
A Novel Approach Based on Real-Time PCR with High-Resolution Melting Analysis for the Simultaneous Identification of Staphylococcus aureus and Staphylococcus argenteus

 ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reference Strains and Bacterial Isolates
2.2. Bacterial Growth and DNA Extraction
2.3. Primers and Real-Time PCR-HRM Conditions
2.4. Limit of Detection and Efficiency of the Assay
3. Results
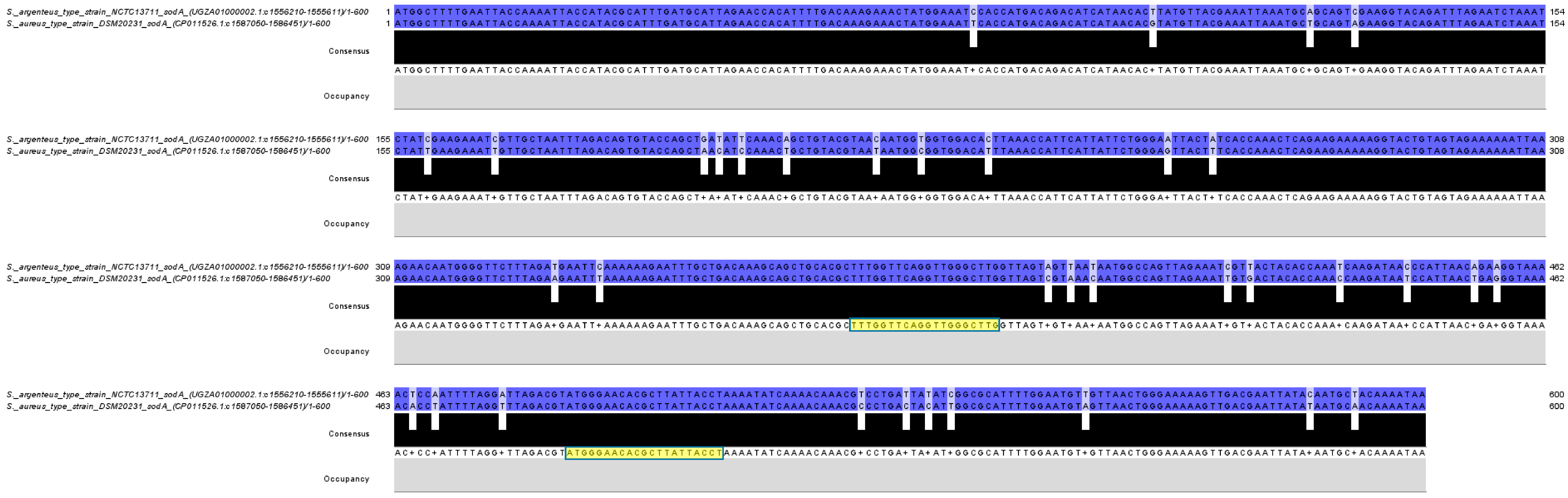
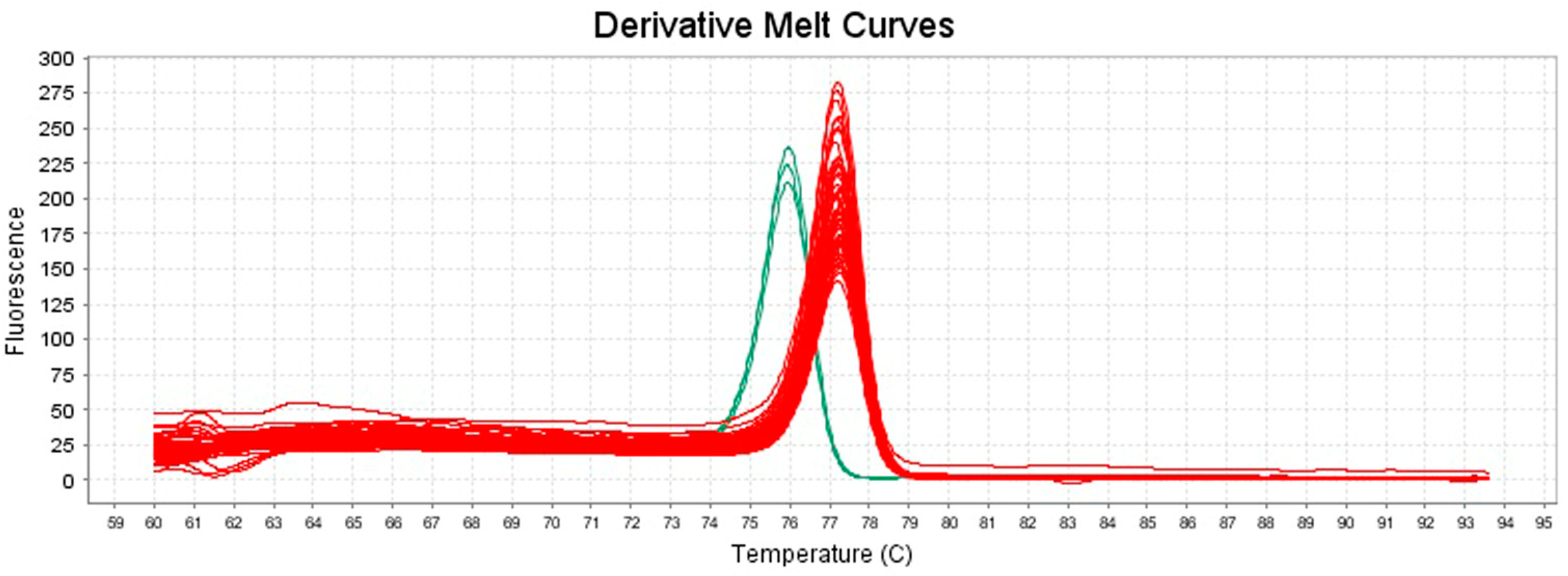
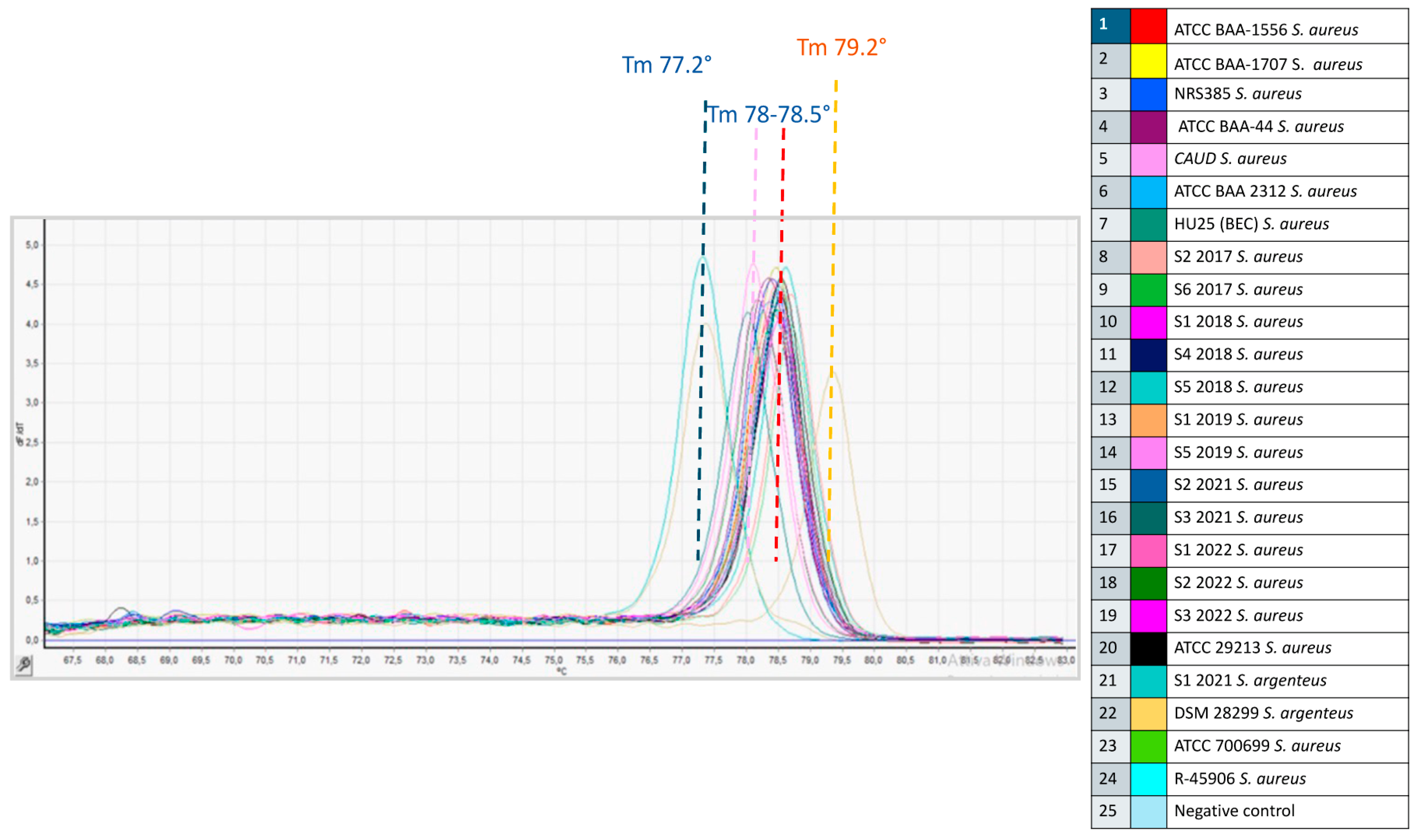
3.1. HRM Analysis of the Tested Isolates
3.2. Limit of Detection
3.3. Standard Curves and Efficiency
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Licitra, G. Etymologia: Staphylococcus. Emerg. Infect. Dis. 2013, 19, 1553. [Google Scholar] [CrossRef]
- Fusco, V.; Quero, G.M.; Morea, M.; Blaiotta, G.; Visconti, A. Rapid and reliable identification of Staphylococcus aureus harbouring the enterotoxin gene cluster (egc) and quantitative detection in raw milk by real time PCR. Int. J. Food Microbiol. 2011, 144, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Chieffi, D.; Fanelli, F.; Cho, G.S.; Schubert, J.; Blaiotta, G.; Franz, C.M.A.P.; Bania, J.; Fusco, V. Novel insights into the enterotoxigenic potential and genomic background of Staphylococcus aureus isolated from raw milk. Food Microbiol. 2020, 90, 103482. [Google Scholar] [CrossRef] [PubMed]
- Dicks, J.; Turnbull, J.D.; Russell, J.; Parkhill, J.; Alexander, S. Genome sequencing of a historic Staphylococcus aureus collection reveals new enterotoxin genes and sheds light on the evolution and genomic organization of this key virulence gene family. J. Bacteriol. 2021, 203, e00587-20. [Google Scholar] [CrossRef]
- Fanelli, F.; Chieffi, D.; Cho, G.S.; Schubert, J.; Mekhloufi, O.A.; Bania, J.; Franz, C.M.A.P.; Fusco, V. First genome-based characterisation and staphylococcal enterotoxin production ability of methicillin-susceptible and methicillin-resistant Staphylococcus aureus strains isolated from ready-to-eat foods in Algiers (Algeria). Toxins 2022, 14, 731. [Google Scholar] [CrossRef]
- Hennekinne, J.A.; Ostyn, A.; Guillier, F.; Herbin, S.; Prufer, A.L.; Dragacci, S. How should staphylococcal food poisoning outbreaks be characterized? Toxins 2010, 2, 2106–2116. [Google Scholar] [CrossRef]
- Seo, K.S.; Bohach, G.A. Staphylococcal Food Poisoning. In Pathogens and Toxins in Foods; Juneja, V.K., Sofos, J.N., Eds.; ASM Press: Washington, DC, USA, 2009. [Google Scholar] [CrossRef]
- Tong, S.Y.C.; Schaumburg, F.; Ellington, M.J.; Corander, J.; Pichon, B.; Leendertz, F.; Bentley, S.D.; Parkhill, J.; Holt, D.C.; Peterset, G.; et al. Novel staphylococcal species that form part of a Staphylococcus aureus-related complex: The non-pigmented Staphylococcus argenteus sp. nov. and the non-human primate-associated Staphylococcus schweitzeri sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 15–22. [Google Scholar] [CrossRef]
- Chew, K.L.; Octavia, S.; Lai, D.; Lin, R.T.P.; Teo, J.W.P. Staphylococcus singaporensis sp. nov., a new member of the Staphylococcus aureus complex, isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 2021, 71, 005067. [Google Scholar] [CrossRef]
- Schutte, A.H.J.; Strepis, N.; Zandijk, W.H.A.; Bexkens, M.L.; Bode, L.G.M.; Klaassen, C.H.W. Characterization of Staphylococcus roterodami sp. nov., a new species within the Staphylococcus aureus complex isolated from a human foot infection. Int. J. Syst. Evol. Microbiol. 2021, 71, 004996. [Google Scholar] [CrossRef]
- Monecke, S.; Schaumburg, F.; Shittu, A.O.; Schwarz, S.; Mühldorfer, K.; Brandt, C.; Braun, S.D.; Collatz, M.; Diezel, C.; Gawlik, D.; et al. Description of staphylococcal strains from straw-coloured fruit bat (Eidolon helvum) and diamond firetail (Stagonopleura guttata) and a review of their phylogenetic relationships to other staphylococci. Front. Cell. Infect. Microbiol. 2022, 12, 878137. [Google Scholar] [CrossRef] [PubMed]
- Chieffi, D.; Fanelli, F.; Fusco, V. Antimicrobial and biocide resistance in Staphylococcus aureus: Genomic features, decontamination strategies, and the role of S. aureus complex-related species, with a focus on ready-to-eat food and food-contact surfaces. Front. Food Sci. Technol. 2023, 3, 1165871. [Google Scholar] [CrossRef]
- Fusco, V.; Chieffi, D.; Fanelli, F.; Logrieco, A.F.; Cho, G.-S.; Kabisch, J.; Böhnlein, C.; Franz, C.M.A.P. Microbial quality and safety of milk and milk products in the 21st century. Comp. Rev. Food Sci. Food Saf. 2020, 19, 2013–2049. [Google Scholar] [CrossRef] [PubMed]
- Chantratita, N.; Wikraiphat, C.; Tandhavanant, S.; Wongsuvan, G.; Ariyaprasert, P.; Suntornsut, P.; Thaipadungpanit, J.; Teerawattanasook, N.; Jutrakul, Y.; Srisurat, N.; et al. Comparison of community-onset Staphylococcus argenteus and Staphylococcus aureus sepsis in Thailand: A prospective multicentre observational study. Clin. Microbiol. Infect. 2016, 22, e11–e19. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; You, B.; Tan, L.; Yu, S.; Li, H.; Bai, G.; Li, S.; Rao, X.; Xie, Z.; Shi, X.; et al. Clinical Staphylococcus argenteus develops to small colony variants to promote persistent infection. Front. Microbiol. 2018, 9, 1347. [Google Scholar] [CrossRef]
- Ohnishi, T.; Shinjoh, M.; Ohara, H.; Kawai, T.; Kamimaki, I.; Mizushima, R.; Kamada, K.; Itakura, Y.; Iguchi, S.; Uzawa, Y.; et al. Purulent lymphadenitis caused by Staphylococcus argenteus, representing the first Japanese case of Staphylococcus argenteus (multilocus sequence type 2250) infection in a 12-year-old boy. J. Infect. Chemother. 2018, 24, 925–927. [Google Scholar] [CrossRef]
- Hirai, J.; Suzuki, H.; Sakanashi, D.; Kuge, Y.; Kishino, T.; Asai, N.; Hagihara, M.; Mikamo, H. The first case report of community-acquired infective endocarditis due to sequence type 1223 Staphylococcus argenteus complicated with convexity subarachnoid hemorrhage. Infect. Drug Resist. 2022, 15, 4963–4970. [Google Scholar] [CrossRef]
- Indrawattana, N.; Pumipuntu, N.; Suriyakhun, N.; Jangsangthong, A.; Kulpeanprasit, S.; Chantratita, N.; Sookrung, N.; Chaicumpa, W.; Buranasinsup, S. Staphylococcus argenteus from rabbits in Thailand. MicrobiologyOpen 2019, 8, e00665. [Google Scholar] [CrossRef]
- Pumipuntu, N.; Tunyong, W.; Chantratita, N.; Diraphat, P.; Pumirat, P.; Sookrung, N.; Chaicumpa, W.; Indrawattana, N. Staphylococcus spp. associated with subclinical bovine mastitis in central and northeast provinces of Thailand. PeerJ 2019, 7, e6587. [Google Scholar] [CrossRef]
- Meijer, E.F.J.; van Renssen, A.; Maat, I.; van der Graaf-van Bloois, L.; Duim, B.; Broens, E.M. Canine Staphylococcus argenteus: Case report from The Netherlands. Pathogens 2022, 11, 153. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kubota, H.; Ono, H.K.; Kobayashi, M.; Murauchi, K.; Kato, R.; Hirai, A.; Sadamasu, K. Food poisoning outbreak in Tokyo, Japan caused by Staphylococcus argenteus. Int. J. Food Microbiol. 2017, 262, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, Y.; Umeda, K.; Yonogi, S.; Nakamura, H.; Yamamoto, K.; Kumeda, Y.; Kawatsu, K. Staphylococcal food poisoning caused by Staphylococcus argenteus harboring staphylococcal enterotoxin genes. Int. J. Food Microbiol. 2018, 265, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Cavaiuolo, M.; Lefebvre, D.; Mutel, I.; Vingadassalon, N.; Merda, D.; Hennekinne, J.A.; Nia, Y. First report of enterotoxigenic Staphylococcus argenteus as a foodborne pathogen. Int. J. Food Microbiol. 2023, 394, 110182. [Google Scholar] [CrossRef] [PubMed]
- Brakstad, O.G.; Aasbakk, K.; Maeland, J.A. Detection of Staphylococcus aureus by polymerase chain reaction amplification of the nuc gene. J. Clin. Microbiol. 1992, 30, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- Kaden, R.; Engstrand, L.; Rautelin, H.; Johansson, C. Which methods are appropriate for the detection of Staphylococcus argenteus and is it worthwhile to distinguish S. argenteus from S. aureus? Infect. Drug Resist. 2018, 11, 2335–2344. [Google Scholar] [CrossRef]
- Ata, Z.; Buyukcangaz, E. A 13-Plex PCR for high-resolution melting-based screening of clinically important Staphylococcus species in animals. J. Microbiol. Methods 2019, 164, 105673. [Google Scholar] [CrossRef]
- Tong, S.Y.; Giffard, P.M. Microbiological applications of high-resolution melting analysis. J. Clin. Microbiol. 2012, 50, 3418–3421. [Google Scholar] [CrossRef]
- Blaiotta, G.; Fusco, V.; von Eiff, C.; Villani, F.; Becker, K. Biotyping of enterotoxigenic Staphylococcus aureus by enterotoxin gene cluster (egc) polymorphism and spa typing analyses. Appl. Environ. Microbiol. 2006, 72, 6117–6123. [Google Scholar] [CrossRef]
- Mekhloufi, O.A.; Chieffi, D.; Hammoudi, A.; Bensefia, S.A.; Fanelli, F.; Fusco, V. Prevalence, Enterotoxigenic Potential and Antimicrobial Resistance of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus (MRSA) Isolated from Algerian Ready to Eat Foods. Toxins 2021, 13, 835. [Google Scholar] [CrossRef]
- Campanile, F.; Bongiorno, D.; Perez, M.; Mongelli, G.; Sessa, L.; Benvenuto, S.; Gona, F. AMCLI–S. aureus Survey Participants; Varaldo, P.E.; Stefani, S. Epidemiology of Staphylococcus aureus in Italy: First nationwide survey, 2012. J. Glob. Antimicrob. Resist. 2015, 3, 247–254. [Google Scholar] [CrossRef]
- Bongiorno, D.; Musso, N.; Caruso, G.; Lazzaro, L.M.; Caraci, F.; Stefani, S.; Campanile, F. Staphylococcus aureus ST228 and ST239 as models for expression studies of diverse markers during osteoblast infection and persistence. Microbiologyopen 2021, 10, e1178. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, D.G.; Fry, N.K. Molecular techniques for the detection and identification of new bacterial pathogens. J Infect. 2000, 40, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Zhang, D.F.; Zhi, X.Y.; Zhang, J.; Paoli, G.C.; Cui, Y.; Shi, C.; Shi, X. Preliminary comparative genomics revealed pathogenic potential and international spread of Staphylococcus argenteus. BMC Genom. 2017, 18, 808. [Google Scholar] [CrossRef]
- Doležel, J.; Bartos, J.; Voglmayr, H.; Greilhuber, J. Nuclear DNA content and genome size of trout and human. Cytometry. Part A J. Int. Soc. Anal. Cytol. 2003, 51, 127–129. [Google Scholar] [CrossRef]
- Schuster, D.; Rickmeyer, J.; Gajdiss, M.; Thye, T.; Lorenzen, S.; Reif, M.; Josten, M.; Szekat, C.; Melo, L.D.R.; Schmithausen, R.M.; et al. Differentiation of Staphylococcus argenteus (formerly: Staphylococcus aureus clonal complex 75) by mass spectrometry from S. aureus using the first strain isolated from a wild African great ape. Int. J. Med. Microbiol. 2017, 307, 57–63. [Google Scholar] [CrossRef]
- Chen, S.Y.; Lee, H.; Wang, X.M.; Lee, T.F.; Liao, C.H.; Teng, L.J.; Hsueh, P.R. High mortality impact of Staphylococcus argenteus on patients with community-onset staphylococcal bacteraemia. Int. J. Antimicrob. Agents 2018, 52, 747–753. [Google Scholar] [CrossRef]
- Becker, K.; Schaumburg, F.; Kearns, A.; Larsen, A.R.; Lindsay, J.A.; Skov, R.L.; Westh, H. Implications of identifying the recently defined members of the Staphylococcus aureus complex S. argenteus and S. schweitzeri: A position paper of members of the ESCMID Study Group for Staphylococci and Staphylococcal Diseases (ESGS). Clin. Microbiol. Infect. 2019, 25, 1064–1070. [Google Scholar] [CrossRef]
- Harshitha, R.; Arunraj, D.R. Real-time quantitative PCR: A tool for absolute and relative quantification. Biochem. Mol. Biol. Educ. 2021, 49, 800–812. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Polymerase Chain Reaction. Cold Spring Harb. Protoc. 2019, 158, 1154–1157. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Quero, G.M. Culture-dependent and -independent nucleic acid-based methods used in the microbial safety assessment of milk and dairy products. Compr. Rev. Food Sci. Food Saf. 2014, 13, 493–537. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Fanelli, F.; Chieffi, D. Authenticity of probiotic foods and dietary supplements: A pivotal issue to address. Crit. Rev. Food Sci. Nutr. 2022, 62, 6854–6871. [Google Scholar] [CrossRef]
- Landolt, P.; Stephan, R.; Scherrer, S. Development of a new High Resolution Melting (HRM) assay for identification and differentiation of Mycobacterium tuberculosis complex samples. Sci. Rep. 2019, 9, 1850. [Google Scholar] [CrossRef]
- Slany, M.; Vanerkova, M.; Nemcova, E.; Zaloudikova, B.; Ruzicka, F.; Freiberger, T. Differentiation of Staphylococcus spp. by high-resolution melting analysis. Can. J. Microbiol. 2010, 56, 1040–1049. [Google Scholar] [CrossRef]
- Poyart, C.; Quesne, G.; Boumaila, C.; Trieu-Cuot, P. Rapid and accurate species level identification of coagulase-negative staphylococci by using the sodA gene as a target. J. Clin. Microbiol. 2001, 39, 4296–4301. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions, and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Origin | Isolate | Origin | Isolate | Origin |
|---|---|---|---|---|---|
| 178P a | Raw milk | 363P a | Raw milk | SA23 b | Minced meat |
| 179P a | Raw milk | 372P a | Raw milk | SA33 b | Mashed potatoes |
| 180P a | Raw milk | 364P a | Raw milk | SA43 b | Rice |
| 181P a | Raw milk | 365P a | Raw milk | SA44 b | Salad |
| 182P a | Raw milk | 366P a | Raw milk | SA50 b | Salad |
| 183P a | Raw milk | 236P a | Raw milk | SA53 b | Minced meat |
| 184P a | Raw milk | 237P a | Raw milk | SA54 b | Beet salad |
| 185P a | Raw milk | 238P a | Raw milk | SA58 b | Meat |
| 186P a | Raw milk | 239P a | Raw milk | SA73 b | Salad |
| 187P a | Raw milk | 240P a | Raw milk | SA78 b | Lentil soup |
| 188P a | Raw milk | 241P a | Raw milk | SA83 b | Beans |
| 189P a | Raw milk | 242P a | Raw milk | SA84 b | Chicken |
| 190P a | Raw milk | 243P a | Raw milk | SA86 b | Sausages |
| 191P a | Raw milk | 244P a | Raw milk | SA87 b | Pastry |
| 192P a | Raw milk | 367P a | Raw milk | SA03 b | Salad |
| 193P a | Raw milk | 373P a | Raw milk | SA04 b | Meat |
| 194P a | Raw milk | 374P a | Raw milk | SA09 b | Pastry |
| 195P a | Raw milk | 200P a | Raw milk | SA29 b | Meat |
| 196P a | Raw milk | 201P a | Raw milk | SA31 b | Salad |
| 197P a | Raw milk | 211P a | Raw milk | SA72 b | Pastry |
| 198P a | Raw milk | 212P a | Raw milk | SA17 b | Turkey pieces |
| 199P a | Raw milk | 213P a | Raw milk | SA46 b | Braised beef |
| 356P a | Raw milk | 214P a | Raw milk | ATCC 27664 (FRI326) a | Chicken tetrazzini |
| 357P a | Raw milk | 234P a | Raw milk | BS4 a | “Napoli-type” salami |
| 358P a | Raw milk | SA01 b | Potato in sauce | DS18g a | “Napoli-type” salami |
| 359P a | Raw milk | SA06 b | Chicken | AS14g a | “Napoli-type” salami |
| 360P a | Raw milk | SA11 b | Couscous with meat | 382F a | Unspecified food |
| 361P a | Raw milk | SA13 b | Rice | ED-4 a | Raw poultry meat |
| 362P a | Raw milk | SA41 b | Salad | AB-8802 a | Raw poultry meat |
| Isolate | Species | MSSA/MRSA | ST |
|---|---|---|---|
| S1 2021 | S. argenteus | - | - |
| ATCC 29213 | S. aureus | MSSA | - |
| ATCC BAA-1556 (USA300) | S. aureus | MRSA | 8 |
| ATCC BAA-1707 (MW2/USA400) | S. aureus | MRSA | 1 |
| NRS385 (USA500) | S. aureus | MRSA | 8 |
| ATCC 700699 (MU50) | S. aureus | MRSA | 5 |
| NR-45906 (COL) | S. aureus | MRSA | 250 |
| ATCC BAA-44 (HPV107) | S. aureus | MRSA | 247 |
| CAUD | S. aureus | MRSA | 88 |
| ATCC BAA 2312-M10/0148 | S. aureus | MRSA | 130 |
| HU25 (BEC) | S. aureus | MRSA | 239 |
| S2 2017 | S. aureus | MRSA | 111 |
| S6 2017 | S. aureus | MRSA | 398 |
| S1 2018 | S. aureus | MRSA | 30 |
| S4 2018 | S. aureus | MRSA | 93 |
| S5 2018 | S. aureus | MRSA | 2361 |
| S1 2019 | S. aureus | MRSA | 8 |
| S5 2019 | S. aureus | MSSA | 398 |
| S2 2021 | S. aureus | MRSA | 22 |
| S3 2021 | S. aureus | MRSA | 398 |
| S1 2022 | S. aureus | MRSA | 121 |
| S2 2022 | S. aureus | MSSA | 1 |
| S3 2022 | S. aureus | MSSA | 152 |
| ATCC 29971 | S. xylosus | - | - |
| ATCC 27847 | S. hominis | - | - |
| ATCC 14990 | S. epidermidis | - | - |
| ATCC 27848 | S. simulans | - | - |
| ATCC 49052 | S. intermedius | - | - |
| ATCC 29970 | S. haemolyticus | - | - |
| ATCC 27840 | S. capitis | - | - |
| Samples | Run 1 | Run 2 | Inter-Assay | |||
|---|---|---|---|---|---|---|
| Tm ± SD c | CV% | Tm ± SD c | CV% | Tm ± SD c | CV% | |
| Reference Strains | ||||||
| S. argenteus DSM 28299T | 75.94 ± 0.02 | 0.03 | 75.92 ± 0.04 | 0.05 | 75.93 ± 0.03 | 0.04 |
| S. aureus DSM 20231T | 77.24 ± 0.02 | 0.02 | 77.20 ± 0.04 | 0.05 | 77.22 ± 0.04 | 0.05 |
| Isolates | ||||||
| S. aureus (n = 13) a | 77.44 ± 0.05 | 0.06 | 77.25 ± 0.04 | 0.05 | 77.34 ± 0.11 | 0.14 |
| S. aureus (n = 6) b | 77.12 ± 0.03 | 0.05 | 77.20 ± 0.03 | 0.04 | 77.16 ± 0.05 | 0.07 |
| TOT S. aureus (n = 19) a,b | 77.34 ± 0.16 | 0.21 | 77.23 ± 0.04 | 0.05 | 77.28 ± 0.13 | 0.17 |
| Reference Strains | Genome Equivalents | Ct a | SD b |
|---|---|---|---|
| S. argenteus DSM 28299T | 107 | 15.20 | 0.17 |
| 106 | 17.44 | 0.07 | |
| 105 | 20.44 | 0.08 | |
| 104 | 23.61 | 0.13 | |
| 103 | 27.18 | 0.07 | |
| 102 | 30.85 | 0.18 | |
| 101 | 33.97 | 0.17 | |
| S. aureus DSM 20231T | 107 | 15.59 | 0.30 |
| 106 | 17.93 | 0.07 | |
| 105 | 21.18 | 0.04 | |
| 104 | 24.39 | 0.22 | |
| 103 | 27.90 | 0.19 | |
| 102 | 31.44 | 0.17 | |
| 101 | 34.72 | 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chieffi, D.; Bongiorno, D.; Licitra, A.; Campanile, F.; Fusco, V. A Novel Approach Based on Real-Time PCR with High-Resolution Melting Analysis for the Simultaneous Identification of Staphylococcus aureus and Staphylococcus argenteus. Foods 2024, 13, 3004. https://doi.org/10.3390/foods13183004
Chieffi D, Bongiorno D, Licitra A, Campanile F, Fusco V. A Novel Approach Based on Real-Time PCR with High-Resolution Melting Analysis for the Simultaneous Identification of Staphylococcus aureus and Staphylococcus argenteus. Foods. 2024; 13(18):3004. https://doi.org/10.3390/foods13183004
Chicago/Turabian StyleChieffi, Daniele, Dafne Bongiorno, Anna Licitra, Floriana Campanile, and Vincenzina Fusco. 2024. "A Novel Approach Based on Real-Time PCR with High-Resolution Melting Analysis for the Simultaneous Identification of Staphylococcus aureus and Staphylococcus argenteus" Foods 13, no. 18: 3004. https://doi.org/10.3390/foods13183004
APA StyleChieffi, D., Bongiorno, D., Licitra, A., Campanile, F., & Fusco, V. (2024). A Novel Approach Based on Real-Time PCR with High-Resolution Melting Analysis for the Simultaneous Identification of Staphylococcus aureus and Staphylococcus argenteus. Foods, 13(18), 3004. https://doi.org/10.3390/foods13183004