
High Fischer Ratio Oligopeptides of Gluten Alleviate Alcohol-Induced Liver Damage by Regulating Lipid Metabolism and Oxidative Stress in Rats

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of HFO
2.3. Characterization of HFO
2.3.1. Amino Acid Analysis
2.3.2. Microscopic Morphology (SEM)
2.3.3. Determination of Antioxidant Activity
- (1)
- DPPH free radical scavenging
- (2)
- OH radical scavenging rate experiment
- (3)
- Measurement of ABTS free radical scavenging capacity
- (4)
- Measurement of iron reducing power
2.4. Animal Experimental Design
2.5. Liver Index Assay
2.6. Serum Biochemical Analysis
2.7. Antioxidant Analysis of the Liver
2.8. Histological Analysis
2.9. RNA Extraction and mRNA Quantification by qRT-PCR
2.10. Statistical Analysis
3. Results
3.1. Amino Acid Analysis
3.2. Micromorphology of High Fischer Ratio Oligopeptides
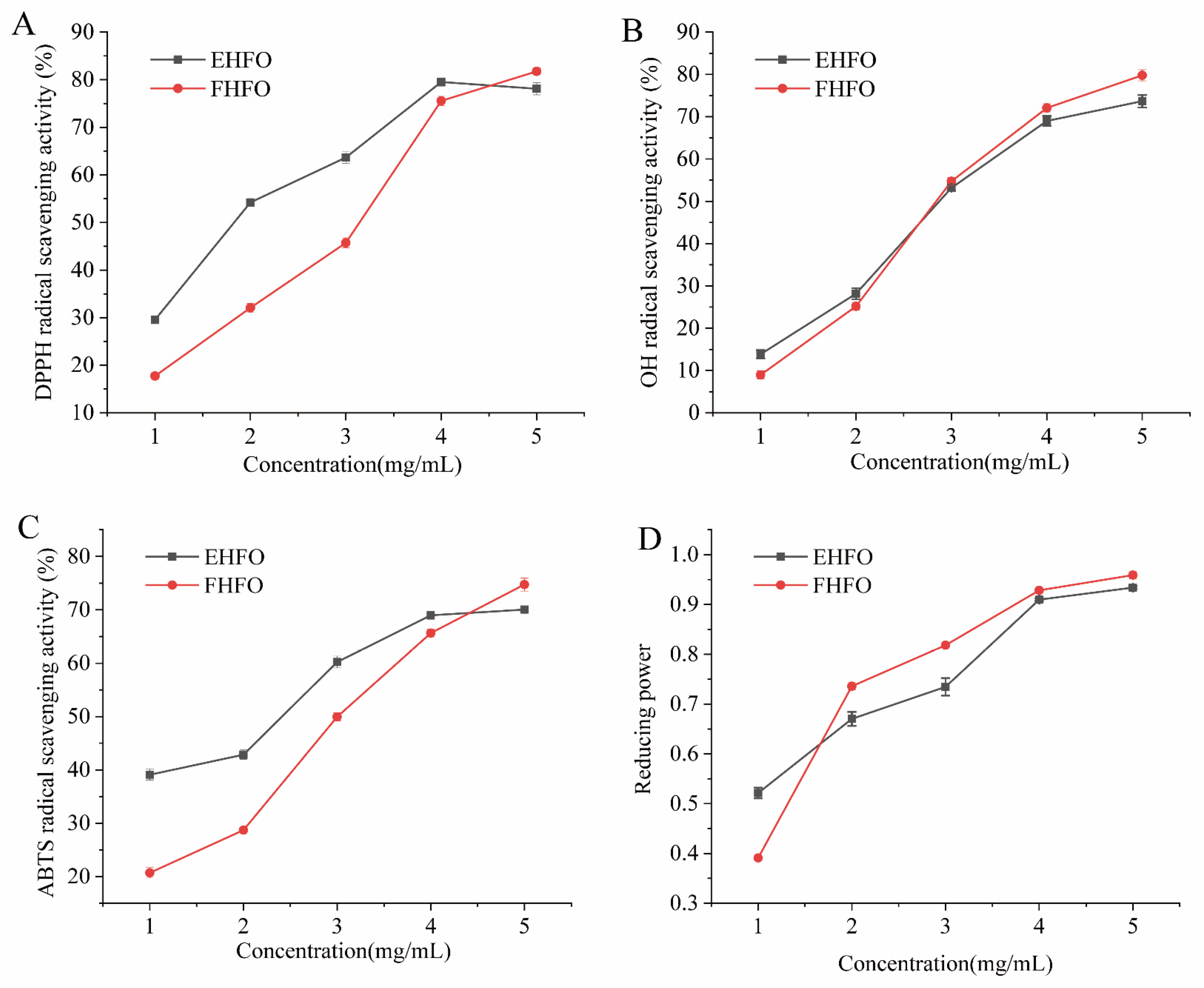
3.3. Antioxidant Analysis of HFO
3.4. Effect of HFO on Liver Index
3.5. Effect of HFO on Serum Biochemical Factors
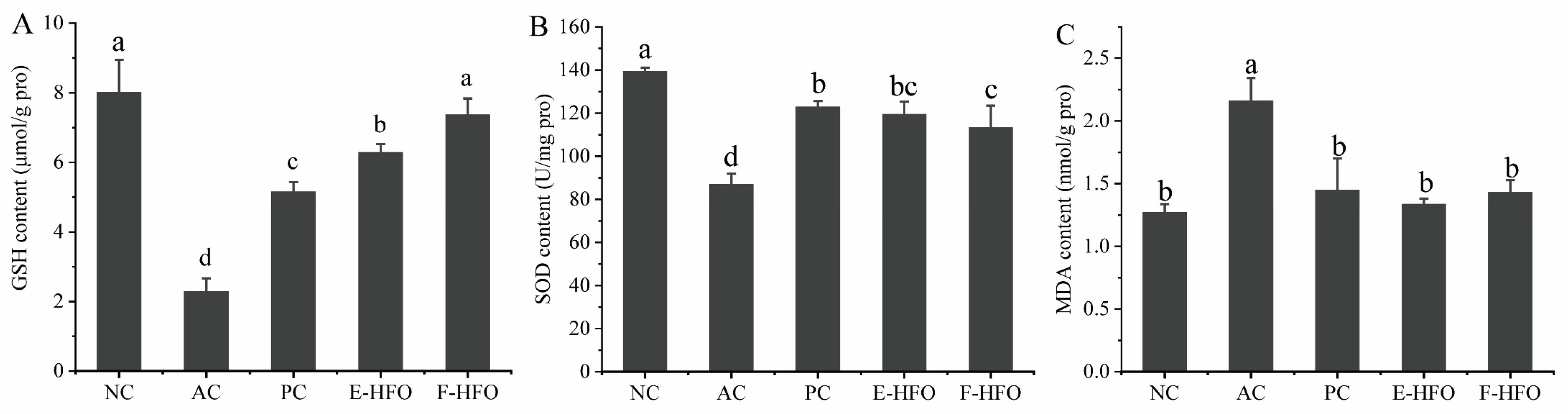
3.6. Effect of HFO on Liver Antioxidant Capacity
3.7. Effect of HFO on Liver Tissue Status
3.8. Effects of HFOs on the Expression of ADH4, ALDH2, and CYP2E1 in Rat Liver
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bergman, R.N.; Piccinini, F.; Kabir, M.; Ader, M. Novel aspects of the role of the liver in carbohydrate metabolism. Metab. Clin. Exp. 2019, 99, 119–125. [Google Scholar] [CrossRef]
- Yoo, J.E.; Han, K.; Shin, D.W.; Kim, D.; Kim, B.S.; Chun, S.; Jeon, K.H.; Jung, W.; Park, J.; Park, J.H.; et al. Association between changes in alcohol consumption and cancer risk. JAMA Netw. Open 2022, 5, e2228544. [Google Scholar] [CrossRef]
- Felgendreff, P.; Raschzok, N.; Kunze, K.; Leder, A.; Lippert, S.; Klunk, S.; Tautenhahn, H.; Hau, H.; Schmuck, R.B.; Reutzel-Selke, A.; et al. Tissue-based mirna mapping in alcoholic liver cirrhosis: Different profiles in cirrhosis with or without hepatocellular carcinoma. Biomarkers 2020, 25, 62–68. [Google Scholar] [CrossRef]
- Goltz, D.; Hittetiya, K.; Vössing, L.M.; Kirfel, J.; Spengler, U.; Fischer, H. A1-antitrypsin pimz heterozygosity has an independent aggravating effect on liver fibrosis in alcoholic liver disease. Virchows Arch. 2014, 465, 539–546. [Google Scholar] [CrossRef]
- Effenberger, M.; Widjaja, A.A.; Grabherr, F.; Schaefer, B.; Grander, C.; Mayr, L.; Schwaerzler, J.; Enrich, B.; Moser, P.; Fink, J.; et al. Interleukin-11 drives human and mouse alcohol-related liver disease. Gut 2023, 72, 168–179. [Google Scholar] [CrossRef]
- Chen, X.; Li, H.; Guo, F.; Hoffmeister, M.; Brenner, H. Alcohol consumption, polygenic risk score, and early- and late-onset colorectal cancer risk. EClinicalMedicine 2022, 49, 101460. [Google Scholar] [CrossRef]
- Park, J.H.; Han, K.; Hong, J.Y.; Park, Y.S.; Park, J.O. Association between alcohol consumption and pancreatic cancer risk differs by glycaemic status: A nationwide cohort study. Eur. J. Cancer 2022, 163, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Rumgay, H.; Shield, K.; Charvat, H.; Ferrari, P.; Sornpaisarn, B.; Obot, I.; Islami, F.; Lemmens, V.E.P.P.; Rehm, J.; Soerjomataram, I. Global burden of cancer in 2020 attributable to alcohol consumption: A population-based study. Lancet Oncol. 2021, 22, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Krittanawong, C.; Isath, A.; Rosenson, R.S.; Khawaja, M.; Wang, Z.; Fogg, S.E.; Virani, S.S.; Qi, L.; Cao, Y.; Long, M.T.; et al. Alcohol consumption and cardiovascular health. Am. J. Med. 2022, 135, 1213–1230. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Moschen, A.R.; Kaneider, N.C. Pathways of liver injury in alcoholic liver disease. J. Hepatol. 2011, 55, 1159–1161. [Google Scholar] [CrossRef]
- Yang, Y.M.; Cho, Y.E.; Hwang, S. Crosstalk between oxidative stress and inflammatory liver injury in the pathogenesis of alcoholic liver disease. Int. J. Mol. Sci. 2022, 23, 774. [Google Scholar] [CrossRef]
- Rodriguez, W.E.; Wahlang, B.; Wang, Y.; Zhang, J.; Vadhanam, M.V.; Joshi Barve, S.; Bauer, P.; Cannon, R.; Ahmadi, A.R.; Sun, Z.; et al. Phosphodiesterase 4 inhibition as a therapeutic target for alcoholic liver disease: From bedside to bench. Hepatology 2019, 70, 1958–1971. [Google Scholar] [CrossRef]
- Kong, X.; Liang, W.; Li, X.; Qiu, M.; Xu, W.; Chen, H. Characterization of an acidic polysaccharides from carrot and its hepatoprotective effect on alcoholic liver injury in mice. Chem. Biodivers. 2021, 18, e2100359. [Google Scholar] [CrossRef]
- Ren, R.; He, Y.; Ding, D.; Cui, A.; Bao, H.; Ma, J.; Hou, X.; Li, Y.; Feng, D.; Li, X.; et al. Aging exaggerates acute-on-chronic alcohol-induced liver injury in mice and humans by inhibiting neutrophilic sirtuin 1-c/ebp α-mirna-223 axis. Hepatology 2022, 75, 646–660. [Google Scholar] [CrossRef]
- Ume Salma, N.; Govindaraju, K.; Gnanesh Kumar, B.S.; Muthukumar, S.P.; Jyothi Lakshmi, A. Ameliorative effect of enhanced fischer ratio flaxseed protein hydrolysate in combination with antioxidant micronutrients on ethanol-induced hepatic damage in a rat model. Brit. J. Nutr. 2022, 127, 696–710. [Google Scholar] [CrossRef] [PubMed]
- Lan, C.; Zhao, Y.; Li, X.; Wang, B. High fischer ratio oligopeptides determination from antartic krill: Preparation, peptides profiles, and in vitro antioxidant activity. J. Food Biochem. 2019, 43, e12827. [Google Scholar] [CrossRef] [PubMed]
- Korenaga, M.; Nishina, S.; Korenaga, K.; Tomiyama, Y.; Yoshioka, N.; Hara, Y.; Sasaki, Y.; Shimonaka, Y.; Hino, K. Branched-chain amino acids reduce hepatic iron accumulation and oxidative stress in hepatitis c virus polyprotein-expressing mice. Liver Int. 2015, 35, 1303–1314. [Google Scholar] [CrossRef] [PubMed]
- Tamai, Y.; Chen, Z.; Wu, Y.; Okabe, J.; Kobayashi, Y.; Chiba, H.; Hui, S.; Eguchi, A.; Iwasa, M.; Ito, M.; et al. Branched-chain amino acids and l-carnitine attenuate lipotoxic hepatocellular damage in rat cirrhotic liver. Biomed. Pharmacother. 2021, 135, 111181. [Google Scholar] [CrossRef] [PubMed]
- Di Miceli, M.; Gronier, B. Pharmacology, systematic review and recent clinical trials of metadoxine. Rev. Recent Clin. Trials 2018, 13, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Shi, T.; Li, Y.; Liu, X. Optimization of enzymatic hydrolysis and activated carbon adsorption for producing high fischer ratio peptides from soy protein. Int. J. Pept. Res. Ther. 2021, 27, 1363–1372. [Google Scholar] [CrossRef]
- Wang, Y.; Song, X.; Feng, Y.; Cui, Q. Changes in peptidomes and fischer ratios of corn-derived oligopeptides depending on enzyme hydrolysis approaches. Food Chem. 2019, 297, 124931. [Google Scholar] [CrossRef]
- Chen, G.; Chen, B.; Song, D. Co-microbiological regulation of phenolic release through solid-state fermentation of corn kernels (Zea mays L.) To improve their antioxidant activity. LWT 2021, 142, 111003. [Google Scholar] [CrossRef]
- Mei, X.; Yang, W.; Huang, G.; Huang, H. The antioxidant activities of balsam pear polysaccharide. Int. J. Biol. Macromol. 2020, 142, 232–236. [Google Scholar] [CrossRef]
- He, S.; Ma, Y.; Zhou, S.; Song, W.; Wang, R. Antioxidant activities of fermented soybean prepared with bacillus subtilis. Asian J. Chem. 2013, 25, 10565–10568. [Google Scholar] [CrossRef]
- Ma, H.; Liu, Y.; Li, Z.; Yu, L.; Gao, Y.; Ye, X.; Yang, B.; Li, H.; Shi, J. Propofol protects against hepatic ischemia reperfusion injury via inhibiting bnip3-mediated oxidative stress. Inflammation 2021, 44, 1288–1301. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Liao, A.; Cui, Y.; Yu, G.; Hou, Y.; Pan, L.; Chen, W.; Zheng, S.; Li, X.; Ma, J.; et al. Wheat embryo globulin protects against acute alcohol-induced liver injury in mice. Food Chem. Toxicol. 2021, 153, 112240. [Google Scholar] [CrossRef] [PubMed]
- Fotio, A.L.; Nguepi, M.S.D.; Tonfack, L.B.; Temdie, R.J.G.; Nguelefack, T.B. Acetaminophen induces liver injury and depletes glutathione in mice brain: Prevention by moringa oleifera extract. S. Afr. J. Bot. 2020, 129, 317–323. [Google Scholar] [CrossRef]
- Li, T.; Tian, Y.; Sun, F.; Wang, Z.; Zhou, N. Preparation of high fischer’s ratio corn oligopeptides using directed enzymatic hydrolysis combined with adsorption of aromatic amino acids for efficient liver injury repair. Process Biochem. 2019, 84, 60–72. [Google Scholar] [CrossRef]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef]
- Helmut, K.; Seitz, R.B.H.C. Alcoholic liver disease. Nat. Rev. Dis. Primers 2018, 4, 17. [Google Scholar] [CrossRef]
- Qin, Y.; Cheng, M.; Fan, X.; Shao, X.; Wang, C.; Jiang, H.; Zhang, X. Preparation and antioxidant activities of high fischer’s ratio oligopeptides from goat whey. Food Sci. Anim. Resour. 2022, 42, 800–815. [Google Scholar] [CrossRef] [PubMed]
- Chinnappan, R.; Mir, T.A.; Alsalameh, S.; Makhzoum, T.; Adeeb, S.; Al-Kattan, K.; Yaqinuddin, A. Aptasensors are conjectured as promising alt and ast diagnostic tools for the early diagnosis of acute liver injury. Life 2023, 13, 1273. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Qing, S.; Miao, Z.; Luo, H.; Tian, J.; Zhang, X.; Wang, S.; Zhang, T.; Miao, C. Hepatic nampt deficiency aggravates dyslipidemia and fatty liver in high fat diet fed mice. Cells 2023, 12, 568. [Google Scholar] [CrossRef] [PubMed]
- Yanti; Violina, V.; Putri, C.E.; Lay, B.W. Branched chain amino acid content and antioxidant activity of mung bean tempeh powder for developing oral nutrition supplements. Foods 2023, 12, 2789. [Google Scholar] [CrossRef] [PubMed]
- Higuera-De, L.T.F.; Servin-Caamano, A.I.; Cruz-Herrera, J.; Serralde-Zuniga, A.E.; Abdo-Francis, J.M.; Gutierrez-Reyes, G.; Perez-Hernandez, J.L. Treatment with metadoxine and its impact on early mortality in patients with severe alcoholic hepatitis. Ann. Hepatol. 2014, 13, 343–352. [Google Scholar] [CrossRef]
- Barry Heffernan, C.; Ekena, J.; Dowling, S.; Pinkerton, M.E.; Viviano, K. Biomarkers of oxidative stress as an assessment of the redox status of the liver in dogs. J. Vet. Intern. Med. 2019, 33, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Busch, C.J.L.; Hendrikx, T.; Weismann, D.; Jäckel, S.; Walenbergh, S.M.A.; Rendeiro, A.F.; Weißer, J.; Puhm, F.; Hladik, A.; Göderle, L.; et al. Malondialdehyde epitopes are sterile mediators of hepatic inflammation in hypercholesterolemic mice. Hepatology 2017, 65, 1181–1195. [Google Scholar] [CrossRef]
- Xu, Y.; Jing, X.; Zhai, W.; Li, X. The enantioselective enrichment, metabolism, and toxicity of fenoxaprop-ethyl and its metabolites in zebrafish. Chirality 2020, 32, 990–997. [Google Scholar] [CrossRef]
- Lu, J.; Lyu, Y.; Li, M.; Sun, J.; Huang, Z.; Lu, F.; Lu, Z. Alleviating acute alcoholic liver injury in mice with bacillus subtilis co-expressing alcohol dehydrogenase and acetaldehyde dehydrogenase. J. Funct. Foods 2018, 49, 342–350. [Google Scholar] [CrossRef]
- Sun, Q.; Wang, G.; Gao, L.; Shi, L.; Qi, Y.; Lv, X.; Jin, Y. Roles of cyp2e1 in 1,2-dichloroethane-induced liver damage in mice. Environ. Toxicol. 2016, 31, 1430–1438. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | F Primer | R Primer |
|---|---|---|
| ADH4 | GGGAACGTTCTTTGGTGGTTG | ATCAGGTCAATCGCGTCGTT |
| ALDH2 | ATCCTCGGCTACATCAAATCG | GTCTTTTACGTCCCCGAACAC |
| CYP2E1 | CAAGTCTTTCACCAAGTTGGCA | CCCCCGTCCAGAAAACTCAT |
| GAPDH | GGTGAAGGTCGGTGTGAACG | CTCGCTCCTGGAAGATGGTG |
| Free Amino Acids | EHFO | FHFO | Type |
|---|---|---|---|
| Asp | 1.168 | 0.657 | |
| Thr | 0.939 | 0.634 | |
| Ser | 1.696 | 0.782 | |
| Glu | 10.611 | 7.435 | |
| Gly | 1.176 | 1.000 | |
| Ala | 1.032 | 1.526 | |
| Cys | 0.175 | 0.126 | |
| Val | 2.380 | 2.320 | BCAAs |
| Met | 0.465 | 0.548 | |
| Ile | 1.868 | 1.801 | BCAAs |
| Leu | 2.224 | 2.722 | BCAAs |
| Tyr | 0.128 | 0.127 | AAAs |
| Phe | 0.149 | 0.145 | AAAs |
| His | 0.796 | 0.883 | |
| Lys | 0.576 | 0.601 | |
| Arg | 0.981 | 0.432 | |
| Pro | 2.933 | 2.946 |
| Free Amino Acids | EHFO | FHFO |
|---|---|---|
| BCAAs | 6.472 | 6.843 |
| AAAs | 0.276 | 0.272 |
| Fischer ratio | 23.436 | 25.195 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, P.; Hou, Y.; Chen, X.; Zhang, M.; Hu, Z.; Chen, L.; Huang, J. High Fischer Ratio Oligopeptides of Gluten Alleviate Alcohol-Induced Liver Damage by Regulating Lipid Metabolism and Oxidative Stress in Rats. Foods 2024, 13, 436. https://doi.org/10.3390/foods13030436
Zhao P, Hou Y, Chen X, Zhang M, Hu Z, Chen L, Huang J. High Fischer Ratio Oligopeptides of Gluten Alleviate Alcohol-Induced Liver Damage by Regulating Lipid Metabolism and Oxidative Stress in Rats. Foods. 2024; 13(3):436. https://doi.org/10.3390/foods13030436
Chicago/Turabian StyleZhao, Penghui, Yinchen Hou, Xinyang Chen, Mingyi Zhang, Zheyuan Hu, Lishui Chen, and Jihong Huang. 2024. "High Fischer Ratio Oligopeptides of Gluten Alleviate Alcohol-Induced Liver Damage by Regulating Lipid Metabolism and Oxidative Stress in Rats" Foods 13, no. 3: 436. https://doi.org/10.3390/foods13030436
APA StyleZhao, P., Hou, Y., Chen, X., Zhang, M., Hu, Z., Chen, L., & Huang, J. (2024). High Fischer Ratio Oligopeptides of Gluten Alleviate Alcohol-Induced Liver Damage by Regulating Lipid Metabolism and Oxidative Stress in Rats. Foods, 13(3), 436. https://doi.org/10.3390/foods13030436