. Sunflower Meal Fractionation
2.3.1. Method of Conventional Sunflower Meal Fractionation
2.3.2. Enzyme-Aided Method of Sunflower Meal Fractionation
2.4. Obtained Xylan Analysis
2.4.1. Fourier-Transform Infrared Spectroscopy (FTIR) Analysis
2.4.2. Determination of Monomeric Composition
2.4.3. Determination of the Molecular Mass (Mw) Distribution by High-Performance Size-Exclusion Chromatography with a Evaporative Light Scattering Detector (HPSEC-ELSD)
2.5. Enzymatic Production of Xylo-Oligosaccharides (XOS)
2.6. XOS Analysis
2.6.1. Determination of Total Reducing Sugars
2.6.2. High-Performance Liquid Chromatography (HPLC) Analysis of Reaction Mixture
2.6.3. Matrix-Assisted Laser Desorption/Ionization Time-Of-Flight Mass Spectrometry Analysis (MALDI-TOF-MS)
2.6.4. Statistical Analysis
3. Results and Discussion
3.1. Chemical Characterization of Sunflower Meal
3.2. Sunflower Meal Fractionation and Xylan Extraction
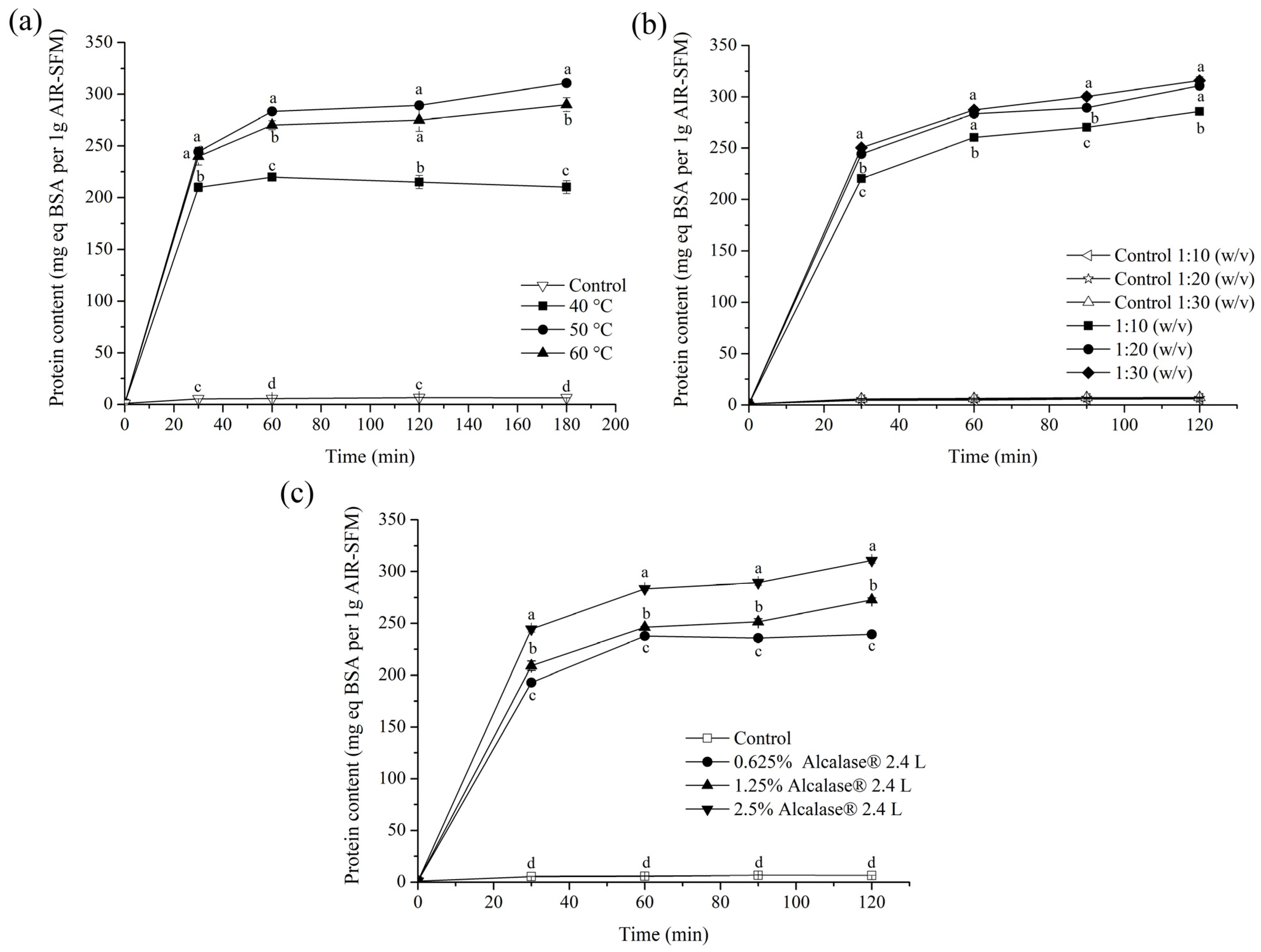
3.3. Optimization of Protein Extraction from AIR-SFM Using Alcalase® 2.4 L
3.4. Enzyme-Aided Fractionation of Sunflower Meal
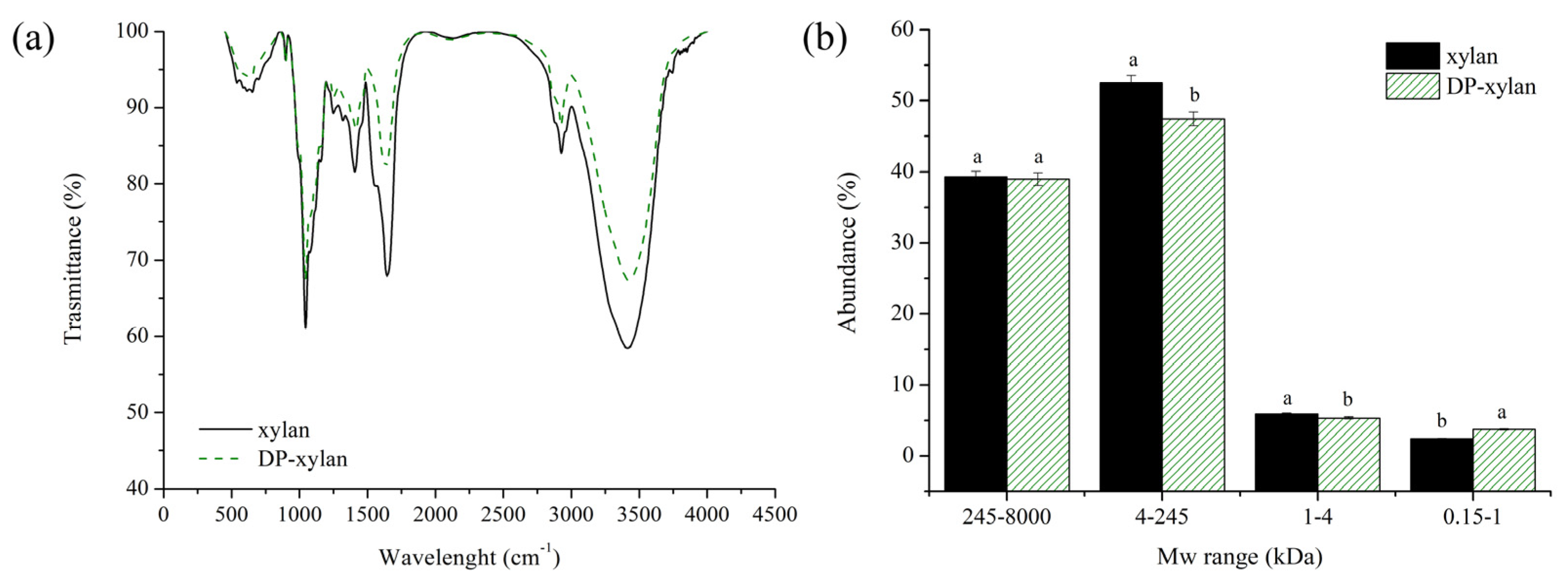
3.5. Characterization and Comparison of Obtained Xylans
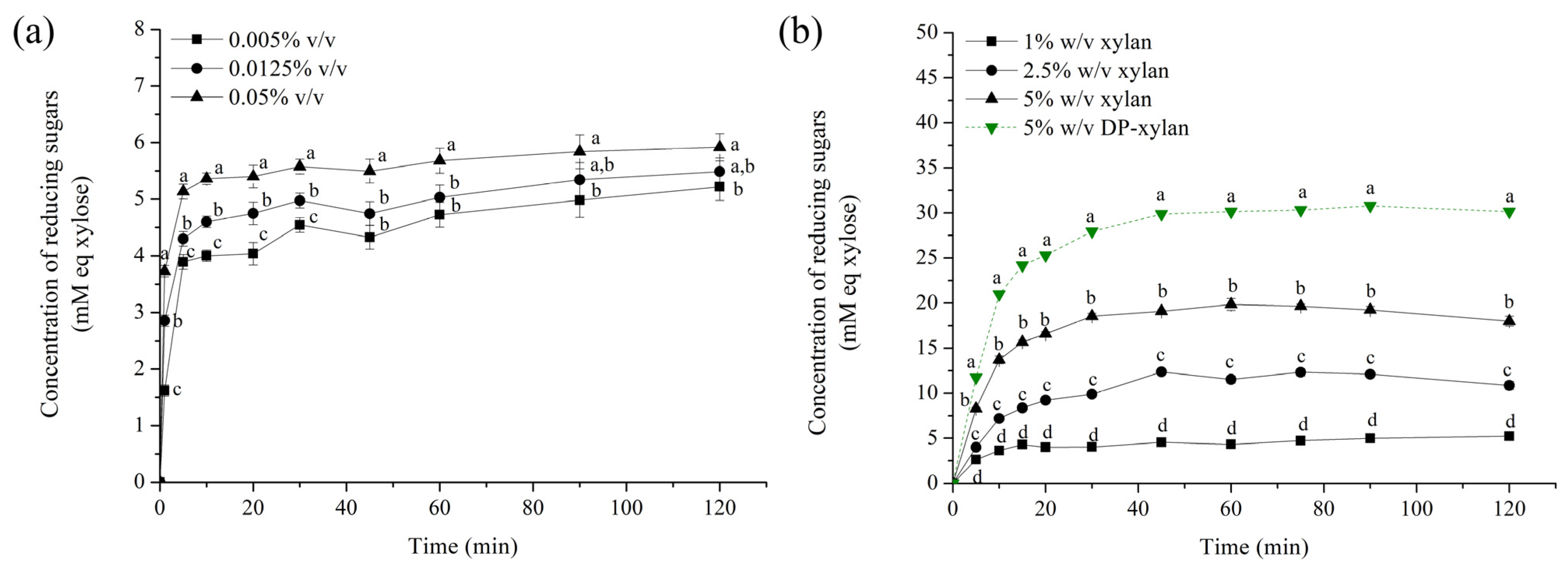
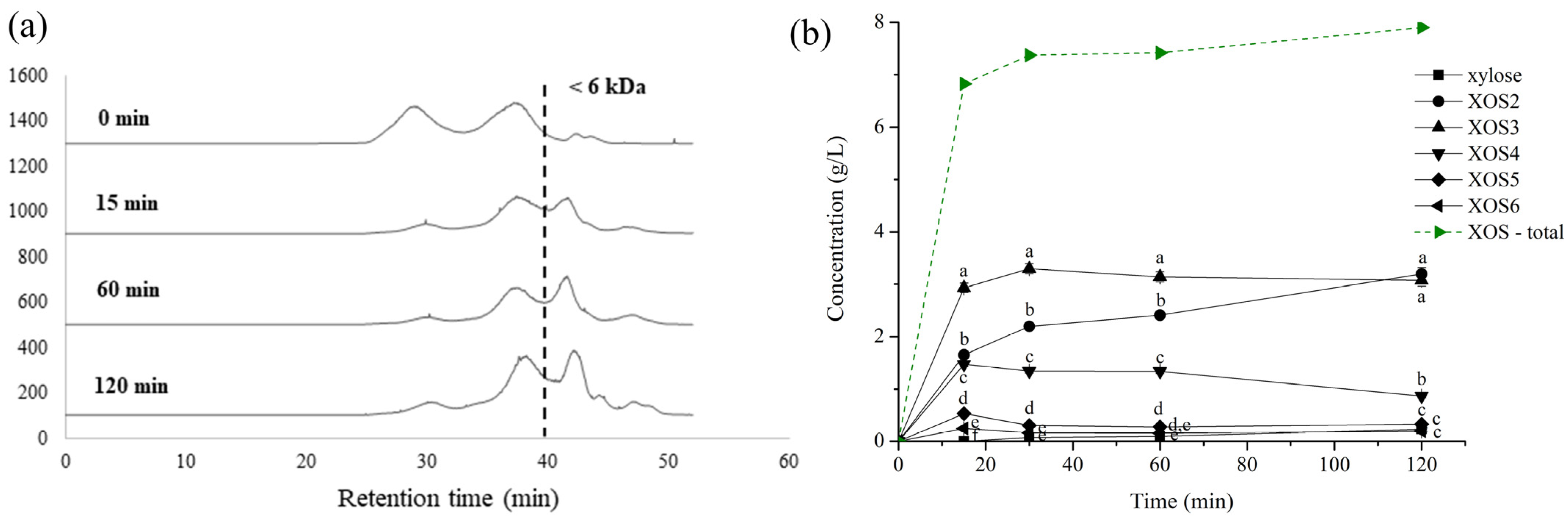
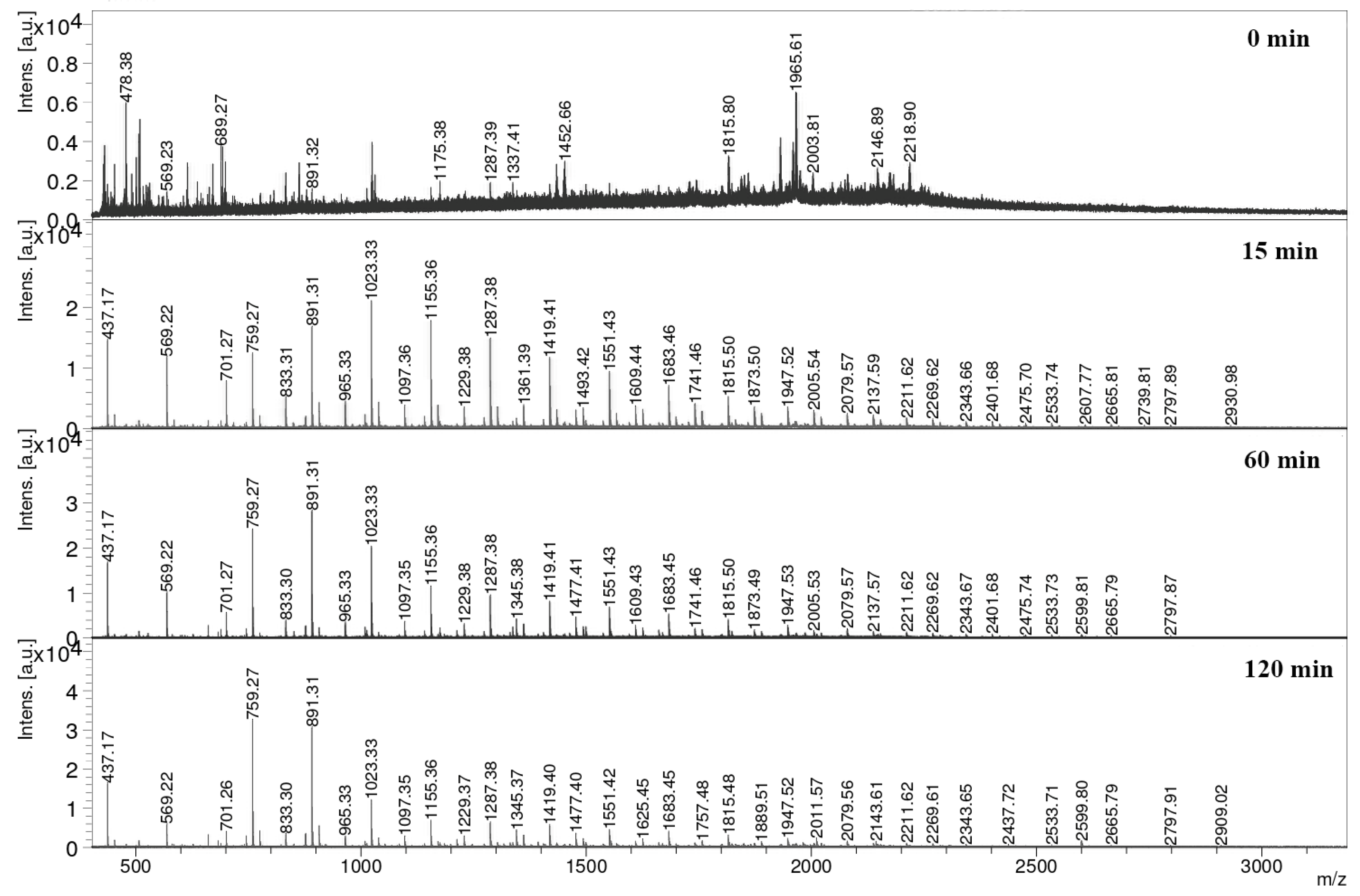
3.6. Hydrolysis of Obtained Xylans toward Xylo-Oligosaccharides Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adeleke, B.S.; Babalola, O.O. Oilseed crop sunflower (Helianthus annuus) as a source of food: Nutritional and health benefits. Food Sci. Nutr. 2020, 8, 4666–4684. [Google Scholar] [CrossRef] [PubMed]
- Lomascolo, A.; Uzan-Boukhris, E.; Sigoillot, J.-C.; Fine, F. Rapeseed and sunflower meal: A review on biotechnology status and challenges. Appl. Microbiol. Biotechnol. 2012, 95, 1105–1114. [Google Scholar] [CrossRef]
- Khurana, S.; Singh, R. Sunflower (Helianthus annuus) Seed. In Oilseeds: Health Attributes and Food Applications; Tanwar, B., Goyal, A., Eds.; Springer: Singapore, 2021; pp. 123–143. [Google Scholar]
- Khan, S.; Choudhary, S.; Pandey, A.; Khan, M.K.; Thomas, G. Sunflower oil: Efficient oil source for human consumption. Emergent Life Sci. Res. 2015, 1, 1–3. [Google Scholar]
- Kaur, R.; Ghoshal, G. Sunflower protein isolates-composition, extraction and functional properties. Adv. Colloid Interface Sci. 2022, 306, 102725. [Google Scholar] [CrossRef]
- Hadidi, M.; Aghababaei, F.; McClements, D.J. Sunflower meal/cake as a sustainable protein source for global food demand: Towards a zero-hunger world. Food Hydrocoll. 2024, 147, 109329. [Google Scholar] [CrossRef]
- de Oliveira Filho, J.G.; Egea, M.B. Sunflower seed byproduct and its fractions for food application: An attempt to improve the sustainability of the oil process. J. Food Sci. 2021, 86, 1497–1510. [Google Scholar] [CrossRef]
- Vidosavljević, S.; Bojanić, N.; Ilić, P.; Rakić, D.; Đuragić, O.; Banjac, V.; Fišteš, A. Optimization of Grinding Process of Sunflower Meal for Obtaining Protein-Enriched Fractions. Processes 2022, 10, 2704. [Google Scholar] [CrossRef]
- Rama Rao, S.V.; Raju, M.V.L.N.; Panda, A.K.; Reddy, M.R. Sunflower seed meal as a substitute for soybean meal in commercial broiler chicken diets. Br. Poult. Sci. 2006, 47, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Zardo, I.; de Espíndola Sobczyk, A.; Marczak, L.D.F.; Sarkis, J. Optimization of Ultrasound Assisted Extraction of Phenolic Compounds from Sunflower Seed Cake Using Response Surface Methodology. Waste Biomass Valorization 2019, 10, 33–44. [Google Scholar] [CrossRef]
- Kachrimanidou, V.; Kopsahelis, N.; Alexandri, M.; Strati, A.; Gardeli, C.; Papanikolaou, S.; Komaitis, M.; Kookos, I.K.; Koutinas, A.A. Integrated sunflower-based biorefinery for the production of antioxidants, protein isolate and poly(3-hydroxybutyrate). Ind. Crops Prod. 2015, 71, 106–113. [Google Scholar] [CrossRef]
- Dabbour, M.; Jiang, H.; Mintah, B.K.; Wahia, H.; He, R. Ultrasonic-assisted protein extraction from sunflower meal: Kinetic modeling, functional, and structural traits. Innov. Food Sci. Emerg. Technol. 2021, 74, 102824. [Google Scholar] [CrossRef]
- González-Pérez, S.; Vereijken, J.M. Sunflower proteins: Overview of their physicochemical, structural and functional properties. J. Sci. Food Agric. 2007, 87, 2173–2191. [Google Scholar] [CrossRef]
- Zaky, A.A.; Hussein, A.S.; Mostafa, S.; Abd El-Aty, A.M. Impact of Sunflower Meal Protein Isolate Supplementation on Pasta Quality. Separations 2022, 9, 429. [Google Scholar] [CrossRef]
- Shchekoldina, T.; Aider, M. Production of low chlorogenic and caffeic acid containing sunflower meal protein isolate and its use in functional wheat bread making. J. Food Sci. Technol. 2014, 51, 2331–2343. [Google Scholar] [CrossRef]
- Liu, H.-M.; Liu, X.-Y.; Yan, Y.-Y.; Gao, J.-H.; Qin, Z.; Wang, X.-D. Structural properties and antioxidant activities of polysaccharides isolated from sunflower meal after oil extraction. Arab. J. Chem. 2021, 14, 103420. [Google Scholar] [CrossRef]
- Sporck, D.; Reinoso, F.A.M.; Rencoret, J.; Gutiérrez, A.; del Rio, J.C.; Ferraz, A.; Milagres, A.M.F. Xylan extraction from pretreated sugarcane bagasse using alkaline and enzymatic approaches. Biotechnol. Biofuels 2017, 10, 296. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Almagro, N.; Vendrell-Calatayud, M.; Méndez-Albiñana, P.; Moreno, R.; Cano, M.P.; Villamiel, M. Extraction optimization and structural characterization of pectin from persimmon fruit (Diospyros kaki Thunb. var. Rojo brillante). Carbohydr. Polym. 2021, 272, 118411. [Google Scholar] [CrossRef] [PubMed]
- Riyamol; Gada Chengaiyan, J.; Rana, S.S.; Ahmad, F.; Haque, S.; Capanoglu, E. Recent Advances in the Extraction of Pectin from Various Sources and Industrial Applications. ACS Omega 2023, 8, 46309–46324. [Google Scholar] [CrossRef] [PubMed]
- Lehuedé, L.; Henríquez, C.; Carú, C.; Córdova, A.; Mendonça, R.T.; Salazar, O. Xylan extraction from hardwoods by alkaline pretreatment for xylooligosaccharide production: A detailed fractionation analysis. Carbohydr. Polym. 2023, 302, 120381. [Google Scholar] [CrossRef]
- Ngene, G.I.; Roux, J.-C.; Lachenal, D. Xylan extraction strategies and the performance of the resulting high purity cellulose fiber to swelling and dissolution. Carbohydr. Polym. Technol. Appl. 2022, 3, 100196. [Google Scholar] [CrossRef]
- Puițel, A.C.; Suditu, G.D.; Drăgoi, E.N.; Danu, M.; Ailiesei, G.-L.; Balan, C.D.; Chicet, D.-L.; Nechita, M.T. Optimization of Alkaline Extraction of Xylan-Based Hemicelluloses from Wheat Straws: Effects of Microwave, Ultrasound, and Freeze–Thaw Cycles. Polymers 2023, 15, 1038. [Google Scholar] [CrossRef]
- Jin, X.; Hu, Z.; Wu, S.; Song, T.; Yue, F.; Xiang, Z. Promoting the material properties of xylan-type hemicelluloses from the extraction step. Carbohydr. Polym. 2019, 215, 235–245. [Google Scholar] [CrossRef]
- Rowley, J.; Decker, S.R.; Michener, W.; Black, S. Efficient extraction of xylan from delignified corn stover using dimethyl sulfoxide. 3 Biotech 2013, 3, 433–438. [Google Scholar] [CrossRef]
- Adetunji, L.R.; Adekunle, A.; Orsat, V.; Raghavan, V. Advances in the pectin production process using novel extraction techniques: A review. Food Hydrocoll. 2017, 62, 239–250. [Google Scholar] [CrossRef]
- Sabater, C.; Corzo, N.; Olano, A.; Montilla, A. Enzymatic extraction of pectin from artichoke (Cynara scolymus L.) by-products using Celluclast(®)1.5L. Carbohydr. Polym. 2018, 190, 43–49. [Google Scholar] [CrossRef]
- Sabater, C.; Sabater, V.; Olano, A.; Montilla, A.; Corzo, N. Ultrasound-assisted extraction of pectin from artichoke by-products. An artificial neural network approach to pectin characterisation. Food Hydrocoll. 2020, 98, 105238. [Google Scholar] [CrossRef]
- Ma, X.; Yu, J.; Jing, J.; Zhao, Q.; Ren, L.; Hu, Z. Optimization of sunflower head pectin extraction by ammonium oxalate and the effect of drying conditions on properties. Sci. Rep. 2021, 11, 10616. [Google Scholar] [CrossRef]
- Muñoz-Almagro, N.; Rico-Rodriguez, F.; Wilde, P.J.; Montilla, A.; Villamiel, M. Structural and technological characterization of pectin extracted with sodium citrate and nitric acid from sunflower heads. Electrophoresis 2018, 39, 1984–1992. [Google Scholar] [CrossRef]
- Ezzati, S.; Ayaseh, A.; Ghanbarzadeh, B.; Heshmati, M.K. Pectin from sunflower by-product: Optimization of ultrasound-assisted extraction, characterization, and functional analysis. Int. J. Biol. Macromol. 2020, 165, 776–786. [Google Scholar] [CrossRef]
- Zhang, Q.; Cheng, L.; Ma, X.; Zhou, X.; Xu, Y. Revalorization of sunflower stalk pith as feedstock for the coproduction of pectin and glucose using a two-step dilute acid pretreatment process. Biotechnol. Biofuels 2021, 14, 194. [Google Scholar] [CrossRef] [PubMed]
- Valladares-Diestra, K.K.; de Souza Vandenberghe, L.P.; Vieira, S.; Goyzueta-Mamani, L.D.; de Mattos, P.B.G.; Manzoki, M.C.; Soccol, V.T.; Soccol, C.R. The Potential of Xylooligosaccharides as Prebiotics and Their Sustainable Production from Agro-Industrial by-Products. Foods 2023, 12, 2681. [Google Scholar] [CrossRef]
- Palaniappan, A.; Antony, U.; Emmambux, M.N. Current status of xylooligosaccharides: Production, characterization, health benefits and food application. Trends Food Sci. Technol. 2021, 111, 506–519. [Google Scholar] [CrossRef]
- Santibáñez, L.; Henríquez, C.; Corro-Tejeda, R.; Bernal, S.; Armijo, B.; Salazar, O. Xylooligosaccharides from lignocellulosic biomass: A comprehensive review. Carbohydr. Polym. 2021, 251, 117118. [Google Scholar] [CrossRef]
- Cebin, A.V.; Ralet, M.-C.; Vigouroux, J.; Karača, S.; Martinić, A.; Komes, D.; Bonnin, E. Valorisation of walnut shell and pea pod as novel sources for the production of xylooligosaccharides. Carbohydr. Polym. 2021, 263, 117932. [Google Scholar] [CrossRef] [PubMed]
- Mertens, D. Critical conditions in determining detergent fibers. In Proceedings of the Forage Analysis Workshop, Denver, CO, USA, 16–17 September 1992; pp. C1–C8. [Google Scholar]
- Akusuand, O.M.; Kiin-Kabari, D.B. Effect of Frozen Storage on the Chemical Stability, Functional and Sensory Properties of ‘Ogbono’ (Irvingiagarbonensis var. excelsa) and ‘Egusi’ (Citrullus vulgaris schrad) Seed Flours. Am. J. Food Sci. Technol. 2016, 4, 20–24. [Google Scholar]
- Katić, K.; Banjanac, K.; Simović, M.; Ćorović, M.; Milivojević, A.; Marinković, A.; Bezbradica, D. Development of protease nanobiocatalysts and their application in hydrolysis of sunflower meal protein isolate. Int. J. Food Sci. Technol. 2021, 56, 4287–4297. [Google Scholar] [CrossRef]
- Petrov Ivanković, A.; Milivojević, A.; Ćorović, M.; Simović, M.; Banjanac, K.; Jansen, P.; Vukoičić, A.; van den Bogaard, E.; Bezbradica, D. In vitro evaluation of enzymatically derived blackcurrant extract as prebiotic cosmetic ingredient: Extraction conditions optimization and effect on cutaneous microbiota representatives. Chem. Biol. Technol. Agric. 2023, 10, 125. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Y.; Chen, F.; Zheng, F. Comparison of determination of sugar-PMP derivatives by two different stationary phases and two HPLC detectors: C18 vs. amide columns and DAD vs. ELSD. J. Food Compos. Anal. 2021, 96, 103715. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, J.; Zhang, L.; Zhang, W.; Zhang, Y. Determination of 1-phenyl-3-methyl-5-pyrazolone-labeled carbohydrates by liquid chromatography and micellar electrokinetic chromatography. J. Chromatogr. B 2003, 793, 159–165. [Google Scholar] [CrossRef]
- Alagawany, M.; Farag, M.R.; El-Hack, M.E.A.; Dhama, K. The Practical Application of Sunflower Meal in Poultry Nutrition. Adv. Anim. Vet. Sci. 2015, 3, 634–648. [Google Scholar] [CrossRef]
- Lannuzel, C.; Smith, A.; Mary, A.L.; Della Pia, E.A.; Kabel, M.A.; de Vries, S. Improving fiber utilization from rapeseed and sunflower seed meals to substitute soybean meal in pig and chicken diets: A review. Anim. Feed. Sci. Technol. 2022, 285, 115213. [Google Scholar] [CrossRef]
- Xiao, B.; Sun, X.F.; Sun, R. Chemical, structural, and thermal characterizations of alkali-soluble lignins and hemicelluloses, and cellulose from maize stems, rye straw, and rice straw. Polym. Degrad. Stab. 2001, 74, 307–319. [Google Scholar] [CrossRef]
- Cruz-Solis, I.; Ibarra-Herrera, C.C.; Rocha-Pizaña, M.d.R.; Luna-Vital, D. Alkaline Extraction–Isoelectric Precipitation of Plant Proteins. In Green Protein Processing Technologies from Plants: Novel Extraction and Purification Methods for Product Development; Hernández-Álvarez, A.J., Mondor, M., Nosworthy, M.G., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 1–29. [Google Scholar]
- Milutinović, M.; Dimitrijević-Branković, S.; Rajilić-Stojanović, M. Plant Extracts Rich in Polyphenols as Potent Modulators in the Growth of Probiotic and Pathogenic Intestinal Microorganisms. Front. Nutr. 2021, 8, 688843. [Google Scholar] [CrossRef] [PubMed]
- Streimikyte, P.; Viskelis, P.; Viskelis, J. Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications. Int. J. Mol. Sci. 2022, 23, 2359. [Google Scholar] [CrossRef] [PubMed]
- Ashori, A.; Ornelas, M.; Sheshmani, S.; Cordeiro, N. Influence of mild alkaline treatment on the cellulosic surfaces active sites. Carbohydr. Polym. 2012, 88, 1293–1298. [Google Scholar] [CrossRef]
- Awuor, O.L.; Kirwa, M.E.; Betty, M.; Jackim, M.F. Optimization of Alcalase hydrolysis conditions for production of Dagaa (Rastrineobola argentea) Protein hydrolysate with antioxidative properties. Ind. Chem. 2017, 3, 122. [Google Scholar] [CrossRef]
- Vovk, H.; Karnpakdee, K.; Ludwig, R.; Nosenko, T. Enzymatic Pretreatment of Plant Cells for Oil Extraction. Food Technol. Biotechnol. 2023, 61, 160–178. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.M.d. Study on the Structure and Properties of Xylan Extracted from Eucalyptus, Sugarcane Bagasse and Sugarcane Straw. Ph.D. Thesis, KTH Royal Institute of Technology, Stockholm, Sweden, 2015. [Google Scholar]
- Kačuráková, M.; Belton, P.S.; Wilson, R.H.; Hirsch, J.; Ebringerová, A. Hydration properties of xylan-type structures: An FTIR study of xylooligosaccharides. J. Sci. Food Agric. 1998, 77, 38–44. [Google Scholar] [CrossRef]
- Sun, J.X.; Sun, X.F.; Sun, R.C.; Su, Y.Q. Fractional extraction and structural characterization of sugarcane bagasse hemicelluloses. Carbohydr. Polym. 2004, 56, 195–204. [Google Scholar] [CrossRef]
- Álvarez, C.; González, A.; Ballesteros, I.; Gullón, B.; Negro, M.J. In Vitro Assessment of the Prebiotic Potential of Xylooligosaccharides from Barley Straw. Foods 2023, 12, 83. [Google Scholar] [CrossRef]
- Bian, J.; Peng, F.; Peng, X.-P.; Peng, P.; Xu, F.; Sun, R.-C. Structural features and antioxidant activity of xylooligosaccharides enzymatically produced from sugarcane bagasse. Bioresour. Technol. 2013, 127, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.; Yan, Q.; Jiang, Z.; Fan, G.; Shi, B. Production of xylooligosaccharides from the steam explosion liquor of corncobs coupled with enzymatic hydrolysis using a thermostable xylanase. Bioresour. Technol. 2010, 101, 7679–7682. [Google Scholar] [CrossRef] [PubMed]
- Valladares-Diestra, K.K.; Porto de Souza Vandenberghe, L.; Soccol, C.R. A biorefinery approach for enzymatic complex production for the synthesis of xylooligosaccharides from sugarcane bagasse. Bioresour. Technol. 2021, 333, 125174. [Google Scholar] [CrossRef] [PubMed]
- Babbar, N.; Dejonghe, W.; Sforza, S.; Elst, K. Enzymatic pectic oligosaccharides (POS) production from sugar beet pulp using response surface methodology. J. Food Sci. Technol. 2017, 54, 3707–3715. [Google Scholar] [CrossRef]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Concentration |
|---|---|
| Dry matter (%) | 91.5 ± 0.3 |
| Simple sugars (% DM) | 5.5 ± 0.3 |
| Hemicellulose (% DM) | 12.7 ± 0.9 |
| Cellulose (% DM) | 13.5 ± 0.1 |
| Lignin (% DM) | 6.8 ± 0.5 |
| Proteins (% DM) | 42.8 ± 0.1 |
| Fats (% DM) | 2.8 ± 0.0 |
| Ash (% DM) | 7.4 ± 0.1 |
| Component | Concentration | ||
|---|---|---|---|
| SFM | Xylan | DP-xylan | |
| Xylose | 21.8 ± 1.9 c | 53.7 ± 5.4 b | 63.4 ± 1.1 a |
| Arabinose | 21.1 ± 0.8 a | 11.2 ± 0.1 b | 9.7 ± 0.9 b |
| Rhamnose | 3.6 ± 0.2 a | 3.4 ± 0.3 a | 3.3 ± 0.1 a |
| Fructose | 2.4 ± 0.4 a | 1.1 ± 0.6 b | 1.7 ± 0.1 a,b |
| Galactose | 10.6 ± 0.4 a | 11.8 ± 7.3 a | 5.8 ± 0.6 a |
| Mannose | 3.9 ± 0.3 a | 2.0 ± 0.2 b | 0.5 ± 0.1 c |
| Glucose | 22.3 ± 2.9 a | 10.4 ± 1.1 b | 8.0 ± 0.2 b |
| Galacturonic acid | 14.4 ± 0.4 a | 6.5 ± 0.5 c | 7.7 ± 0.1 b |
| Characteristic | Xylan | DP-xylan |
|---|---|---|
| Quantity (g) | 6.7 ± 0.3 a | 5.8 ± 0.6 a |
| Dry matter (%) | 96.8 ± 1.1 a | 95.0 ± 2.2 a |
| Yield (%) | 7.1 ± 0.5 a | 6.1 ± 0.6 a |
| Recovery yield (%) | 56.0 ± 2.2 a | 47.8 ± 0.9 b |
| Carbohydrate content (%) | 52.5 ± 2.3 b | 92.2 ± 3.3 a |
| Protein content (%) | 25.3 ± 1.2 a | 7.1 ± 1.1 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simović, M.; Banjanac, K.; Veljković, M.; Nikolić, V.; López-Revenga, P.; Montilla, A.; Moreno, F.J.; Bezbradica, D. Sunflower Meal Valorization through Enzyme-Aided Fractionation and the Production of Emerging Prebiotics. Foods 2024, 13, 2506. https://doi.org/10.3390/foods13162506
Simović M, Banjanac K, Veljković M, Nikolić V, López-Revenga P, Montilla A, Moreno FJ, Bezbradica D. Sunflower Meal Valorization through Enzyme-Aided Fractionation and the Production of Emerging Prebiotics. Foods. 2024; 13(16):2506. https://doi.org/10.3390/foods13162506
Chicago/Turabian StyleSimović, Milica, Katarina Banjanac, Milica Veljković, Valentina Nikolić, Paula López-Revenga, Antonia Montilla, Francisco Javier Moreno, and Dejan Bezbradica. 2024. "Sunflower Meal Valorization through Enzyme-Aided Fractionation and the Production of Emerging Prebiotics" Foods 13, no. 16: 2506. https://doi.org/10.3390/foods13162506