Comprehensive and Comparative Metabolomic Profiling of Wheat, Barley, Oat and Rye Using Gas Chromatography-Mass Spectrometry and Advanced Chemometrics



Abstract
:1. Introduction
2. Experimental Section
2.1. Metabolite Extraction and Sample Derivatization
2.2. GC-MS Data Acquisition
2.3. Data Analysis
3. Results and Discussion
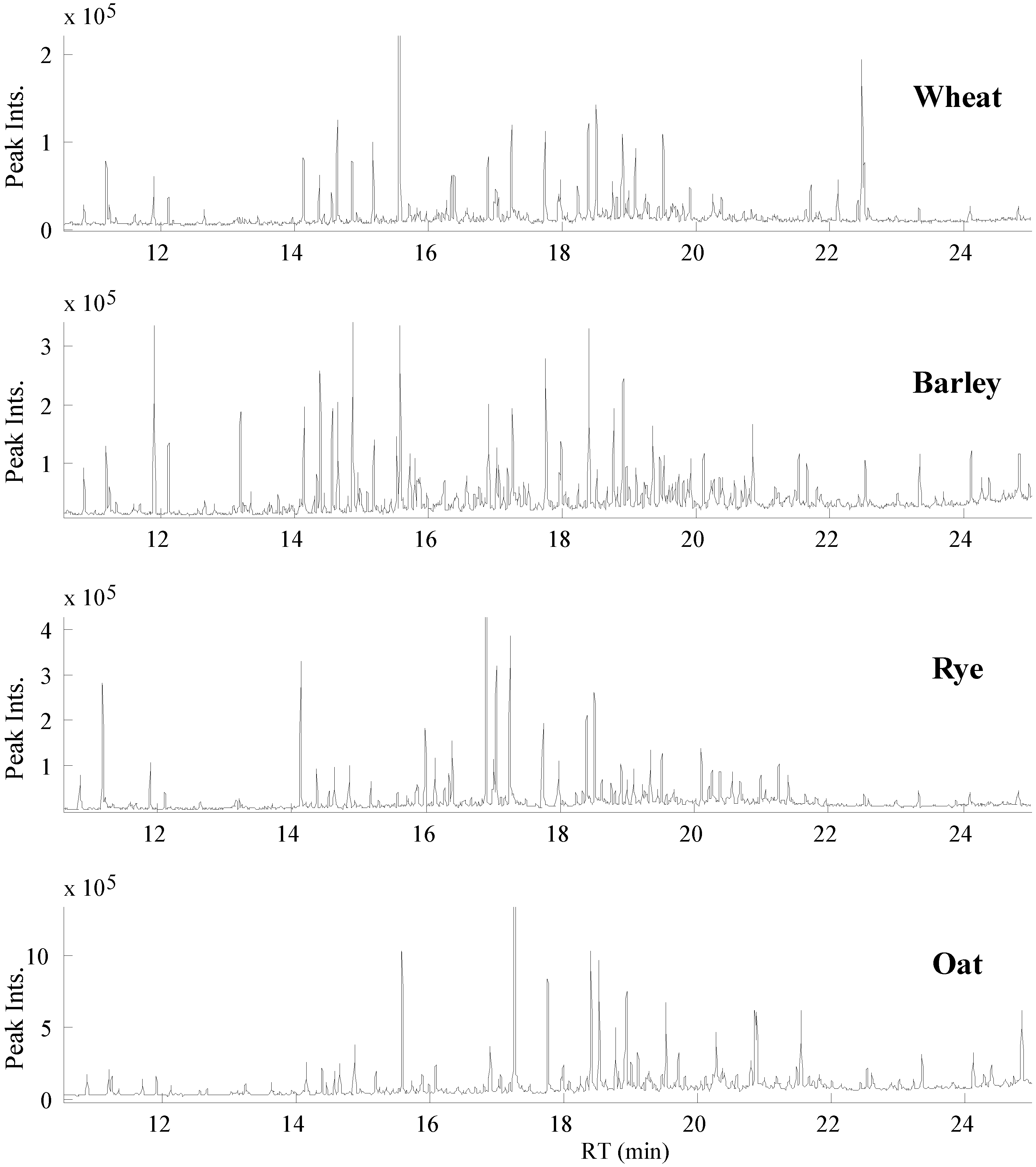
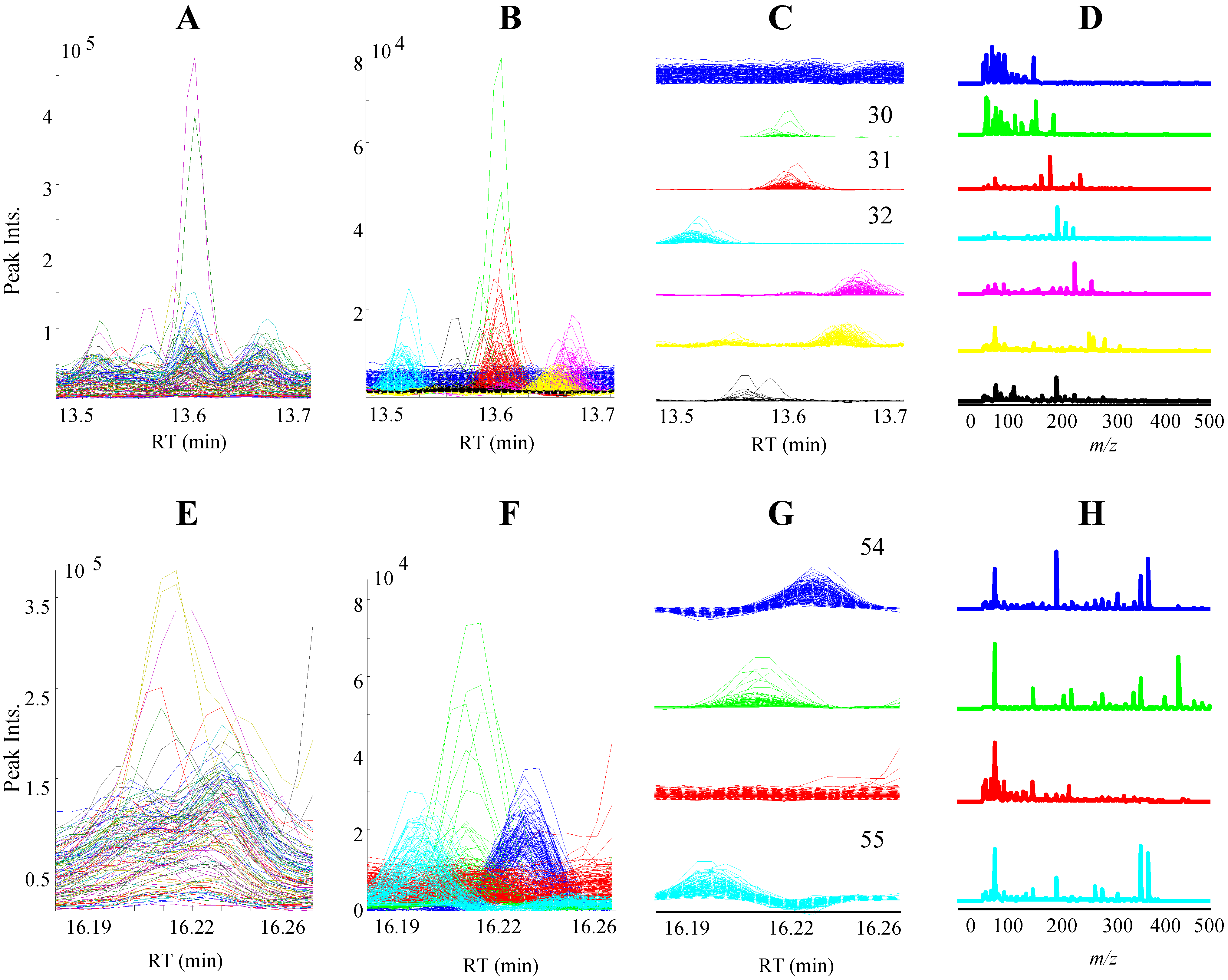
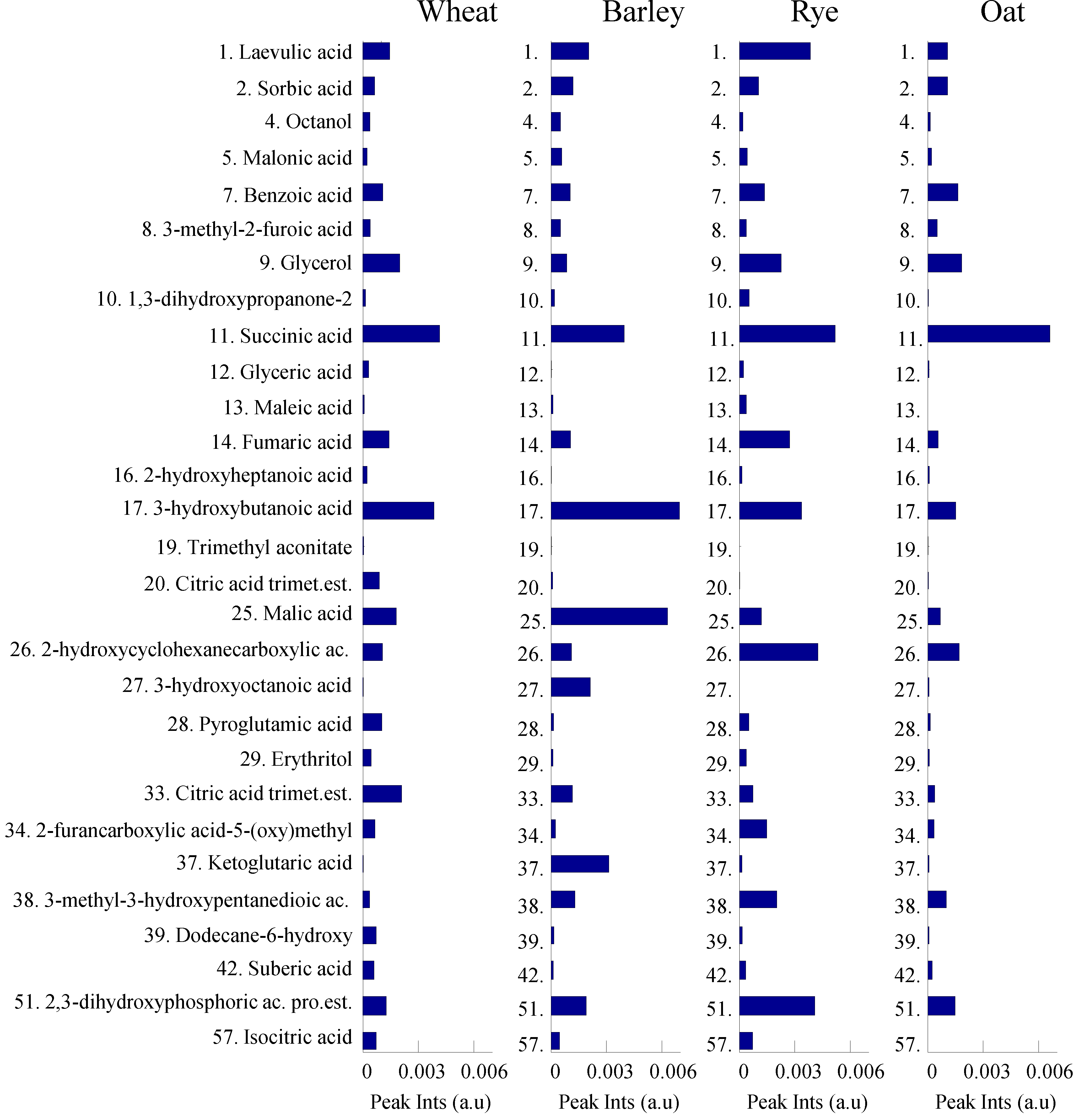
3.1. GC-MS Metabolomic Profiling and PARAFAC2 Based Data Processing



{kind=link}
{kind=link}
{kind=link}
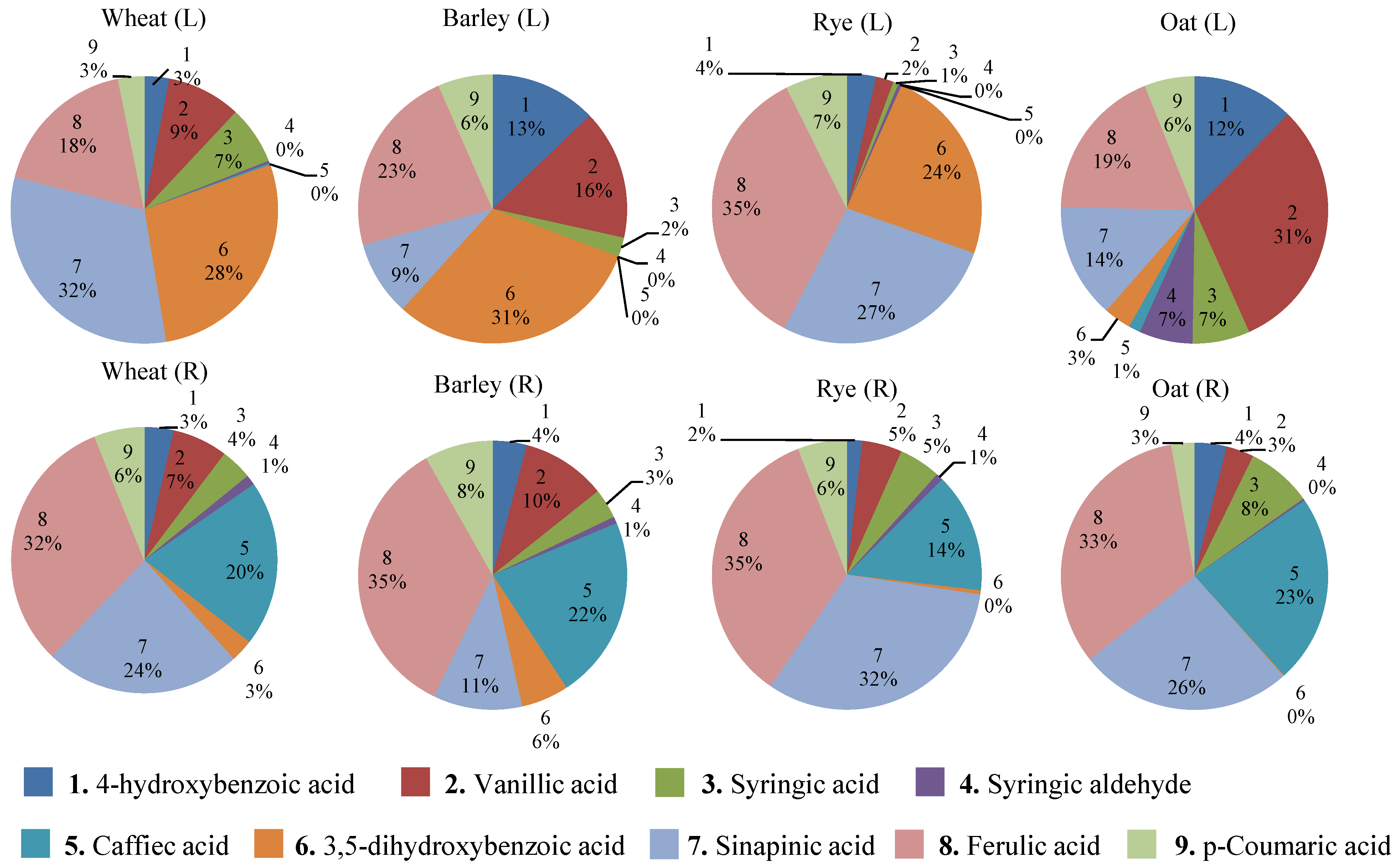{kind=link}
{kind=link}
{kind=link}
| No | Metabolites | RT min | RI (r) | RI (c) |
|---|---|---|---|---|
| 1. | Laevulic acid-1TMS | 9.04 | 1030 | 1070 |
| 2. | Sorbic acid-1TMS | 9.06 | 1009 | 1071 |
| 3. | Hepta-2,4-dienoic acid, methyl ester | 9.28 | 1000 | 1080 |
| 4. | Octanol-1-1TMS | 9.51 | 1101 | 1090 |
| 5. | Malonic acid-2TMS | 9.99 | 1205 | 1207 |
| 6. | (3,3-Dimethyl-1-cyclohexen-1-yl)oxy]-1TMS | 9.97 | 1110 | 1206 |
| 7. | Benzoic acid-1TMS | 10.42 | 1228 | 1226 |
| 8. | 3-Methyl-2-furoic acid-1TMS | 10.38 | 1107 | 1224 |
| 9. | Glycerol-3TMS | 10.88 | 1282 | 1246 |
| 10. | 1,3-Dihydroxypropanone-2-2TMS | 11.03 | 1249 | |
| 11. | Succinic acid-2TMS | 11.24 | 1292 | 1262 |
| 12. | Glyceric acid-3TMS | 11.51 | 1199 | 1274 |
| 13. | Maleic acid-2TMS | 11.55 | 1286 | 1275 |
| 14. | Fumaric acid-2TMS | 11.60 | 1178 | 1278 |
| 15. | p-Hydroxybenzaldehyde-1TMS | 11.85 | 1280 | 1289 |
| 16. | 2-Hydroxyheptanoic acid-2TMS | 11.83 | 1312 | 1288 |
| 17. | 3-Hydroxybutanoic acid-2TMS | 12.12 | 1403 | 1401 |
| 18. | Resorcinol-2TMS | 12.2 | 1378 | 1404 |
| 19. | Trimethyl aconitate | 12.50 | 1428 | 1419 |
| 20. | Citric acid, trimethyl ester | 12.82 | 1442 | 1435 |
| 21. | 3-Hydroxyanthranilic acid, methyl ester-1TMS | 12.8 | 1434 | |
| 22. | 2,4-Dihydroxy-5-methylpyrimidine-2TMS | 12.89 | 1403 | 1439 |
| 23. | 5-Hydroxy-2-(hydroxymethyl)-4H-pyran-4-one-2TMS | 13.08 | 1492 | 1448 |
| 24. | Maseptol-1TMS | 13.12 | 1358 | 1450 |
| 25. | Malic acid-2TMS | 13.19 | 1494 | 1453 |
| 26. | 2-Hydroxycyclohexanecarboxylic acid-2TMS | 13.23 | 1402 | 1456 |
| 27. | 3-Hydroxyoctanoic acid-2TMS | 13.35 | 1452 | 1462 |
| 28. | Pyroglutamic acid-2TMS | 13.46 | 1466 | 1467 |
| 29. | Erythritol-4TMS | 13.47 | 1467 | |
| 30. | Dimethyl azelate | 13.61 | 1485 | 1474 |
| 31. | 4-Hydroxybenzeneacetic acid, methyl ester-1TMS | 13.62 | 1458 | 1475 |
| 32. | Vanillin-1TMS | 13.55 | 1469 | 1471 |
| 33. | Citric acid, trimethyl ester-1TMS | 13.76 | 1482 | |
| 34. | 2-Furancarboxylic acid, 5-[(oxy)methyl]-1TMS | 13.72 | 1540 | 1480 |
| 35. | 4-Hydroxyphenylethanol-2TMS | 13.92 | 1475 | 1490 |
| 36. | Anozol | 14.15 | 1603 | 1601 |
| 37. | 2-Ketoglutaric acid-3TMS | 14.34 | 1622 | 1612 |
| 38. | 3-Methyl-3-hydroxypentanedioic acid-3TMS | 14.3 | 1610 | 1609 |
| 39. | Dodecane-6-hydroxy-1TMS | 14.40 | 1631 | 1615 |
| 40. | 4-Hydroxybenzoic acid-2TMS | 14.45 | 1618 | 1618 |
| 41. | Methyl Isovanillate-1TMS | 14.66 | 1547 | 1629 |
| 42. | Suberic acid-2TMS | 15.11 | 1682 | 1654 |
| 43. | Syringaldehyde -1TMS | 15.15 | 1658 | 1656 |
| 44. | β-d-Arabinopyranose-4TMS a | 15.23 | 1692 | 1660 |
| 45. | β-d-Xylopyranose-4TMS | 15.30 | 1694 | 1664 |
| 46. | 3,5-Dihydroxybenzoic ac. met.est.-2TMS | 15.35 | 1656 | 1667 |
| 47. | 2,5-Dimethoxymandelic acid-2TMS | 15.38 | 1867 | 1669 |
| 48. | Vanillic acid-2TMS | 15.72 | 1656 | 1687 |
| 49. | 4-Hydroxycinnamic acid, methyl ester -1TMS | 15.88 | 1565 | 1696 |
| 50. | Azelaic acid-2TMS | 15.98 | 1800 | 1802 |
| 51. | 2,3-Dihydroxyphosphoric acid, propyl ester-4TMS | 15.86 | 1708 | 1695 |
| 52. | Methyl 2-(oxy)-2-(4-(oxy)phenyl)propanoate-2TMS | 16.14 | 1757 | 1811 |
| 53. | α-d-Galactofuranoside, methyl-2,3,5,6-tetrakis-4TMS a | 16.11 | 1845 | 1810 |
| 54. | 3,5-Dihydroxy benzoic ac.-3TMS | 16.24 | 1826 | 1818 |
| 55. | 3,4-Dihydroxy benzoic ac.-3TMS | 16.20 | 1826 | 1815 |
| 56. | d-Fructose-5TMS | 16.41 | 1867 | 1828 |
| 57. | Isocitric acid-4TMS | 16.34 | 1835 | 1823 |
| 58. | Catechin-nTMS a | 16.44 | 1830 | |
| 59. | Homovanilic acid-2TMS | 16.4 | 1867 | 1827 |
| 60. | β-d-Galactopyranoside, methyl 2,3,4,6-tetrakis-4TMS a | 16.68 | 1900 | 1844 |
| 61. | Catechin-nTMS a | 16.77 | 1849 | |
| 62. | 2,5-Dihydroxy benzoic ac.-3TMS | 16.78 | 1796 | 1850 |
| 63. | α-d-Glucopyranoside, methyl 2,3,4,6-tetrakis-4TMS a | 16.90 | 1928 | 1857 |
| 64. | Syringic acid-2TMS | 16.88 | 1845 | 1856 |
| 65. | β-d-Glucopyranoside, methyl 2,3,4,6-tetrakis-4TMS a | 17.05 | 1928 | 1866 |
| 66. | α-d-Glucopyranose, 1,2,3,4,6-pentakis-5TMS a | 17.02 | 1924 | 1864 |
| 67. | Palmitic acid, methyl ester | 17.01 | 1870 | 1864 |
| 68. | d-Galactose, 2,3,4,5,6-pentakis-5TMS a | 17.12 | 1970 | 1871 |
| 69. | p-Coumaric acid-2TMS | 17.18 | 1924 | 1874 |
| 70. | Ferulic acid, methyl ester-1TMS | 17.25 | 1765 | 1878 |
| 71. | 3,4,5-Trihydrozy benzoic ac.-4TMS | 17.45 | 1976 | 1890 |
| 72. | 2-Hydroxymandelic acid, ethyl ester-2TMS | 17.34 | 1777 | 1884 |
| 73. | 4’-Cyclohexylacetophenone | 17.58 | 1703 | 1898 |
| 74. | Caffeic acid methyl ester-2TMS | 17.76 | 1863 | 2010 |
| 75. | β-d-Glucopyranose-5TMS a | 17.75 | 1970 | 2009 |
| 76. | 2-Hydroxysebacic acid-3TMS | 18.13 | 2059 | 2034 |
| 77. | Ferulic acid-2TMS | 18.40 | 2076 | 2052 |
| 78. | 8,11-Octadecadienoic acid, methyl ester | 18.35 | 2093 | 2049 |
| 79. | Sinapinic acid methyl ester-1TMS | 18.51 | 1943 | 2059 |
| 80. | Methyl vanillactate-2TMS | 18.55 | 2030 | 2062 |
| 81. | Caffeic acid-3TMS | 18.76 | 2114 | 2076 |
| 82. | 9-Methoxy-4α-methyl-2,3,7-trihydroxy-4,4a-dihydro-2H-benzo[c]chromen-6(3H)-one b | 18.85 | 2082 | |
| 83. | Linoleic acid-1TMS | 19.23 | 2202 | 2207 |
| 84. | 4,8-Dihydroxy-2-quinolinecarboxylic acid-3TMS | 19.46 | 2265 | 2224 |
| 85. | Sinapinic acid-2TMS | 19.52 | 2221 | 2228 |
| 86. | Androsterone type plant sterol b | 19.89 | 2254 | |
| 87. | 3-Hydroxyandrostan-17-one-1TMS | 19.98 | 2186 | 2261 |
| 88. | 19-Norandrosterone-3-TMS b | 20.36 | 2198 | 2288 |
| 89. | 9,10-Dihydroxystearic acid-3TMS | 20.87 | 2517 | 2426 |
| 90. | 3,7-di-Hydroxy-androstan-17-one-2TMS | 21.09 | 2432 | 2443 |
| 91. | 9,10-Dihydroxystearic acid, dimethyl ester-2TMS | 21.49 | 2784 | 2474 |
| 92. | 2,3-Dihydroxypalmitic acid, propyl ester-2TMS | 21.84 | 2581 | 2601 |
| 93. | 2-Deoxy-6-phosphogluconolactone-5TMS | 23.26 | 2820 | |
| 94. | 2-Hydroxytetracosanoic acid, methyl ester-1TMS | 23.69 | 2894 | 2858 |
| 95. | 3,7-Dihydroxycholest-5-ene-2TMS | 23.95 | 2900 | 2881 |
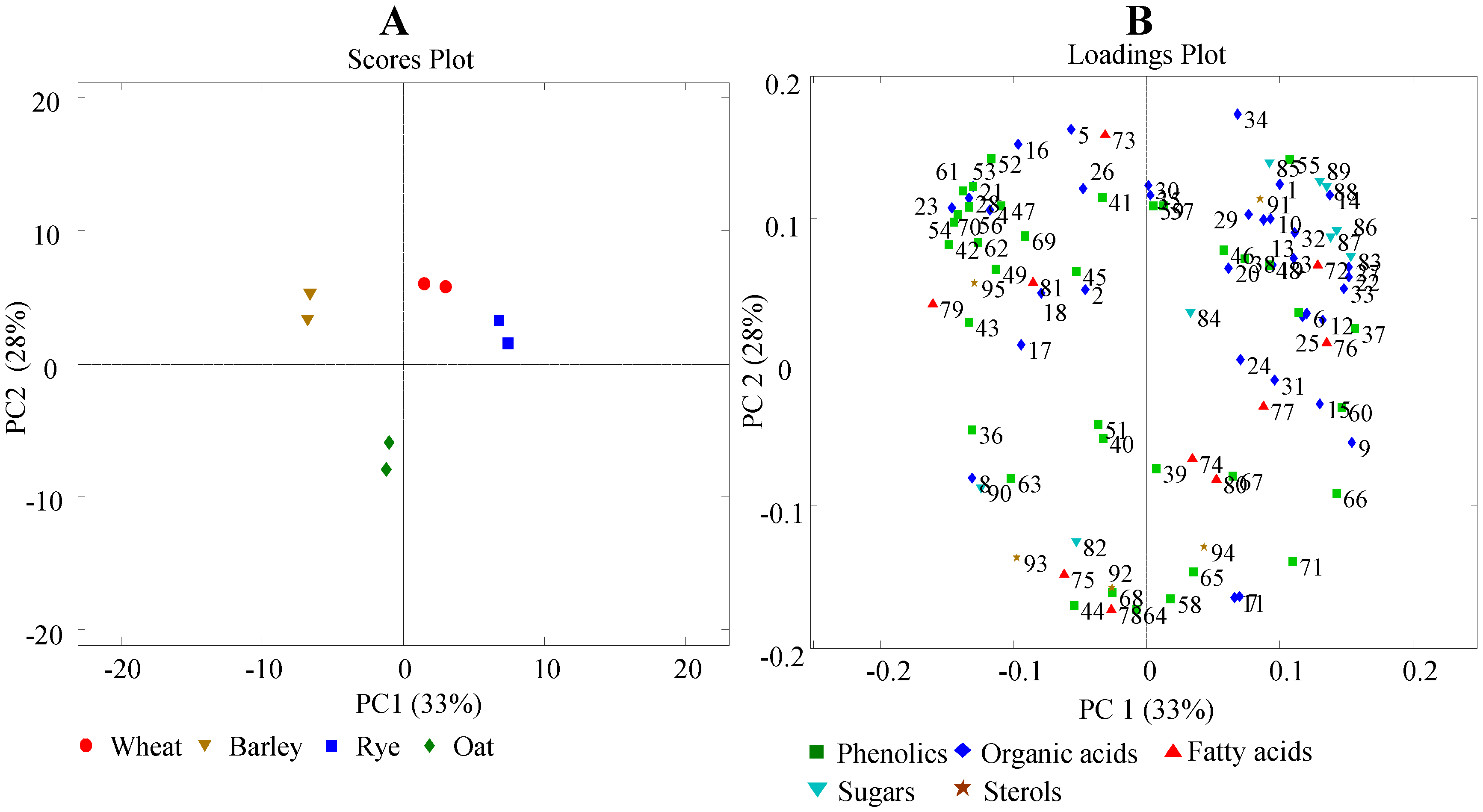
3.2. Principal Component Analysis (PCA)

3.3. Variation of Phenolics and Organic Acids in Cereals



4. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Zilic, S.; Sukalovic, V.H.T.; Dodig, D.; Maksimovic, V.; Maksimovic, M.; Basic, Z. Antioxidant activity of small grain cereals caused by phenolics and lipid soluble antioxidants. J. Cereal Sci 2011, 54, 417–424. [Google Scholar] [CrossRef]
- Björck, I.; Östman, E.; Kristensen, M.; Anson, N.M.; Price, R.K.; Haenen, G.R.M.M.; Havenaar, R.; Knudsen, K.E.B.; Frid, A.; Mykkänen, H.; et al. Cereal grains for nutrition and health benefits: Overview of results from in vitro, animal and human studies in the HEALTHGRAIN project. Trends Food Sci. Technol. 2012, 25, 87–100. [Google Scholar] [CrossRef]
- Andersson, A.A.M.; Andersson, R.; Piironen, V.; Lampi, A.M.; Nystrom, L.; Boros, D.; Fras, A.; Gebruers, K.; Courtin, C.M.; Delcour, J.A.; et al. Contents of dietary fibre components and their relation to associated bioactive components in whole grain wheat samples from the HEALTHGRAIN diversity screen. Food Chem. 2013, 136, 1243–1248. [Google Scholar] [CrossRef]
- Amarowicz, R.; Zegarska, Z.; Pegg, R.B.; Karamac, M.; Kosinska, A. Antioxidant and radical scavenging activities of a barley crude extract and its fractions. Czech J. Food Sci. 2007, 25, 73–80. [Google Scholar]
- Wood, P.J. Cereal beta-glucans in diet and health. J. Cereal Sci. 2007, 46, 230–238. [Google Scholar] [CrossRef]
- Mcintosh, G.H.; Whyte, J.; Mcarthur, R.; Nestel, P.J. Barley and wheat foods—Influence on plasma-cholesterol concentrations in hypercholesterolemic men. Am. J. Clin. Nutr. 1991, 53, 1205–1209. [Google Scholar]
- Madhujith, T.; Shahidi, F. Antioxidative and antiproliferative properties of selected barley (Hordeum vulgarae L.) cultivars and their potential for inhibition of low-density lipoprotein (LDL) cholesterol oxidation. J. Agric. Food Chem. 2007, 55, 5018–5024. [Google Scholar] [CrossRef]
- Behall, K.M.; Scholfield, D.J.; Hallfrisch, J. Diets containing barley significantly reduce lipids in mildly hypercholesterolemic men and women. Am. J. Clin. Nutr. 2004, 80, 1185–1193. [Google Scholar]
- Gibson, S.M.; Strauss, G. Implication of phenolic-acids as texturizing agents during cooking-extrusion cereals. Abstr. Pap. Am. Chem. Soc. 1991, 202, 150. [Google Scholar]
- Vinson, J.A.; Erk, K.M.; Wang, S.Y.; Marchegiani, J.Z.; Rose, M.F. Total polyphenol antioxidants in whole grain cereals and snacks: Surprising sources of antioxidants in the US diet. Abstr. Pap. Am. Chem. Soc. 2009, 238, 246. [Google Scholar]
- Khakimov, B.; Bak, S.; Engelsen, S.B. High-throughput cereal metabolomics: Current analytical technologies, challenges and perspectives. J. Cereal Sci. 2014, 59, 393–418. [Google Scholar] [CrossRef]
- Soltesz, A.; Smedley, M.; Vashegyi, I.; Galiba, G.; Harwood, W.; Vagujfalvi, A. Transgenic barley lines prove the involvement of TaCBF14 and TaCBF15 in the cold acclimation process and in frost tolerance. J. Exp. Bot. 2013, 64, 1849–1862. [Google Scholar] [CrossRef]
- Widodo; Patterson, J.H.; Newbigin, E.; Tester, M.; Bacic, A.; Roessner, U. Metabolic responses to salt stress of barley (Hordeum vulgare L.) cultivars, Sahara and Clipper, which differ in salinity tolerance. J. Exp. Bot. 2009, 60, 4089–4103. [Google Scholar] [CrossRef]
- Manavalan, L.P.; Chen, X.; Clarke, J.; Salmeron, J.; Nguyen, H.T. RNAi-mediated disruption of squalene synthase improves drought tolerance and yield in rice. J. Exp. Bot. 2012, 63, 163–175. [Google Scholar] [CrossRef]
- Balmer, D.; Flors, V.; Glauser, G.; Mauch-Mani, B. Metabolomics of cereals under biotic stress: Current knowledge and techniques. Front. Plant Sci. 2013, 4, 82. [Google Scholar]
- Fernie, A.R.; Schauer, N. Metabolomics-assisted breeding: A viable option for crop improvement? Trends Genet. 2009, 25, 39–48. [Google Scholar] [CrossRef]
- Bino, R.J.; Hall, R.D.; Fiehn, O.; Kopka, J.; Saito, K.; Draper, J.; Nikolau, B.J.; Mendes, P.; Roessner-Tunali, U.; Beale, M.H.; et al. Potential of metabolomics as a functional genomics tool. Trends Plant Sci. 2004, 9, 418–425. [Google Scholar] [CrossRef]
- Li, L.; Shewry, P.R.; Ward, J.L. Phenolic acids in wheat varieties in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9732–9739. [Google Scholar] [CrossRef]
- Fernandez-Orozco, R.; Li, L.; Harflett, C.; Shewry, P.R.; Ward, J.L. Effects of environment and genotype on phenolic acids in wheat in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2010, 58, 9341–9352. [Google Scholar] [CrossRef]
- Shewry, P.R.; Piironen, V.; Lampi, A.M.; Edelmann, M.; Kariluoto, S.; Nurmi, T.; Fernandez-Orozco, R.; Ravel, C.; Charmet, G.; Andersson, A.A.M.; et al. The HEALTHGRAIN wheat diversity screen: Effects of genotype and environment on phytochemicals and dietary fiber components. J. Agric. Food Chem. 2010, 58, 9291–9298. [Google Scholar] [CrossRef]
- Andersson, A.A.M.; Lampi, A.M.; Nystrom, L.; Piironen, V.; Li, L.; Ward, J.L.; Gebruers, K.; Courtin, C.M.; Delcour, J.A.; Boros, D.; et al. Phytochemical and dietary fiber components in barley varieties in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9767–9776. [Google Scholar] [CrossRef]
- Shewry, P.R.; Piironen, V.; Lampi, A.M.; Nystrom, L.; Li, L.; Rakszegi, M.; Fras, A.; Boros, D.; Gebruers, K.; Courtin, C.M.; et al. Phytochemical and fiber components in oat varieties in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9777–9784. [Google Scholar] [CrossRef]
- Nyström, L.; Lampi, A.M.; Andersson, A.A.M.; Kamal-Eldin, A.; Gebruers, K.; Courtin, C.M.; Delcour, J.A.; Li, L.; Ward, J.L.; Fras, A.; et al. Phytochemicals and dietary fiber components in rye varieties in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9758–9766. [Google Scholar] [CrossRef]
- Ward, J.L.; Poutanen, K.; Gebruers, K.; Piironen, V.; Lampi, A.M.; Nystrom, L.; Andersson, A.A.M.; Aman, P.; Boros, D.; Rakszegi, M.; et al. The HEALTHGRAIN cereal diversity screen: Concept, results, and prospects. J. Agric. Food Chem. 2008, 56, 9699–9709. [Google Scholar] [CrossRef]
- Arranz, S.; Calixto, F.S. Analysis of polyphenols in cereals may be improved performing acidic hydrolysis: A study in wheat flour and wheat bran and cereals of the diet. J. Cereal Sci. 2010, 51, 313–318. [Google Scholar] [CrossRef]
- Sani, I.M.; Iqbal, S.; Chan, K.W.; Ismail, M. Effect of acid and base catalyzed hydrolysis on the yield of phenolics and antioxidant activity of extracts from germinated brown rice (GBR). Molecules 2012, 17, 7584–7594. [Google Scholar] [CrossRef]
- Khakimov, B.; Motawia, M.S.; Bak, S.; Engelsen, S.B. The use of trimethylsilyl cyanide derivatization for robust and broad-spectrum high-throughput gas chromatography-mass spectrometry based metabolomics. Anal. Bioanal. Chem. 2013, 405, 9193–9205. [Google Scholar] [CrossRef]
- Bro, R.; Andersson, C.A.; Kiers, H.A.L. PARAFAC2—Part II. Modeling chromatographic data with retention time shifts. J. Chemom. 1999, 13, 295–309. [Google Scholar] [CrossRef]
- Amigo, J.M.; Skov, T.; Coello, J.; Maspoch, S.; Bro, R. Solving GC-MS problems with PARAFAC2. Trac-Trends Anal. Chem. 2008, 27, 714–725. [Google Scholar] [CrossRef]
- Khakimov, B.; Amigo, J.M.; Bak, S.; Engelsen, S.B. Plant metabolomics: Resolution and quantification of elusive peaks in liquid chromatography-mass spectrometry profiles of complex plant extracts using multi-way decomposition methods. J. Chromatogr. A 2012, 1266, 84–94. [Google Scholar] [CrossRef]
- Vandendool, H.; Kratz, P.D. A generalization of retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. 1963, 11, 463. [Google Scholar] [CrossRef]
- Golm Metabolome Database. Available online: http://gmd.mpimp-golm.mpg.de/ (accessed on 5 November 2013).
- Hotelling, H. Analysis of a complex of statistical variables into principal components. J. Educ. Psychol. 1933, 24, 417–441. [Google Scholar] [CrossRef]
- Sumner, L.; Amberg, A.; Barrett, D.; Beale, M.; Beger, R.; Daykin, C.; Fan, T.; Fiehn, O.; Goodacre, R.; Griffin, J.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khakimov, B.; Jespersen, B.M.; Engelsen, S.B. Comprehensive and Comparative Metabolomic Profiling of Wheat, Barley, Oat and Rye Using Gas Chromatography-Mass Spectrometry and Advanced Chemometrics. Foods 2014, 3, 569-585. https://doi.org/10.3390/foods3040569
Khakimov B, Jespersen BM, Engelsen SB. Comprehensive and Comparative Metabolomic Profiling of Wheat, Barley, Oat and Rye Using Gas Chromatography-Mass Spectrometry and Advanced Chemometrics. Foods. 2014; 3(4):569-585. https://doi.org/10.3390/foods3040569
Chicago/Turabian StyleKhakimov, Bekzod, Birthe Møller Jespersen, and Søren Balling Engelsen. 2014. "Comprehensive and Comparative Metabolomic Profiling of Wheat, Barley, Oat and Rye Using Gas Chromatography-Mass Spectrometry and Advanced Chemometrics" Foods 3, no. 4: 569-585. https://doi.org/10.3390/foods3040569
APA StyleKhakimov, B., Jespersen, B. M., & Engelsen, S. B. (2014). Comprehensive and Comparative Metabolomic Profiling of Wheat, Barley, Oat and Rye Using Gas Chromatography-Mass Spectrometry and Advanced Chemometrics. Foods, 3(4), 569-585. https://doi.org/10.3390/foods3040569