
Valorization of Coffee Cherry By-Products Through Fermentation by Human Intestinal Lactobacilli in Functional Fermented Milk Beverages
 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Coffee Cherry Extract Preparation and Preliminary Characterization
2.2. Strains Used and Growing Conditions
2.3. Effect of Coffee Cherry Extracts on Lactobacilli Metabolism
2.4. Preparation of the Coffee Cherry Pulp Extract (CPE)
2.5. CPE Characterization
2.6. CPE Effect on Lactobacilli Growth and Metabolism
2.7. CPE Effect on Lactobacilli Biofilm Formation
2.8. CPE Effect on Lactobacilli Antioxidant Capacities
2.9. CPE Effect on Lactobacilli Antimicrobial Activities
2.10. Manufacture of CPE Fermented Milk Products
2.11. CPE Fermented Milk Product Characterization
2.12. Statistical Analysis
3. Results and Discussion
3.1. Coffee Cherry Extracts: Preliminary Characterization and Selection
3.2. CPE Characterization
3.3. CPE Effect on Bacterial Growth and Metabolism
3.4. Functional and Probiotic Capacities of Selected Strains in the Presence of CPE
3.4.1. CPE Effect on Lactobacilli Biofilm Formation
3.4.2. CPE Effect on Strain Antioxidant Capacity
3.4.3. CPE Effect on Intestinal Lactobacilli Antimicrobial Activity Against Pathogenic Strains
3.5. Manufacture and Characterization of CPE Fermented Milk Products
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). Available online: https://www.fao.org/markets-and-trade/commodities/coffee/en/ (accessed on 27 September 2024).
- Food and Agriculture Organization of the United Nations (FAO). Faostat, Crops and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 27 September 2024).
- Alonso-Salces, R.M.; Serra, F.; Reniero, F.; Héberger, K. Botanical and geographical characterization of green coffee (Coffea arabica and Coffea canephora): Chemometric evaluation of phenolic and methylxanthine contents. J. Agric. Food Chem. 2009, 57, 4224–4235. [Google Scholar] [CrossRef] [PubMed]
- Viencz, T.; Acre, L.B.; Rocha, R.B.; Alves, E.A.; Ramalho, A.R.; de Toledo Benassi, M. Caffeine, trigonelline, chlorogenic acids, melanoidins, and diterpenes contents of Coffea canephora coffees produced in the Amazon. J. Food Compos. Anal. 2023, 117, 105140. [Google Scholar] [CrossRef]
- Freitas, V.V.; Borges, L.L.R.; Vidigal, M.C.T.R.; dos Santos, M.H.; Stringheta, P.C. Coffee: A comprehensive overview of origin, market, and the quality process. Trends Food Sci. Technol. 2024, 146, 104411. [Google Scholar] [CrossRef]
- Torres-Valenzuela, L.S.; Ballesteros-Gómez, A.; Rubio, S. Supramolecular solvent extraction of bioactives from coffee cherry pulp. J. Food Eng. 2020, 278, 109933. [Google Scholar] [CrossRef]
- Pandey, A.; Soccol, C.R.; Nigam, P.; Brand, D.; Mohan, R.; Roussos, S. Biotechnological potential of coffee pulp and coffee husk for bioprocesses. Biochem. Eng. J. 2024, 6, 153–162. [Google Scholar] [CrossRef]
- Gemechu, F.G. Embracing nutritional qualities, biological activities and technological properties of coffee byproducts in functional food formulation. Trends Food Sci. Technol. 2020, 104, 235–261. [Google Scholar] [CrossRef]
- Aloo, O.S.; Gemechu, F.G.; Oh, H.-J.; Kilel, E.C.; Chelliah, R.; Gonfa, G.; Oh, D.-H. Harnessing fermentation for sustainable beverage production: A tool for improving the nutritional quality of coffee bean and valorizing coffee byproducts. Biocatal. Agric. Biotechnol. 2024, 59, 103263. [Google Scholar] [CrossRef]
- Machado, M.; Ferreira, H.; Oliveira, M.B.P.P.; Alves, R.C. Coffee by-products: An underexplored source of prebiotic ingredients. Crit. Rev. Food Sci. Nutr. 2024, 64, 7181–7200. [Google Scholar] [CrossRef]
- Aravind, S.M.; Wichienchot, S.; Tsao, R.; Ramakrishnan, S.; Chakkaravarthi, S. Role of dietary polyphenols on gut microbiota, their metabolites and health benefits. Food Res. Int. 2021, 142, 110189. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenteriol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Dueñas, M.; Muñoz-González, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. A survey of modulation of gut microbiota by dietary polyphenols. BioMed Res. Int. 2015, 2015, 850902. [Google Scholar] [CrossRef]
- Evangelista, S.R.; da Cruz Pedrozo Miguel, M.G.; de Souza Cordeiro, C.; Silva, C.F.; Pinheiro, A.C.M.; Schwan, R.F. Inoculation of starter cultures in a semi-dry coffee (Coffea arabica) fermentation process. Food Microbiol. 2014, 44, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Oberg, T.S.; McMahon, D.J.; Culumber, M.D.; McAuliffe, O.; Oberg, C.J. Invited review: Review of taxonomic changes in dairy-related lactobacilli. J. Dairy Sci. 2024, 105, 2750–2770. [Google Scholar] [CrossRef] [PubMed]
- Gaya, P.; Arqués, J.L.; Medina, M.; Álvarez, I.; Landete, J.M. A New HPLC-PAD/HPLC-ESI-MS Method for the analysis of phytoestrogens produced by bacterial metabolism. Food Anal. Methods 2016, 9, 537–547. [Google Scholar] [CrossRef]
- Gaya, P.; Peirotén, A.; Landete, J.M. Transformation of plant isoflavones into bioactive isoflavones by lactic acid bacteria and bifidobacteria. J. Funct. Foods 2017, 39, 198–205. [Google Scholar] [CrossRef]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Rodríguez, E.; Arqués, J.L.; Rodríguez, R.; Peirotén, A.; Landete, J.M.; Medina, M. Antimicrobial properties of probiotic strains isolated from breast-fed infants. J. Funct. Foods 2012, 4, 542–551. [Google Scholar] [CrossRef]
- Cogan, T.M.; Barbosa, M.; Beuvier, E.; Bianchi-Salvadori, B.; Cocconcelli, P.S.; Fernandes, I.; Gomez, J.; Gomez, R.; Kalantzopoulos, G.; Ledda, A.; et al. Characterization of the lactic acid bacteria in artisanal dairy products. J. Dairy Res. 1997, 64, 409–421. [Google Scholar] [CrossRef]
- Capurso, L. Thirty years of Lactobacillus rhamnosus GG: A Review. J. Clin. Gastroenterol. 2019, 53, S1–S41. [Google Scholar] [CrossRef]
- Rodríguez-Mínguez, E.; Ríos, M.G.; Sánchez, C.; Picon, A. Mangosteen extracts: Effects on intestinal bacteria, and application to functional fermented milk products. Food Res. Int. 2024, 191, 114720. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Lee, B.H.; Cole, S.; Badel-Berchoux, S.; Guillier, L.; Felix, B.; Krezdorn, N.; Hébraud, M.; Bernardi, T.; Sultan, I.; Piveteau, P. Biofilm formation of Listeria monocytogenes strains under food processing environments and pan-genome-wide association study. Front. Microbiol. 2019, 10, 2698. [Google Scholar] [CrossRef]
- Gil-Rodríguez, A.M.; Carrascosa, A.V.; Requena, T. Yeasts in foods and beverages: In vitro characterisation of probiotic traits. LWT-Food Sci. Technol. 2015, 64, 1156–1162. [Google Scholar] [CrossRef]
- Langa, S.; Peirotén, A.; Gaya, P.; Garde, S.; Arqués, J.L.; Nuñez, M.; Medina, M.; Rodríguez-Mínguez, E. Human Bifidobacterium strains as adjunct cultures in Spanish sheep milk cheese. J. Dairy Sci. 2020, 103, 7695–7706. [Google Scholar] [CrossRef] [PubMed]
- del Olmo, A.; Picon, A.; Nuñez, M. Cheese supplementation with five species of edible seaweeds: Effect on microbiota, antioxidant activity, colour, texture and sensory characteristics. Int. Dairy J. 2018, 84, 36–45. [Google Scholar] [CrossRef]
- Bárcenas, P.; Pérez Elortondo, F.J.; Albisu, M. Selection and screening of a descriptive panel for ewes milk cheese sensory profiling. J. Sens. Stud. 2000, 15, 79–99. [Google Scholar] [CrossRef]
- Caporaso, N.; Whitworth, M.B.; Grebby, S.; Fisk, I.D. Non-destructive analysis of sucrose, caffeine and trigonelline on single green coffee beans by hyperspectral imaging. Food Res. Int. 2018, 106, 193–203. [Google Scholar] [CrossRef]
- Manasa, V.; Padmanabhan, A.; Anu Appaiah, K.A. Utilization of coffee pulp waste for rapid recovery of pectin and polyphenols for sustainable material recycle. Waste Manag. 2021, 120, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Portela, C.d.S.; de Almeida, I.F.; Mori, A.L.B.; Yamashita, F.; Benassi, M.d.T. Brewing conditions impact on the composition and characteristics of cold brew Arabica and Robusta coffee beverages. LWT 2021, 143, 111090. [Google Scholar] [CrossRef]
- Heeger, A.; Kosińska-Cagnazzo, A.; Cantergiani, E.; Andlauer, W. Bioactives of coffee cherry pulp and its utilisation for production of Cascara beverage. Food Chem. 2017, 221, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Duangjai, A.; Suphrom, N.; Wungrath, J.; Ontawong, A.; Nuengchamnong, N.; Yosboonruang, A. Comparison of antioxidant, antimicrobial activities and chemical profiles of three coffee (Coffea arabica L.) pulp aqueous extracts. Integr. Med. Res. 2016, 5, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Garrett, R.; Vaz, B.G.; Hovell, A.M.C.; Eberlin, M.N.; Rezende, C.M. Arabica and Robusta Coffees: Identification of major polar compounds and quantification of blends by direct-infusion electrospray ionization-mass spectrometry. J. Agric. Food Chem. 2012, 60, 4253–4258. [Google Scholar] [CrossRef]
- Ashihara, H. Chapter 3—Plant Biochemistry: Trigonelline Biosynthesis in Coffea arabica and Coffea canephora. In Coffee in Health and Disease Prevention; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2015; pp. 19–28. [Google Scholar]
- Membrez, M.; Migliavacca, E.; Christen, S.; Yaku, K.; Trieu, J.; Lee, A.K.; Morandini, F.; Giner, M.P.; Stiner, J.; Makarov, M.V.; et al. Trigonelline is an NAD+ precursor that improves muscle function during ageing and is reduced in human sarcopenia. Nat. Metab. 2024, 6, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Martinez, J.R. Phenolic compounds in coffee pulp: Quantitative determination by HPLC. J. Sci. Food Agric. 1988, 43, 135–144. [Google Scholar] [CrossRef]
- Rodríguez-Daza, M.C.; Pulido-Mateos, E.C.; Lupien-Meilleur, J.; Guyonnet, D.; Desjardins, Y.; Roy, D. Polyphenol-Mediated Gut Microbiota Modulation: Toward Prebiotics and Further. Front. Nutr. 2021, 8, 689456. [Google Scholar] [CrossRef]
- Sun, C.Q.; O’Connor, C.J.; Turner, S.J.; Lewis, G.D.; Stanley, R.A.; Roberton, A.M. The effect of pH on the inhibition of bacterial growth by physiological concentrations of butyric acid: Implications for neonates fed on suckled milk. Chem.-Biol. Interact. 1998, 113, 117–131. [Google Scholar] [CrossRef]
- Pereira, G.V.D.M.; de Carvalho Neto, D.P.; Medeiros, A.B.P.; Soccol, V.T.; Neto, E.; Woiciechowski, A.L.; Soccol, C.R. Potential of lactic acid bacteria to improve the fermentation and quality of coffee during on-farm processing. Int. J. Food Sci. Technol. 2016, 51, 1689–1695. [Google Scholar] [CrossRef]
- Muñoz, R.; de las Rivas, B.; Rodríguez, H.; Esteban-Torres, M.; Reverón, I.; Santamaría, L.; Landete, J.M.; Plaza-Vinuesa, L.; Sánchez-Arroyo, A.; Jiménez, N.; et al. Food phenolics and Lactiplantibacillus plantarum. Int. J. Food Microbiol. 2024, 412, 110555. [Google Scholar] [CrossRef] [PubMed]
- Peirotén, A.; Álvarez, I.; Landete, J.M. Production of flavonoid and lignan aglycones from flaxseed and soy extracts by Bifidobacterium strains. Int. J. Food Sci. Technol. 2020, 5, 2122–2131. [Google Scholar] [CrossRef]
- Schulze, A.; Mitterer, F.; Pombo, J.P.; Schild, S. Biofilms by bacterial human pathogens: Clinical relevance—Development, composition and regulation—Therapeutical strategies. Microb. Cell 2021, 8, 28–56. [Google Scholar] [CrossRef] [PubMed]
- Terraf, M.C.L.; Juárez Tomás, M.S.; Nader-Macías, M.E.F.; Silva, C. Screening of biofilm formation by beneficial vaginal lactobacilli and influence of culture media components. J. Appl. Microbiol. 2012, 113, 1517–1529. [Google Scholar] [CrossRef]
- Yamanaka, A.; Kimizuka, R.; Kato, T.; Okuda, K. Inhibitory effects of cranberry juice on attachment of oral streptococci and biofilm formation. Oral Microbiol. Immunol. 2004, 19, 150–154. [Google Scholar] [CrossRef]
- Rathi, B.; Gupta, S.; Kumar, P.; Kesarwani, V.; Dhanda, R.S.; Kushwaha, S.K.; Yadav, M. Anti-biofilm activity of caffeine against uropathogenic E. coli is mediated by curli biogenesis. Sci. Rep. 2022, 12, 18903. [Google Scholar] [CrossRef]
- Amaretti, A.; di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef]
- Wastyk, H.C.; Fragiadakis, G.K.; Perelman, D.; Dahan, D.; Merrill, B.D.; Yu, F.B.; Topf, M.; Gonzalez, C.G.; Van Treuren, W.; Han, S.; et al. Gut-microbiota-targeted diets modulate human immune status. Cell 2021, 184, 4137–4153. [Google Scholar] [CrossRef]
- Saygili, S.; Hegde, S.; Shi, X.-Z. Effects of Coffee on Gut Microbiota and Bowel Functions in Health and Diseases: A Literature Review. Nutrients 2024, 16, 3155. [Google Scholar] [CrossRef]
- Karimi, R.; Mortazavian, A.M.; Da Cruz, A.G. Viability of probiotic microorganisms in cheese during production and storage: A review. Dairy Sci. Technol. 2011, 91, 283–308. [Google Scholar] [CrossRef]
- Chan, M.Z.A.; Toh, M.; Liu, S.-Q. Growth, survival, and metabolic activities of probiotics Lactobacillus rhamnosus GG and Saccharomyces cerevisiae var. boulardii CNCM-I745 in fermented coffee brews. Int. J. Food Microbiol. 2021, 350, 109229. [Google Scholar] [CrossRef] [PubMed]
- Vinderola, G.; Zacarias, M.F.; Bockelmann, W.; Neve, H.; Reinheimer, J.; Heller, K.J. Preservation of functionality of Bifidobacterium animalis subsp. lactis INL1 after incorporation of freeze-dried cells into different food matrices. Food Microbiol. 2012, 30, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Ranadheera, R.D.C.S.; Baines, S.K.; Adams, M.C. Importance of food in probiotic efficacy. Food Res. Int. 2010, 43, 1–7. [Google Scholar] [CrossRef]
- Hussain, N.; Li, R.; Takala, T.M.; Tariq, M.; Zaidi, A.H.; Saris, P.E.J. Generation of lactose- and protease-positive probiotic Lacticaseibacillus rhamnosus GG by conjugation with Lactococcus lactis NCDO 712. Appl. Environ. Microbiol. 2021, 87, 02957-20. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, H.; Karatas, N. Microbial exopolysaccharides: Resources and bioactive properties. Process Biochem. 2018, 72, 41–46. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Variable | Extracts | |||
|---|---|---|---|---|
| Whole Pulp | Ground Pulp | Ground Pulp 4 °C | Bean + Silverskin | |
| pH | 3.78 ± 0.02 b | 3.76 ± 0.01 b | 3.74 ± 0.01 b | 5.41 ± 0.01 a |
| A600 | 0.0268 ± 0.0100 bc | 0.0460 ± 0.0093 b | 0.0193 ± 0.0102 c | 0.1352 ± 0.0098 a |
| TPC 2 | 136.15 ± 1.16 c | 314.18 ± 0.61 a | 160.48 ± 0.45 b | 70.36 ± 0.42 d |
| Strain | Control | Extract | |||
|---|---|---|---|---|---|
| Whole Pulp | Ground Pulp | Ground Pulp 4 °C | Bean + Silver Skin | ||
| INIA P495 | 1+ | 3 | 4 | 4 | 2 |
| INIA P708 | 1+ | 3 | 4 | 4 | 2 |
| INIA P334 | 1 | 2 | 3 | 3 | 1+ |
| INIA TAB84 | 1 | 2 | 3 | 3 | 1 |
| INIA P508 | 1 | 2 | 3 | 3 | 1 |
| INIA P459 | 1 | 3 | 3 | 3 | 1 |
| GG | 1+ | 3 | 4 | 4 | 1+ |
| pH | A600 | DM 1 (%) | Sugar Content (mg GE/mL) | Protein Content (μg BSA/mL) | TPC 2 (μg GAE/mL) | Antioxidant Capacity (mM TE) |
|---|---|---|---|---|---|---|
| 3.81 ± 0.01 | 0.0451 ± 0.01 | 1.98 ± 0.03 | 8.28 ± 0.21 | 47.39 ± 4.37 | 406.43 ± 1.50 | 8.36 ± 0.09 |
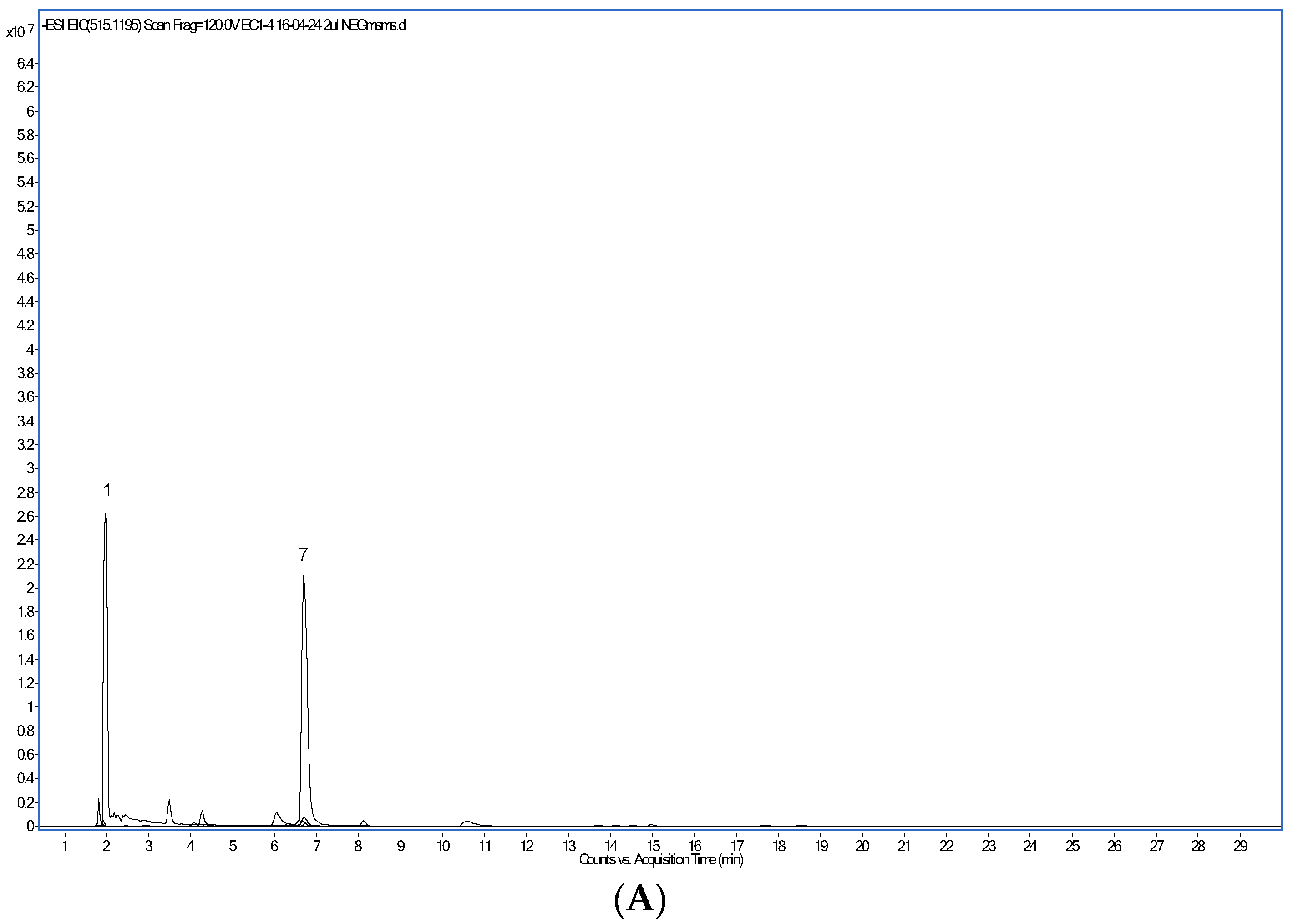
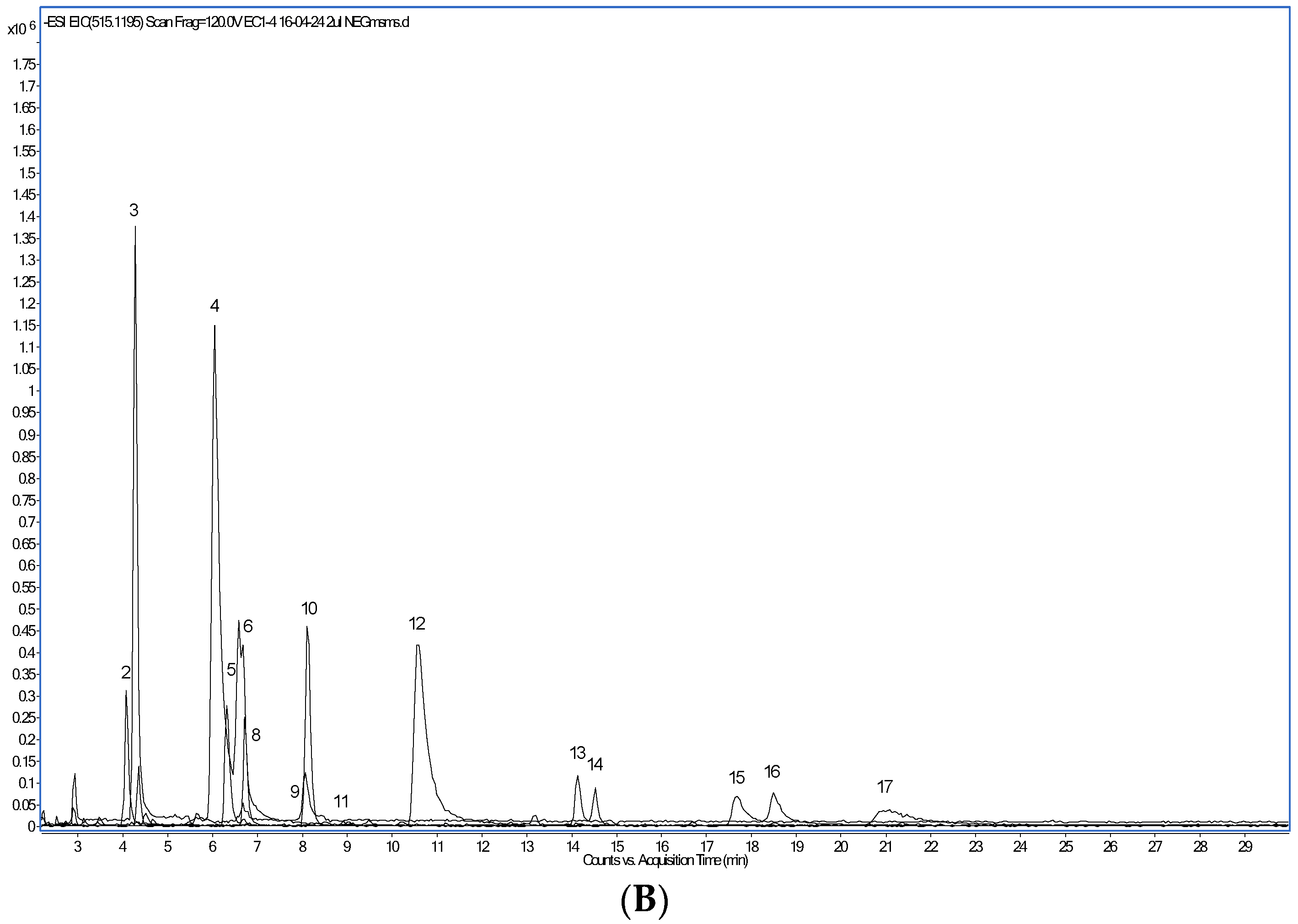
| Peak Number | Retention Time (min) | Compound | [M-H]- m/z | MS2 Ions m/z |
|---|---|---|---|---|
| 1 | 1.9 | Trigonelline | 138.0550 * | 138 (100), 94 (63), 53 (27), 78 (18), 65 (13) |
| 2 | 4.1 | 3-Caffeoylquinic acid | 353.0878 | 191 (100), 135 (35) |
| 3 | 4.4 | Protocatechuic acid | 153.0193 | 109 (100) |
| 4 | 6.1 | Chlorogenic acid (5-Caffeoylquinic acid) | 353.0878 | 191 (100) |
| 5 | 6.3 | Catechin | 289.0718 | 203 (100), 245 (87), 109 (65), 221 (64), 164 (57), 123 (45), 227 (38), 97 (37), 150 (36), 80 (32), 186 (32), 138 (31), 211 (25), 175 (23) |
| 6 | 6.6 | 4-Caffeoylquinic acid | 353.0878 | 191 (100), 173 (100), 135 (34) |
| 7 | 6.7 | Caffeine (1,3,7-Trimethylxanthine) | 195.0877 * | 138 (100), 195 (41), 110 (19) |
| 8 | 6.7 | 3-Feruloylquinic acid | 367.1035 | 193 (100), 134 (19) |
| 9 | 8.1 | Caffeic acid | 179.0350 | 135 (100), 105 (12), 93 (12) |
| 10 | 8.1 | 5-Feruloylquinic acid | 367.1035 | 367 (100) |
| 11 | 8.9 | Epicatechin | 289.0718 | 203 (100), 245 (87), 109 (65), 221 (64), 123 (45), 227 (38), 97 (37), 150 (35), 186 (32), 80 (32), 289 (31), 138 (31), 211 (25) |
| 12 | 10.5 | 4-Feruloylquinic acid | 367.1035 | 191 (100), 173 (24) |
| 13 | 14.1 | Rutin (Quercetin-3-O-rutinoside) | 609.1461 | 609 (100), 300 (11) |
| 14 | 14.5 | 609.1461 | 609 (100), 300 (17) | |
| 15 | 17.7 | 3,4-di-O-caffeoylquinic acid | 515.1195 | 173 (100), 353 (83), 515 (37), 335 (19), 155 (13) |
| 16 | 18.5 | 3,5-di-O-caffeoylquinic acid | 515.1195 | 353 (100), 191 (71), 179 (14), 509 (10) |
| 17 | 21.1 | 4,5-di-O-caffeoylquinic acid | 515.1195 | 353 (100), 173 (69), 311 (14), 516 (10), 263 (10) |
| Strain | A600 | |
|---|---|---|
| BHICO | BHICO + 40% CPE | |
| INIA P495 | 0.77 ± 0.12 a A 0.03 ± 0.01 a D 0.13 ± 0.02 b C 0.39 ± 0.09 b B | 0.97 ± 0.17 a B 0.04 ± 0.02 a C 0.19 ± 0.04 a C 1.26 ± 0.29 a A |
| INIA P708 | ||
| INIA P459 | ||
| GG | ||
| Strain | Time | Percentage of DPPH Reduction | |
|---|---|---|---|
| Control BHIC | BHIC +40% CPE | ||
| INIA P495 | 1 d | 14.70 ± 1.02 b A | 45.90 ± 1.53 a B |
| 6 d | 16.31 ± 0.65 b A | 65.13 ± 0.52 a A | |
| INIA P708 | 1 d | 10.61 ± 1.13 b B | 30.67 ± 1.05 a B |
| 6 d | 18.20 ± 0.57 b A | 54.38 ± 0.52 a A | |
| INIA P459 | 1 d | 12.45 ± 0.86 b A | 39.27 ± 2.64 a B |
| 6 d | 13.92 ± 0.42 b A | 62.16 ± 0.18 a A | |
| GG | 1 d | 12.78 ± 0.67 b B | 53.10 ± 3.72 a A |
| 6 d | 16.75 ± 0.67 b A | 54.53 ± 0.47 a A | |
| Strain | Control BHIC | BHIC + 40% CPE | ||||||
|---|---|---|---|---|---|---|---|---|
| C | K | L | S | C | K | L | S | |
| INIA P495 | - | - | + | - | - | D | +++ | + |
| INIA P708 | - | D | + | - | ++ | ++ | +++ | ++ |
| INIA P459 | D | - | - | - | D | - | + | - |
| GG | - | D | + | - | + | D | +++ | + |
| Strain | Time | Levels in Fermented Milk | Levels After Major GIT Conditions | ||
|---|---|---|---|---|---|
| M100 | M80:CPE20 | M100 | M80:CPE20 | ||
| INIA P495 | 1 d | 8.18 ± 0.08 a C | 8.30 ± 0.24 a B | 8.45 ± 0.12 a A | 8.45 ± 0.20 a A |
| 12 d | 8.38 ± 0.07 a B | 8.21 ± 0.08 a B | 7.71 ± 0.09 a A | 7.73 ± 0.05 a B | |
| 20 d | 8.74 ± 0.06 a A | 8.71 ± 0.10 a A | 8.51 ± 0.19 a A | 8.75 ± 0.38 a A | |
| INIA P708 | 1 d | 9.18 ± 0.01 a B | 9.27 ± 0.13 a A | 8.75 ± 0.16 a B | 8.69 ± 0.14 a B |
| 12 d | 9.42 ± 0.05 a A | 9.31 ± 0.24 a A | 9.33 ± 0.04 a A | 9.29 ± 0.06 a A | |
| 20 d | 9.09 ± 0.15 a B | 9.23 ± 0.03 a A | 9.32 ± 0.00 a A | 9.28 ± 0.04 a A | |
| GG | 1 d | 8.51 ± 0.13 a A | 8.57 ± 0.24 a A | 8.26 ± 0.32 a A | 8.26 ± 0.12 a A |
| 12 d | 8.51 ± 0.16 a A | 8.57 ± 0.12 a A | 8.08 ± 0.00 a A | 7.71 ± 0.07 a B | |
| 20 d | 8.29 ± 0.27 a A | 8.49 ± 0.04 a A | 7.91 ± 0.06 a A | 8.19 ± 0.14 a A | |
| Variable | Strain | Time | M100 | M80:CPE20 |
|---|---|---|---|---|
| pH | Non inoculated | 0 h | 6.58 ± 0.00 a C | 6.31 ± 0.01 b C |
| 1 d | 6.51 ± 0.01 a D | 6.21 ± 0.00 b D | ||
| 12 d | 6.61 ± 0.01 a B | 6.38 ± 0.01 b B | ||
| 20 d | 6.67 ± 0.00 a A | 6.42 ± 0.01 b A | ||
| INIA P495 | 1 d | 6.15 ± 0.03 a A | 5.26 ± 0.02 b A | |
| 12 d | 6.11 ± 0.03 a A | 4.89 ± 0.01 b B | ||
| 20 d | 5.92 ± 0.03 a B | 4.46 ± 0.01 b C | ||
| INIA P708 | 1 d | 5.16 ± 0.02 a A | 4.34 ± 0.00 b A | |
| 12 d | 4.84 ± 0.02 a B | 4.01 ± 0.01 b B | ||
| 20 d | 4.56 ± 0.02 a C | 3.89 ± 0.01 b C | ||
| GG | 1 d | 6.12 ± 0.02 a C | 5.64 ± 0.01 b B | |
| 12 d | 6.30 ± 0.01 a B | 5.81 ± 0.01 b A | ||
| 20 d | 6.35 ± 0.01 a A | 5.84 ± 0.02 b A | ||
| TPC | Non inoculated | 0 h | 155.45 ± 10.29 b AB | 213.57 ± 8.64 a B |
| 1 d | 157.97 ± 1.59 b A | 210.67 ± 4.82 a B | ||
| 12 d | 143.13 ± 1.82 b BC | 199.54 ± 2.41 a B | ||
| 20 d | 141.79 ± 5.95 b C | 231.79 ± 10.90 a A | ||
| INIA P495 | 1 d | 150.82 ± 2.25 b B | 221.37 ± 1.89 a B | |
| 12 d | 150.23 ± 2.95 b B | 214.11 ± 5.67 a B | ||
| 20 d | 159.70 ± 7.34 b A | 269.27 ± 4.15 a A | ||
| INIA P708 | 1 d | 170.99 ± 3.44 b C | 215.21 ± 2.15 a B | |
| 12 d | 185.99 ± 3.40 b B | 214.97 ± 2.36 a B | ||
| 20 d | 223.85 ± 9.11 b A | 280.07 ± 12.02 a A | ||
| GG | 1 d | 146.84 ± 1.48 b A | 215.77 ± 4.05 a B | |
| 12 d | 145.77 ± 2.93 b A | 209.64 ± 2.82 a B | ||
| 20 d | 145.49 ± 8.73 b A | 254.50 ± 7.32 a A | ||
| Antioxidant | Non inoculated | 0 h | 0.71 ± 0.02 b B | 1.76 ± 0.02 a A |
| capacity | 1 d | 0.67 ± 0.03 b B | 1.69 ± 0.06 a B | |
| 12 d | 0.70 ± 0.02 b B | 1.54 ± 0.01 a C | ||
| 20 d | 0.79 ± 0.01 b A | 1.64 ± 0.03 a B | ||
| INIA P495 | 1 d | 0.64 ± 0.02 b C | 1.67 ± 0.04 a AB | |
| 12 d | 0.72 ± 0.01 b B | 1.62 ± 0.04 a B | ||
| 20 d | 0.75 ± 0.01 b A | 1.69 ± 0.02 a A | ||
| INIA P708 | 1 d | 0.67 ± 0.02 b C | 1.72 ± 0.04 a B | |
| 12 d | 0.82 ± 0.02 b B | 1.65 ± 0.01 a B | ||
| 20 d | 0.97 ± 0.04 b A | 1.81 ± 0.04 a A | ||
| GG | 1 d | 0.65 ± 0.03 b B | 1.68 ± 0.01 a A | |
| 12 d | 0.68 ± 0.00 b AB | 1.60 ± 0.02 a A | ||
| 20 d | 0.72 ± 0.02 b A | 1.65± 0.06 a A |
| Product | Aspect | Odour | Flavour |
|---|---|---|---|
| M100 | white, liquid | lactic | lactic |
| M80:20CPE | colour slightly darker than milk, liquid | slightly herbal | aromatic, slightly sweet and astringent, CPE taste |
| M80:20CPE + INIA P495 | colour similar to milk, liquid | yogurt, sweet, toasted, caramel | sweet and toasted. Less astringent and acid, and more herbal than M100:20CPE. |
| M80:20CPE + INIA P708 | colour slightly darker than milk, coagulated, viscous, creamy | yogurt, not herbal | acid, lactic, final astringency |
| M80:20CPE + GG | colour similar to milk, liquid | mild yogurt | similar to M100:20CPE |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Picon, A.; Campanero, Y.; Sánchez, C.; Álvarez, I.; Rodríguez-Mínguez, E. Valorization of Coffee Cherry By-Products Through Fermentation by Human Intestinal Lactobacilli in Functional Fermented Milk Beverages. Foods 2025, 14, 44. https://doi.org/10.3390/foods14010044
Picon A, Campanero Y, Sánchez C, Álvarez I, Rodríguez-Mínguez E. Valorization of Coffee Cherry By-Products Through Fermentation by Human Intestinal Lactobacilli in Functional Fermented Milk Beverages. Foods. 2025; 14(1):44. https://doi.org/10.3390/foods14010044
Chicago/Turabian StylePicon, Antonia, Yolanda Campanero, Carmen Sánchez, Inmaculada Álvarez, and Eva Rodríguez-Mínguez. 2025. "Valorization of Coffee Cherry By-Products Through Fermentation by Human Intestinal Lactobacilli in Functional Fermented Milk Beverages" Foods 14, no. 1: 44. https://doi.org/10.3390/foods14010044
APA StylePicon, A., Campanero, Y., Sánchez, C., Álvarez, I., & Rodríguez-Mínguez, E. (2025). Valorization of Coffee Cherry By-Products Through Fermentation by Human Intestinal Lactobacilli in Functional Fermented Milk Beverages. Foods, 14(1), 44. https://doi.org/10.3390/foods14010044