
Dietary Tomato Pectin Attenuates Hepatic Insulin Resistance and Inflammation in High-Fat-Diet Mice by Regulating the PI3K/AKT Pathway
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation and Extraction of TP
2.2. Animals and Treatment
2.3. Biochemical Analysis
2.4. Glucose (GTT) and Insulin Tolerance (ITT) Tests
2.5. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.6. Western Blot Analysis
2.7. Statistical Analyses
3. Results
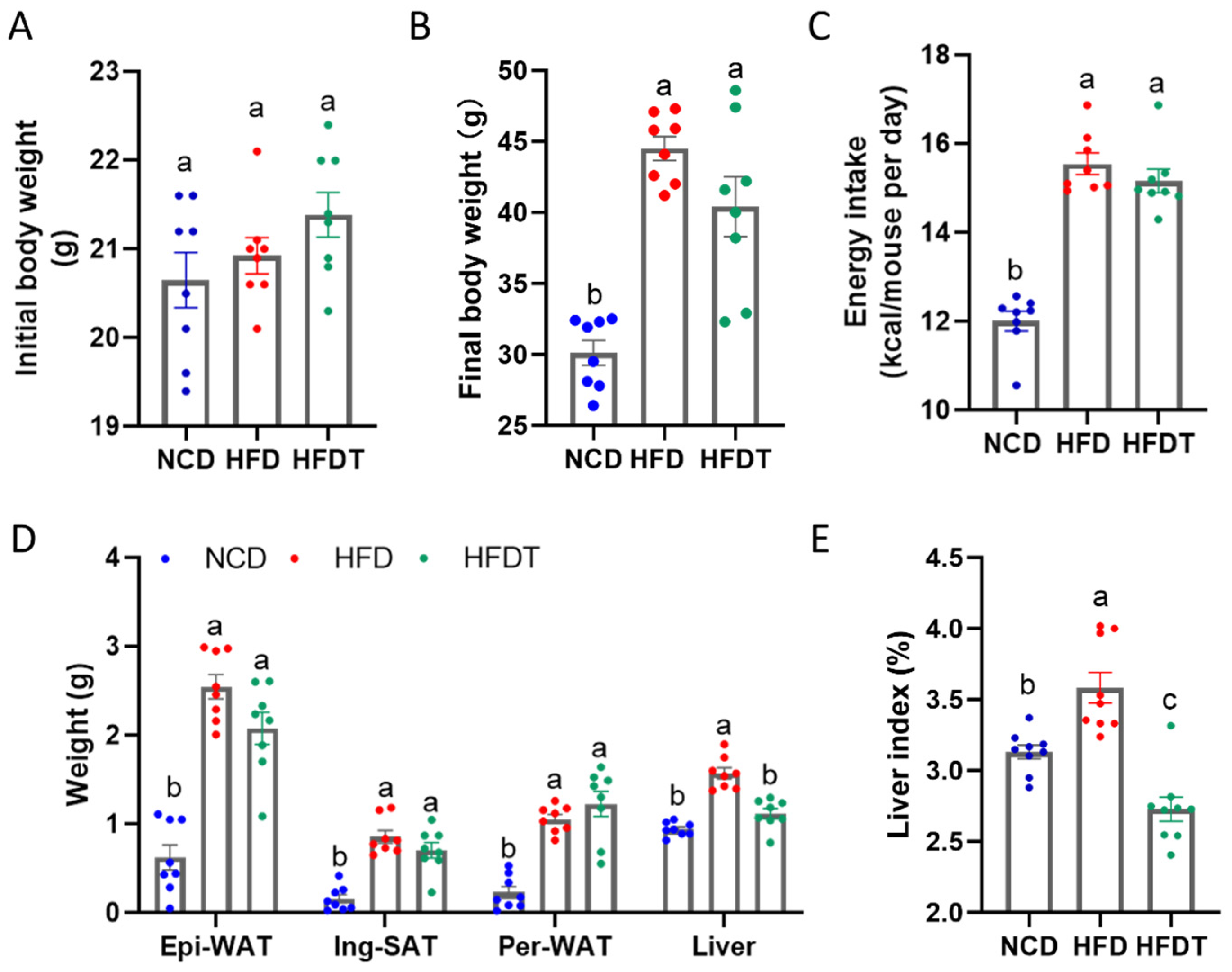
3.1. Effect of TP on Body Weight and Liver Index in HFD-Fed Mice
3.2. TP Ameliorated Fat Accumulation and Hepatic Injury in HFD-Fed Mice
3.3. TP Improved Glucose Tolerance and Insulin Sensitivity in HFD-Fed Mice
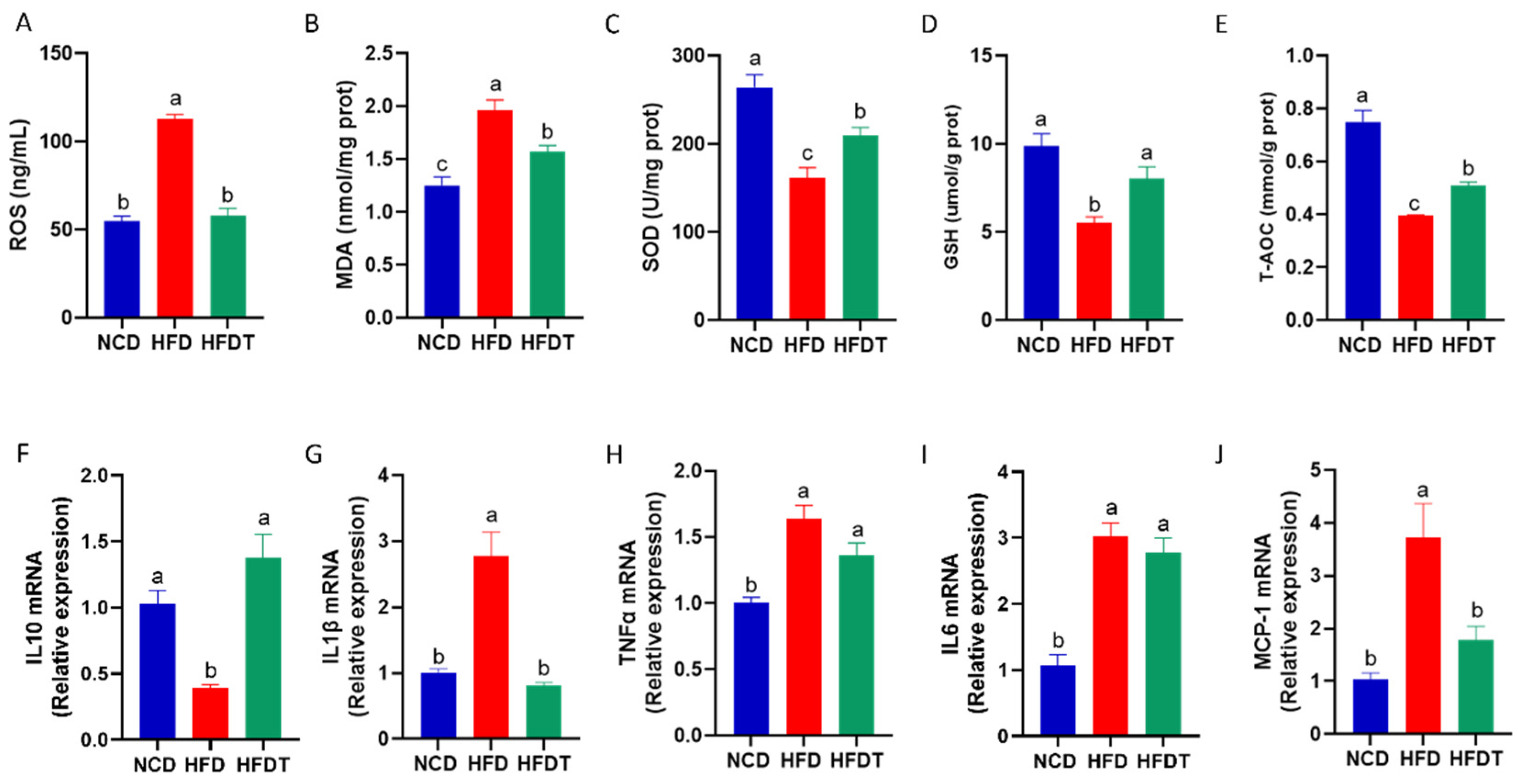
3.4. TP Attenuated Hepatic Oxidative Stress and Inflammation in HFD-Fed Mice
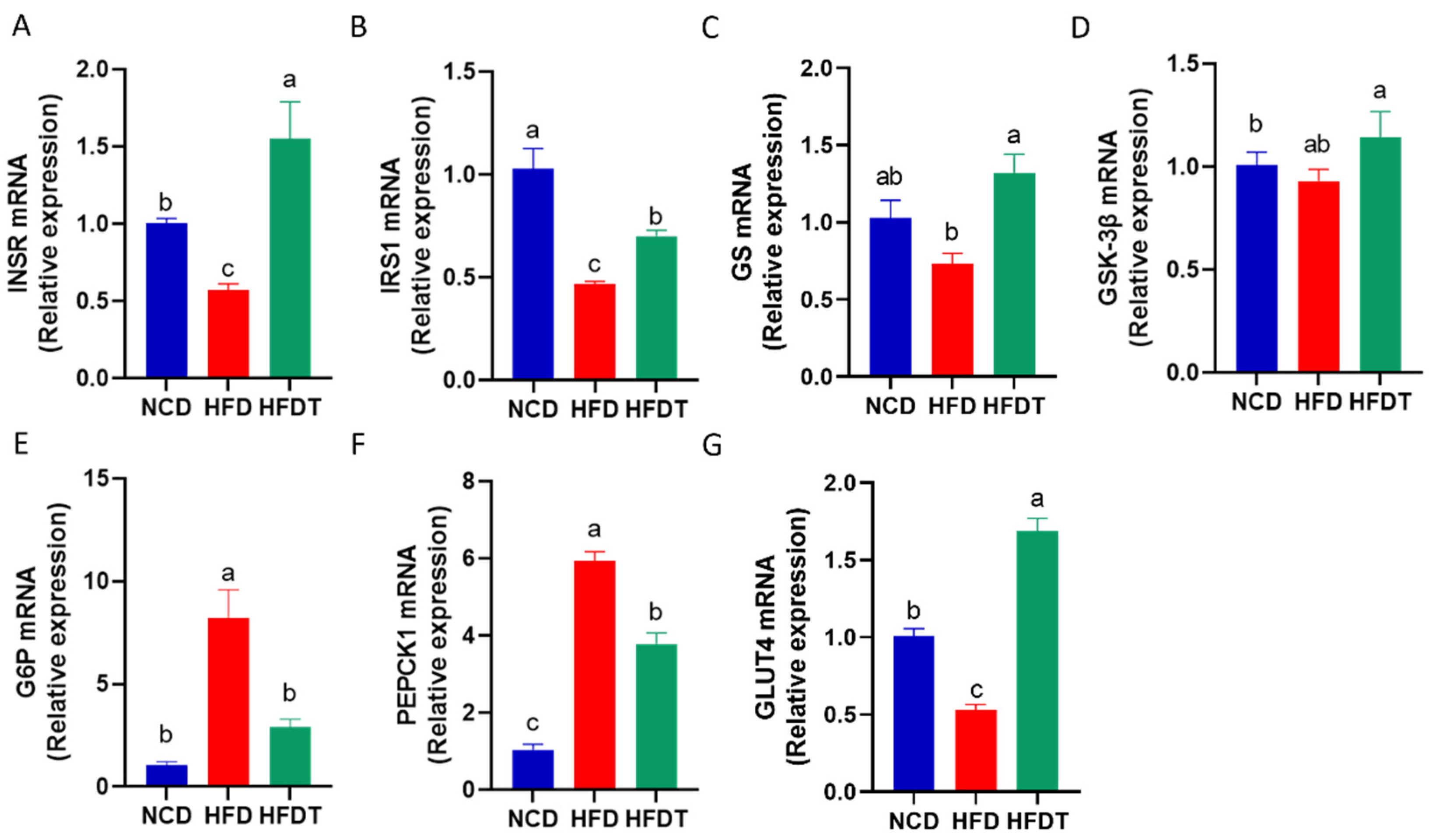
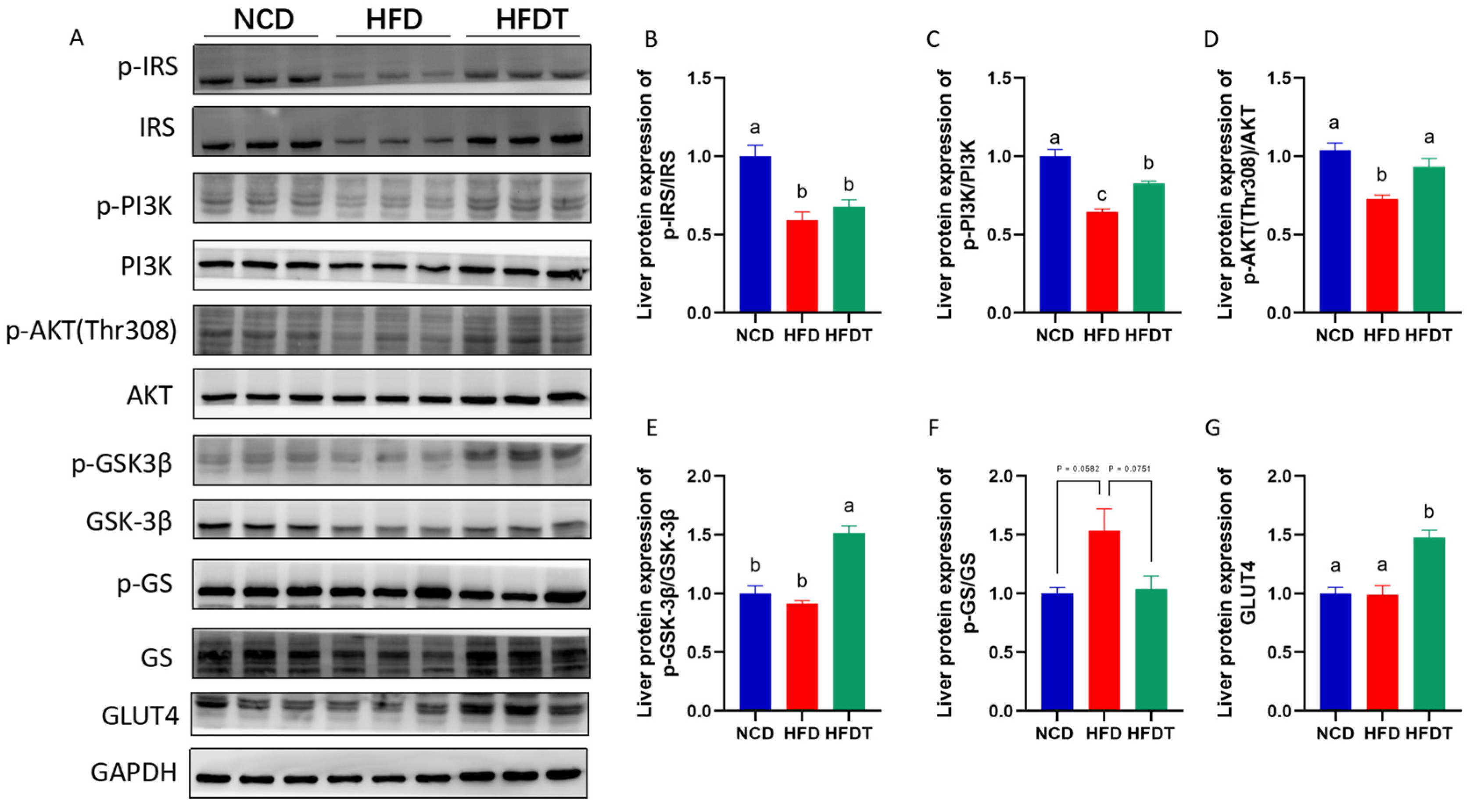
3.5. TP Enhanced Hepatic Insulin Signaling in HFD-Fed Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Costello, K.R.; Schones, D.E. Chromatin Modifications in Metabolic Disease: Potential Mediators of Long-Term Disease Risk. Wiley Interdiscip. Rev. Syst. Biol. Med. 2018, 10, e1416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yuan, H.; Zhang, C.; Guan, Y.; Wu, Y.; Ling, F.; Niu, Y.; Li, Y. Epigallocatechin Gallate Improves Insulin Resistance in Hepg2 Cells through Alleviating Inflammation and Lipotoxicity. Diabetes Res. Clin. Pract. 2018, 142, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.C.; Vatner, D.F.; Shulman, G.I. Regulation of Hepatic Glucose Metabolism in Health and Disease. Nat. Rev. Endocrinol. 2017, 13, 572–587. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Memon, R.S.; Shahid, I.; Rizwan, T.; Zaman, M.; Menezes, R.G.; Kumar, S.; Siddiqi, T.J.; Usman, M.S. Antidiabetic Drugs and Non-Alcoholic Fatty Liver Disease: A Systematic Review, Meta-Analysis and Evidence Map. Dig. Liver Dis. 2021, 53, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, S.Y.; Choi, C.S. Insulin Resistance: From Mechanisms to Therapeutic Strategies. Diabetes Metab. J. 2022, 46, 15–37. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K. Mcp-1 Contributes to Macrophage Infiltration into Adipose Tissue, Insulin Resistance, and Hepatic Steatosis in Obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and Metabolic Disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Wang, P.; Wang, J.; Li, D.; Ke, W.; Chen, F.; Hu, X. Targeting the Gut Microbiota with Resveratrol: A Demonstration of Novel Evidence for the Management of Hepatic Steatosis. J. Nutr. Biochem. 2020, 81, 108363. [Google Scholar] [CrossRef]
- Ahmed, M.A.; Mohamed, M.A.; Rashed, L.A.; Elbast, S.A.; Ahmed, E.A. Rice Bran Oil Improves Insulin Resistance by Affecting the Expression of Antioxidants and Lipid-Regulatory Genes. Lipids 2018, 53, 505–515. [Google Scholar] [CrossRef]
- Gao, W.; Xiao, C.; Hu, J.; Chen, B.; Wang, C.; Cui, B.; Deng, P.; Yang, J.; Deng, Z. Qing Brick Tea (Qbt) Aqueous Extract Protects Monosodium Glutamate-Induced Obese Mice against Metabolic Syndrome and Involves up-Regulation Transcription Factor Nuclear Factor-Erythroid 2-Related Factor 2 (Nrf2) Antioxidant Pathway. Biomed. Pharmacother. 2018, 103, 637–644. [Google Scholar] [CrossRef]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose Tissue and Insulin Resistance in Obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef]
- Jiang, T.; Gao, X.; Wu, C.; Tian, F.; Lei, Q.; Bi, J.; Xie, B.; Wang, H.; Chen, S.; Wang, X. Apple-Derived Pectin Modulates Gut Microbiota, Improves Gut Barrier Function, and Attenuates Metabolic Endotoxemia in Rats with Diet-Induced Obesity. Nutrients 2016, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Bi, J.; Yi, J.; Peng, J.; Ma, Q. Dose-Dependent Effects of Apple Pectin on Alleviating High Fat-Induced Obesity Modulated by Gut Microbiota and Scfas. Food Sci. Hum. Wellness 2022, 11, 143–154. [Google Scholar] [CrossRef]
- Bagabaldo, P.A.; Atienza, L.M.; Castillo-Israel, K.A.T.; Estacio, M.A.C.; Gaban, P.J.V.; Maniwang, J.R.C.; Gapasin, R.P.; Estribillo, A.G.M.; Cena-Navarro, R.B. ‘Saba’banana (Musa Acuminata X Balbisiana Bbb Group) Peel Pectin Supplementation Improves Biomarkers of Obesity and Associated Blood Lipid Disorders in Obese Hypercholesterolemic Mice. Curr. Res. Food Sci. 2022, 5, 251–260. [Google Scholar] [CrossRef]
- Ren, T.; Xu, M.; Zhou, S.; Ren, J.; Li, B.; Jiang, P.; Li, H.; Wu, W.; Chen, C.; Fan, M. Structural Characteristics of Mixed Pectin from Ginseng Berry and Its Anti-Obesity Effects by Regulating the Intestinal Flora. Int. J. Biol. Macromol. 2023, 242, 124687. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Z.; Hu, D.; Xiao, K.; Wu, J. Efficient Extraction of Pectin from Sisal Waste by Combined Enzymatic and Ultrasonic Process. Food Hydrocoll. 2018, 79, 189–196. [Google Scholar] [CrossRef]
- Wang, P.; Li, D.; Ke, W.; Liang, D.; Hu, X.; Chen, F. Resveratrol-Induced Gut Microbiota Reduces Obesity in High-Fat Diet-Fed Mice. Int. J. Obes. 2020, 44, 213–225. [Google Scholar] [CrossRef]
- Pedersen, S.S.; Prause, M.; Sørensen, C.; Størling, J.; Moritz, T.; Mariño, E.; Billestrup, N. Targeted Delivery of Butyrate Improves Glucose Homeostasis, Reduces Hepatic Lipid Accumulation and Inflammation in db/db Mice. Int. J. Mol. Sci. 2023, 24, 4533. [Google Scholar] [CrossRef]
- Liu, Y.; Dong, M.; Yang, Z.; Pan, S. Anti-Diabetic Effect of Citrus Pectin in Diabetic Rats and Potential Mechanism Via Pi3k/Akt Signaling Pathway. Int. J. Biol. Macromol. 2016, 89, 484–488. [Google Scholar] [CrossRef]
- Wu, T.R.; Lin, C.S.; Chang, C.J.; Lin, T.L.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Lu, C.C.; Young, J.D.; Lai, H.C. Gut Commensal Parabacteroides Goldsteinii Plays a Predominant Role in the Anti-Obesity Effects of Polysaccharides Isolated from Hirsutella Sinensis. Gut 2019, 68, 248–262. [Google Scholar] [CrossRef]
- Loh, K.; Deng, H.; Fukushima, A.; Cai, X.; Boivin, B.; Galic, S.; Bruce, C.; Shields, B.J.; Skiba, B.; Ooms, L.M. Reactive Oxygen Species Enhance Insulin Sensitivity. Cell Metab. 2009, 10, 260–272. [Google Scholar] [CrossRef]
- Muñoz, A.; Costa, M. Nutritionally Mediated Oxidative Stress and Inflammation. Oxidative Med. Cell. Longev. 2013, 2013, 610950. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Gao, J.; Ke, W.; Wang, J.; Li, D.; Liu, R.; Jia, Y.; Wang, X.; Chen, X.; Chen, F. Resveratrol Reduces Obesity in High-Fat Diet-Fed Mice Via Modulating the Composition and Metabolic Function of the Gut Microbiota. Free. Radic. Biol. Med. 2020, 156, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Qing, Z.; Wu, X.; Wu, X.; Zou, C. Vitamin C Deficiency Aggravates Tumor Necrosis Factor A-Induced Insulin Resistance. Eur. J. Pharmacol. 2018, 829, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Schulze, R.J.; Schott, M.B.; Casey, C.A.; Tuma, P.L.; McNiven, M.A. The Cell Biology of the Hepatocyte: A Membrane Trafficking Machine. J. Cell Biol. 2019, 218, 2096–2112. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, L.; De, S.J.C.; Harari, A.A.; Backs, J.; Olson, E.N.; Tabas, I. Activation of Calcium/Calmodulin-Dependent Protein Kinase Ii in Obesity Mediates Suppression of Hepatic Insulin Signaling. Cell Metab. 2013, 18, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, C.M.; Emanuelli, B.; Kahn, C.R. Critical Nodes in Signalling Pathways: Insights into Insulin Action. Nat. Rev. Mol. Cell Biol. 2006, 7, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zhao, S.X.; Xu, B.Q.; Zhang, Y.Q. Gynura Divaricata Ameliorates Hepatic Insulin Resistance by Modulating Insulin Signalling, Maintaining Glycolipid Homeostasis and Reducing Inflammation in Type 2 Diabetic Mice. Toxicol. Res. 2019, 8, 928–938. [Google Scholar] [CrossRef]
- Luo, K.; Wang, X.; Zhang, G. Starch and Β-Glucan in a Whole-Grain-Like Structural Form Improve Hepatic Insulin Sensitivity in Diet-Induced Obese Mice. Food Funct. 2019, 10, 5091–5101. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum Parameters | NCD | HFD | HFDT |
|---|---|---|---|
| TG (mg/dL) | 0.36 ± 0.02 c | 1.29 ± 0.09 a | 0.54 ± 0.07 b |
| TC (mg/dL) | 3.50 ± 0.19 c | 6.06 ± 0.29 a | 5.12 ± 0.19 b |
| FFAs (μmol/L) | 844.2 ± 11.68 c | 1655 ± 24.72 a | 1534 ± 39.75 b |
| HDL-C (mmol/L) | 1.77 ± 0.51 b | 4.19 ± 0.24 a | 3.71 ± 0.13 a |
| LDL-C (mmol/L) | 0.32 ± 0.08 b | 1.10 ± 0.23 a | 0.70 ± 0.09 ab |
| ALT (U/L) | 20.10 ± 4.51 b | 129.90 ± 53.12 a | 32.48 ± 4.73 b |
| AST (U/L) | 142.05 ± 16.26 b | 204.53 ± 26.29 a | 158.93 ± 33.35 b |
| AdipoQ (ng/mL) | 350.6 ± 6.65 a | 299.6 ± 1.82 b | 333.8 ± 5.46 a |
| Leptin (ng/mL) | 1.38 ± 0.02 b | 1.71 ± 0.04 a | 1.67 ± 0.02 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Wu, K.; Wang, P.; Wang, Y.; Wang, D.; Zhao, W.; Zhao, Y.; Zhang, C.; Zhao, X. Dietary Tomato Pectin Attenuates Hepatic Insulin Resistance and Inflammation in High-Fat-Diet Mice by Regulating the PI3K/AKT Pathway. Foods 2024, 13, 444. https://doi.org/10.3390/foods13030444
Sun J, Wu K, Wang P, Wang Y, Wang D, Zhao W, Zhao Y, Zhang C, Zhao X. Dietary Tomato Pectin Attenuates Hepatic Insulin Resistance and Inflammation in High-Fat-Diet Mice by Regulating the PI3K/AKT Pathway. Foods. 2024; 13(3):444. https://doi.org/10.3390/foods13030444
Chicago/Turabian StyleSun, Jing, Kongyan Wu, Pan Wang, Yubin Wang, Dan Wang, Wenting Zhao, Yuanyuan Zhao, Chunhong Zhang, and Xiaoyan Zhao. 2024. "Dietary Tomato Pectin Attenuates Hepatic Insulin Resistance and Inflammation in High-Fat-Diet Mice by Regulating the PI3K/AKT Pathway" Foods 13, no. 3: 444. https://doi.org/10.3390/foods13030444