
Molecular Surveillance of Cronobacter spp. Isolated from a Wide Variety of Foods from 44 Different Countries by Sequence Typing of 16S rRNA, rpoB and O-Antigen Genes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Food Samples
2.2. Pre-Enrichment, Secondary Enrichment, and Culture on Chromogenic Media
2.3. Isolate Identification
2.4. DNA Extraction and Real-Time PCR
2.5. PCR Amplification
2.6. DNA Sequencing and Data Analysis
2.7. Nucleotide Sequence Accession Numbers
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Disclaimer
References
- Centers for Disease Control Prevention. Enterobacter sakazakii infections associated with the use of powdered infant formula—Tennessee, 2001. Morb. Mortal. Wkly. Rep. 2002, 51, 297–300. [Google Scholar]
- Lai, K.K. Enterobacter sakazakii Infections among Neonates, Infants, Children, and Adults. Medicine 2001, 80, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Osaili, T.; Forsythe, S. Desiccation resistance and persistence of Cronobacter species in infant formula. Int. J. Food Microbiol. 2009, 136, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Iversen, C.; Mullane, N.; McCardell, B.; Tall, B.D.; Lehner, A.; Fanning, S.; Stephan, R.; Joosten, H. Cronobacter gen. nov., a new genus to accommodate the biogroups of Enterobacter sakazakii, and proposal of Cronobacter sakazakii gen. nov., comb. nov., Cronobacter malonaticus sp. nov., Cronobacter turicensis sp. nov., Cronobacter muytjensii sp. nov., Cronobacter dublinensis sp. nov., Cronobacter genomospecies 1, and of three subspecies, Cronobacter dublinensis subsp. dublinensis subsp. nov., Cronobacter dublinensis subsp. lausannensis subsp. nov. and Cronobacter dublinensis subsp. lactaridi subsp. nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 1442–1447. [Google Scholar] [PubMed]
- Jarvis, K.G.; Yan, Q.Q.; Grim, C.J.; Power, K.A.; Franco, A.A.; Hu, L.; Gopinath, G.; Sathyamoorthy, V.; Kotewicz, M.L.; Kothary, M.H. Identification and Characterization of Five New Molecular Serogroups of Cronobacter spp. Foodborne Pathog. Dis. 2013, 10, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.; Cetinkaya, E.; Drahovska, H.; Levican, A.; Figueras, M.J.; Forsythe, S.J. Cronobacter condimenti sp. nov., isolated from spiced meat, and Cronobacter universalis sp. nov., a species designation for Cronobacter sp. genomospecies 1, recovered from a leg infection, water and food ingredients. Int. J. Syst. Evol. Microbiol. 2011, 62, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.; Sonbol, H.; Hariri, S.; Desai, P.; McClelland, M.; Forsythe, S.J. Diversity of the Cronobacter Genus as Revealed by Multilocus Sequence Typing. J. Clin. Microbiol. 2012, 50, 3031–3039. [Google Scholar] [CrossRef] [PubMed]
- Kandhai, M.C.; Reij, M.W.; Grognou, C.; van Schothorst, M.; Gorris, L.G.; Zwietering, M.H. Effects of preculturing conditions on lag time and specific growth rate of Enterobacter sakazakii in reconstituted powdered infant formula. Appl. Environ. Microbiol. 2006, 72, 2721–2729. [Google Scholar] [CrossRef] [PubMed]
- Strydom, A.; Cameron, M.; Witthuhn, R.C. Phylogenetic Analysis of Cronobacter Isolates Based on the rpoA and 16S rRNA Genes. Curr. Microbiol. 2012, 64, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Jason, J. The Roles of Epidemiologists, Laboratorians, and Public Health Agencies in Preventing Invasive Cronobacter Infection. Front. Pediatr. 2015, 3, 110. [Google Scholar] [CrossRef] [PubMed]
- Hochel, I.; Růžičková, H.; Krásný, L.; Demnerová, K. Occurrence of Cronobacter spp. in retail foods. J. Appl. Microbiol. 2012, 112, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Carter, L.; Lindsey, L.A.; Grim, C.J.; Sathyamoorthy, V.; Jarvis, K.G.; Gopinath, G.; Lee, C.; Sadowski, J.A.; Trach, L.; Pava-Ripoll, M. Multiplex PCR Assay Targeting a Diguanylate Cyclase-Encoding Gene, cgcA, to Differentiate Species within the Genus Cronobacter. Appl. Environ. Microbiol. 2012, 79, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.; Jacobs, E.; Simpson, S.; Kerdahi, K. Genetic Characterization of Fungi Isolated from the Environmental Swabs collected from a Compounding Center Known to Cause Multistate Meningitis Outbreak in United States Using ITS Sequencing. Pathogens 2014, 3, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.; Jacobs, E.; Simpson, S.; Kerdahi, K. Molecular Identification of Isolated Fungi from Unopened Containers of Greek Yogurt by DNA Sequencing of Internal Transcribed Spacer Region. Pathogens 2014, 3, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.M.; Ortega, Y.; Simpson, S.; Kerdahi, K. Genetic characterization of human-pathogenic Cyclospora cayetanensis parasites from three endemic regions at the 18S ribosomal RNA locus. Infect. Genet. Evol. 2014, 22, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Fanning, S.A. Strategies for the Identification and Tracking of Cronobacter Species: An Opportunistic Pathogen of Concern to Neonatal Health. Front. Pediatr. 2015, 3, 38. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.M.; Jacobs, E.; Segars, K.; Simpson, S.; Kerdahi, K. Genetic Characterization of Cronobacter sakazakii Recovered from the Environmental Surveillance Samples During a Sporadic Case Investigation of Foodborne Illness. Curr. Microbiol. 2016, 73, 273–279. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration Bacteriological Analytical Manual (8th Edition). Available online: https://www.fda.gov/food/foodscienceresearch/laboratorymethods/ucm114664.htm (accessed on 24 February 2017).
- Al-Holy, M.A.; Shin, J.H.; Osaili, T.M.; Rasco, B.A. Evaluation of a New Enrichment Broth for Detection of Cronobacter spp. in Powdered Infant Formula. J. Food Prot. 2011, 74, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Pincus, D.H. Microbial Identification Using the BioMerieux VITEK® 2 System. In Encyclopedia of Rapid Microbiological Methods; Parenteral Drug Association: Bethesda, MD, USA, 2006. [Google Scholar]
- Lehner, A.; Tasara, T.; Stephan, R. 16S rRNA gene based analysis of Enterobacter sakazakii strains from different sources and development of a PCR assay for identification. BMC Microbiol. 2004, 4, 43. [Google Scholar] [CrossRef] [PubMed]
- Mollet, C.; Drancourt, M.; Raoult, D. rpoB sequence analysis as a novel basis for bacterial identification. Mol. Microbiol. 1997, 26, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, M.; Wang, Q.; Cao, B.; He, X.; Li, K.; Feng, L.; Wang, L. Genetic Analysis of the Cronobacter sakazakii O4 to O7 O-Antigen Gene Clusters and Development of a PCR Assay for Identification of All C. sakazakii O Serotypes. Appl. Environ. Microbiol. 2012, 78, 3966–3974. [Google Scholar] [CrossRef] [PubMed]
- Bowen, A.B.; Braden, C.R. Invasive Enterobacter sakazakii Disease in Infants. Emerg. Infect. Dis. 2006, 12, 1185–1189. [Google Scholar] [CrossRef] [PubMed]
- Iversen, C.; Forsythe, S. Risk profile of Enterobacter sakazakii, an emergent pathogen associated with infant milk formula. Trends Food Sci.Technol. 2003, 14, 443–454. [Google Scholar] [CrossRef]
- Muytjens, H.L.; Roelofs-Willemse, H.; Jaspar, G.H. Quality of powdered substitutes for breast milk with regard to members of the family Enterobacteriaceae. J. Clin. Microbiol. 1988, 26, 743–746. [Google Scholar] [PubMed]
- Chen, Y.I.; Song, K.-Y.; Brown, E.W.; Lampel, K.A. Development of an Improved Protocol for the Isolation and Detection of Enterobacter sakazakii (Cronobacter) from Powdered Infant Formula. J. Food Prot. 2010, 73, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Segars, K.; Simpson, S.; Kerdahi, K.; Sulaiman, I.M. Evaluation of Cronobacter Growth and Phenotypic Variation Under Modified Culture Conditions. Curr. Microbiol. 2015, 72, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.M.; Fayer, R.; Lal, A.A.; Trout, J.M.; Schaefer, F.W.; Xiao, L. Molecular Characterization of Microsporidia Indicates that Wild Mammals Harbor Host-Adapted Enterocytozoon spp. as well as Human-Pathogenic Enterocytozoon bieneusi. Appl. Environ. Microbiol. 2003, 69, 4495–4501. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.M.; Lal, A.A.; Xiao, L. A Population Genetic Study of the Cryptosporidium parvum Human Genotype Parasites. J. Eukaryot. Microbiol. 2001, 48, 24S–27S. [Google Scholar] [CrossRef]
- Stoop, B.; Lehner, A.; Iversen, C.; Fanning, S.; Stephan, R. Development and evaluation of rpoB based PCR systems to differentiate the six proposed species within the genus Cronobacter. Int. J. Food Microbiol. 2009, 136, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, M.; Liu, H.; Wang, J.; He, X.; Zeng, J.; Guo, X.; Li, K.; Cao, B.; Wang, L. Development of an O-Antigen Serotyping Scheme for Cronobacter sakazakii. Appl. Environ. Microbiol. 2011, 77, 2209–2214. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.M.; Anderson, M.; Khristova, M.; Tang, K.; Sulaiman, N.; Phifer, E.; Simpson, S.; Kerdahi, K. Development of a PCR–Restriction Fragment Length Polymorphism Protocol for Rapid Detection and Differentiation of Four Cockroach Vectors (Group I “Dirty 22” Species) Responsible for Food Contamination and Spreading of Foodborne Pathogens: Public Health Importance. J. Food Prot. 2011, 74, 1883–1890. [Google Scholar] [PubMed]
- Sulaiman, I.M.; Anderson, M.; Oi, D.H.; Simpson, S.; Kerdahi, K. Multilocus Genetic Characterization of Two Ant Vectors (Group II “Dirty 22” Species) Known To Contaminate Food and Food Products and Spread Foodborne Pathogens. J. Food Prot. 2012, 75, 1447–1452. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.M.; Torres, P.; Simpson, S.; Kerdahi, K.; Ortega, Y. Sequence Characterization of Heat Shock Protein Gene of Cyclospora cayetanensis Isolates from Nepal, Mexico, and Peru. J. Parasitol. 2013, 99, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, A.; Loughlin, M.; Caubilla-Barron, J.; Kucerova, E.; Manning, G.; Dowson, C.; Forsythe, S. Multilocus sequence typing of Cronobacter sakazakii and Cronobacter malonaticus reveals stable clonal structures with clinical significance which do not correlate with biotypes. BMC Microbiol. 2009, 9, 223. [Google Scholar] [CrossRef] [PubMed]
- Cetinkaya, E.; Joseph, S.; Ayhan, K.; Forsythe, S.J. Comparison of methods for the microbiological identification and profiling of Cronobacter species from ingredients used in the preparation of infant formula. Mol. Cell. Probes 2013, 27, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Gičová, A.; Oriešková, M.; Oslanecová, L.; Drahovská, H.; Kaclíková, E. Identification and characterization of Cronobacter strains isolated from powdered infant foods. Lett. Appl. Microbiol. 2013, 58, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Killer, J.; Skřivanová, E.; Hochel, I.; Marounek, M. Multilocus Sequence Typing of Cronobacter Strains Isolated from Retail Foods and Environmental Samples. Foodborne Pathog. Dis. 2015, 12, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Stephan, R.; Fricker-Feer, C.; Lehner, A. Genetic Diversity of Cronobacter sakazakii Isolates Collected from a Swiss Infant Formula Production Facility. J. Food Prot. 2013, 76, 883–887. [Google Scholar] [CrossRef] [PubMed]
- Sonbol, H.; Joseph, S.; McAuley, C.M.; Craven, H.M.; Forsythe, S.J. Multilocus sequence typing of Cronobacter spp. from powdered infant formula and milk powder production factories. Int. Dairy J. 2013, 30, 1–7. [Google Scholar] [CrossRef]
- Akineden, Ö.; Murata, K.J.; Gross, M.; Usleber, E. Microbiological Quality of Raw Dried Pasta from the German Market, with Special Emphasis on Cronobacter Species. J. Food Sci. 2015, 80, M2860–M2867. [Google Scholar] [CrossRef] [PubMed]
- Lou, X.; Si, G.; Yu, H.; Qi, J.; Liu, T.; Fang, Z. Possible reservoir and routes of transmission of Cronobacter (Enterobacter sakazakii) via wheat flour. Food Control 2014, 43, 258–262. [Google Scholar] [CrossRef]
- Huang, Y.; Pang, Y.; Wang, H.; Tang, Z.; Zhou, Y.; Zhang, W.; Li, X.; Tan, D.; Li, J.; Lin, Y. Occurrence and Characterization of Cronobacter spp. in Dehydrated Rice Powder from Chinese Supermarket. PLoS ONE 2015, 10, e0131053. [Google Scholar] [CrossRef] [PubMed]
- Ogihara, H.; Kiribe, N.; Fukuda, N.; Furukawa, S.; Morinaga, Y.; Igimi, S. Cronobacter spp. in Commercially Available Dried Food in Japan. Biocontrol Sci. 2014, 19, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Brandão, M.L.L.; Umeda, N.S.; Jackson, E.; Forsythe, S.J.; de Filippis, I. Isolation, molecular and phenotypic characterization, and antibiotic susceptibility of Cronobacter spp. from Brazilian retail foods. Food Microbiol. 2017, 63, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, A.; Grand, M.; Liniger, M.; Iversen, C. Detection and frequency of Cronobacter spp. (Enterobacter sakazakii) in different categories of ready-to-eat foods other than infant formula. Int. J. Food Microbiol. 2009, 136, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, C.; Wu, Q.; Zhang, J.; Huang, J.; Yang, G. Prevalence, molecular characterization, and antibiotic susceptibility of Cronobacter spp. in Chinese ready-to-eat foods. Int. J. Food Microbiol. 2015, 204, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Jaradat, Z.W.; Ababneh, Q.O.; Saadoun, I.M.; Samara, N.A.; Rashdan, A.M. Isolation of Cronobacter spp. (formerly Enterobacter sakazakii) from infant food, herbs and environmental samples and the subsequent identification and confirmation of the isolates using biochemical, chromogenic assays, PCR and 16S rRNA sequencing. BMC Microbiol. 2009, 9, 225. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Goel, G.; Raghav, M. Prevalence and Characterization of Cronobacter spp. from Various Foods, Medicinal Plants, and Environmental Samples. Curr. Microbiol. 2015, 71, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Garbowska, M.; Berthold-Pluta, A.; Stasiak-Różańska, L. Microbiological quality of selected spices and herbs including the presence of Cronobacter spp. Food Microbiol. 2015, 49, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Fei, P.; Man, C.; Lou, B.; Forsythe, S.J.; Chai, Y.; Li, R.; Niu, J.; Jiang, Y. Genotyping and Source Tracking of Cronobacter sakazakii and C. malonaticus Isolates from Powdered Infant Formula and an Infant Formula Production Factory in China. Appl. Environ. Microbiol. 2015, 81, 5430–5439. [Google Scholar] [CrossRef] [PubMed]
- Jackson, E.E.; Flores, J.P.; Fernández-Escartín, E.; Forsythe, S.J. Reevaluation of a Suspected Cronobacter sakazakii Outbreak in Mexico. J. Food Prot. 2015, 78, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
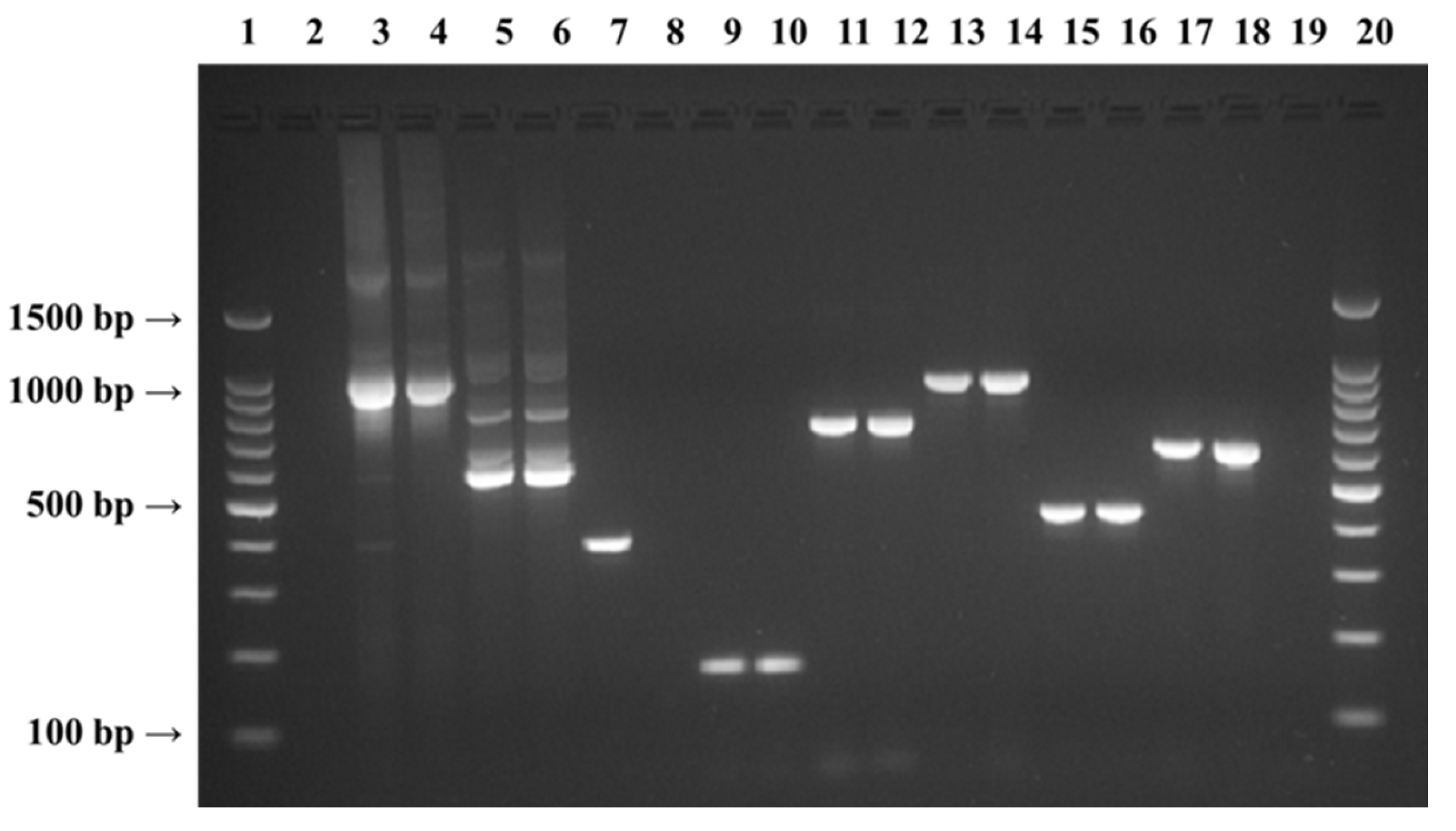

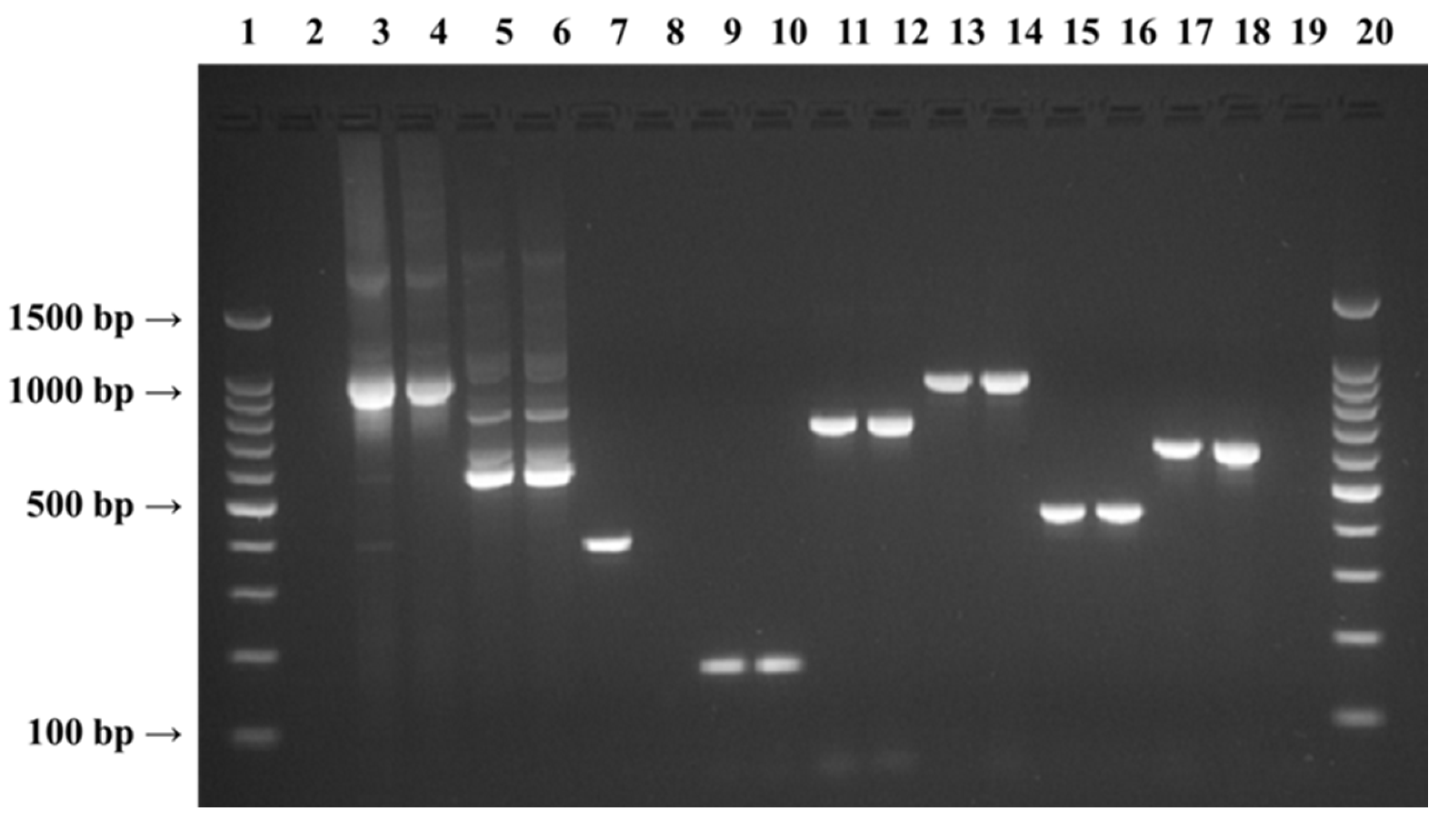{kind=link}
| Origin (Region/Country) | Food Products Tested |
|---|---|
| Americas | |
| Argentina | chia seed, pet food |
| Belize | pet food, papaya |
| Brazil | papaya |
| Canada | pet food, sesame seed |
| Chile | chili powder |
| Colombia | basil |
| Costa Rica | papaya, coriander |
| Dominican Republic | cantaloupe, cilantro, cucumber, papaya |
| Ecuador | ready-to-eat snack |
| El Salvador | okra, spice powder |
| Guatemala | breading flour, mango, papaya, seasoned flour |
| Guyana | brown sauce |
| Haiti | mango |
| Honduras | cucumber |
| Jamaica | spice powder |
| Mexico | avocado, basil, cilantro, kale, octopus, yellow croaker |
| Nicaragua | cheese |
| Peru | paprika powder, shrimp |
| USA | alfalfa sprout, avocado, broccoli sprout, broccoli sprout seed, cheese, clover seed, cucumber, frozen ravioli, kale, organic clover sprout, parsley, pet food, powder infant formula, powder milk, ready-to-eat snack, spice powder, spinach, tomato |
| Africa | |
| Sao Vicente (Cape Verde) | ready-to-eat snack |
| Ghana | ogbono seed, smoked tilapia, spice |
| Kenya | spice powder |
| Morocco | ready-to-eat snack |
| South Africa | pepper, spice powder |
| Asia | |
| China | cauliflower, frozen crab cake, garlic powder, pet food, spice powder, tilapia |
| India | black pepper, crushed red pepper, garlic powder, sesame seed, spice powder, ready-to-eat snack, wafers wheels |
| Indonesia | spice powder, tilapia |
| Israel | basil |
| Malaysia | shrimp |
| Pakistan | ready-to-eat snack, spice powder |
| Philippines | cassava leaf, desiccated coconut |
| Sri Lanka | cinnamon, cinnamon quill |
| South Korea | ready-to-eat snack |
| Taiwan | black sesame powder, spice salt, spice powder |
| Thailand | ready-to-eat snack, spice powder |
| Turkey | laurel leaves, strawberry |
| Vietnam | ground black pepper, ready-to-eat snack, shrimp, tuna, white pepper |
| Europe | |
| Germany | chocolate powder, spice powder |
| Ireland | pet food, ready-to-eat snack |
| Italy | frozen linguine, ready-to-eat snack |
| Netherland | parsley leaf |
| Spain | cantaloupe, crawfish, paprika |
| UK | medical food, salmon |
| Oceania | |
| Australia | alfalfa beans |
| S. No. | Sample Number | Description of Food Products | Country of Origin | Food Product Type/Category | Bacterial Culture | QPCR | PCR Screening | * O-antigen Reference (Sequence Variation, % Similarity) | * O-antigen Sequence Type | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 16S rRNA | rpoB | * O-antigen | |||||||||
| 1 | SRL-66 | Cassava leaf | Philippines | Dried food | TG | + | + | + | 6 | JQ674749, This report (Identical) | C. sakazakii |
| 2 | SRL-80 | Breading flour | Guatemala | Dried food | TG | + | + | + | 1 | CP011047, This report (Identical) | C. sakazakii |
| 3 | SRL-154 | Frozen Ravioli | USA | Frozen food | TG | + | + | + | 6 | JQ674749, This report (4-point-mutation) | C. sakazakii |
| 4 | SRL-86 | Pet food | USA | Pet food | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 5 | SRL-91 | Pet food | Canada | Pet food | TG | + | + | + | NOA | NOA | NOA |
| 6 | SRL-93 | Pet food | Canada | Pet food | TG | + | + | + | 1 | CP000783, This report (Identical) | C. sakazakii |
| 7 | SRL-101 | Pet food | Canada | Pet food | TG | + | + | + | 1 | CP000783, This report (Identical) | C. sakazakii |
| 8 | SRL-163 | Pet food | China | Pet food | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 9 | SRL-186 | Pet food | USA | Pet food | TG | + | + | + | NOA | NOA | NOA |
| 10 | SRL-199 | Pet food | China | Pet food | TG | + | + | + | 1 | CP011047, This report (Identical) | C. sakazakii |
| 11 | SRL-12 | Basil | Mexico | Produce | TG | + | + | + | NOA | NOA | NOA |
| 12 | SRL-13 | Parsley | USA | Produce | TG | + | + | + | NOA | NOA | NOA |
| 13 | SRL-20 | Basil | Colombia | Produce | TG | + | + | + | 1 | CP000783, This report (1-point-mutation) | C. sakazakii |
| 14 | SRL-35 | Alfalfa beans | Australia | Produce | TG | + | + | + | 1 | CP011047, This report (Identical) | C. sakazakii |
| 15 | SRL-42 | Basil | Colombia | Produce | TG | + | + | + | NOA | NOA | NOA |
| 16 | SRL-87 | Parsley leaf | Netherland | Produce | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 17 | SRL-94 | Alfalfa sprout | USA | Produce | TG | + | + | + | NOA | NOA | NOA |
| 18 | SRL-140 | Avocado | USA | Produce | TG | + | + | + | 6 | JQ674749, This report (Identical) | C. sakazakii |
| 19 | SRL-173 | Avocado | USA | Produce | TG | + | + | + | 3 | HQ646169, This report (Identical) | C. sakazakii |
| 20 | SRL-194 | Avocado | USA | Produce | TG | + | + | + | 6 | JQ674749, This report (4-point-mutation) | C. sakazakii |
| 21 | SRL-95 | Smoked Tilapia | Ghana | Seafood | TG | + | + | + | 7 | JQ674750, This report (2-point-mutation) | C. sakazakii |
| 22 | SRL-4 | Garlic powder | India | Spice | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 23 | SRL-36 | Spice powder | Pakistan | Spice | TG | + | + | + | 3 | HQ646169, This report (Identical) | C. sakazakii |
| 24 | SRL-40 | Spice powder | South Africa | Spice | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 25 | SRL-41 | Pepper | South Africa | Spice | TG | + | + | + | 1 | CP011047, This report (Identical) | C. sakazakii |
| 26 | SRL-43 | Spice salt | Taiwan | Spice | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 27 | SRL-47 | Black sesame powder | Taiwan | Spice | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 28 | SRL-56 | Spice powder | India | Spice | TG | + | + | + | 1 | CP011047, This report (Identical) | C. sakazakii |
| 29 | SRL-65 | Spice powder | India | Spice | TG | + | + | + | 4 | JQ674747, This report (Identical) | C. sakazakii |
| 30 | SRL-77 | Spice powder | India | Spice | TG | + | + | + | 1 | CP011047, This report (Identical) | C. sakazakii |
| 31 | SRL-81 | Spice powder | Pakistan | Spice | TG | + | + | + | 7 | JQ674750, This report (2-point-mutation) | C. sakazakii |
| 32 | SRL-82 | Spice powder | India | Spice | TG | + | + | + | 3 | HQ646169, This report (Identical) | C. sakazakii |
| 33 | SRL-102 | Ogbono seed | Ghana | Spice | TG | + | + | + | NOA | NOA | NOA |
| 34 | SRL-104 | Spice powder | Jamaica | Spice | TG | + | + | + | 1 | CP011047, This report (Identical) | C. sakazakii |
| 35 | SRL-109 | Paprika powder | Peru | Spice | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 36 | SRL-124 | Spice powder | Germany | Spice | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 37 | SRL-126 | Chili powder | Chile | Spice | TG | + | + | + | NOA | NOA | NOA |
| 38 | SRL-128 | Spice powder | Kenya | Spice | TG | + | + | + | 6 | JQ674749, This report (identical) | C. sakazakii |
| 39 | SRL-160 | Spice powder | El Salvador | Spice | TG | + | + | + | 3 | HQ646169, This report (Identical) | C. sakazakii |
| 40 | SRL-171 | Spice powder | India | Spice | TG | + | + | + | 1 | CP011047, This report (Identical) | C. sakazakii |
| 41 | SRL-172 | Spice powder | India | Spice | TG | + | + | + | 6 | JQ674749, This report (Identical) | C. sakazakii |
| 42 | SRL-179 | Spice powder | India | Spice | TG | + | + | + | NOA | NOA | NOA |
| 43 | SRL-180 | Spice powder | India | Spice | TG | + | + | + | 6 | JQ674749, This report (Identical) | C. sakazakii |
| 44 | SRL-181 | Spice powder | India | Spice | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 45 | SRL-187 | Spice powder | India | Spice | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 46 | SRL-51 | Ready-to-eat snack | India | Snack | TG | + | + | + | 4 | JQ674747, This report (Identical) | C. sakazakii |
| 47 | SRL-72 | Ready-to-eat snack | Vietnam | Snack | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 48 | SRL-79 | Ready-to-eat snack | India | Snack | TG | + | + | + | 2 | EU076546, This report (Identical) | C. sakazakii |
| 49 | SRL-99 | Ready-to-eat snack | USA | Snack | TG | + | + | + | 1 | CP000783, This report (Identical) | C. sakazakii |
| 50 | SRL-131 | Ready-to-eat snack | India | Snack | TG | + | + | + | 6 | JQ674749, This report (Identical) | C. sakazakii |
| 51 | SRL-152 | Ready-to-eat snack | India | Snack | TG | + | + | + | 1 | CP011047, This report (Identical) | C. sakazakii |
| Target | Primer Name | Primer Sequence (5′–3′) | Reference |
|---|---|---|---|
| * 16S rRNA | 616V | AGAGTTGATYMTGGCTC | [21] |
| 630R | CAKAAAGGAGGTGATCC | ||
| * rpoB | rpoB-F | AACCAGTTCCGCGTTGGCCTGG | [22] |
| rpoB-R | CCTGAACAACACGCTCGGA | ||
| ** Wzy, Serotype O1 | wl-35646 | CCCGCTTGTATGGATGTT | [23] |
| wl-35647 | CTTTGGGAGCGTTAGGTT | ||
| ** Wzy, Serotype O2 | wl-37256 | ATTGTTTGCGATGGTGAG | [23] |
| wl-37257 | AAAACAATCCAGCAGCAA | ||
| ** Wzy, Serotype O3 | wl-37258 | CTCTGTTACTCTCCATAGTGTTC | [23] |
| wl-37259 | GATTAGACCACCATAGCCA | ||
| ** Wzy, Serotype O4 | wl-39105 | ACTATGGTTTGGCTATACTCCT | [23] |
| wl-39106 | ATTCATATCCTGCGTGGC | ||
| ** Wzy, Serotype O5 | wl-39873 | GATGATTTTGTAAGCGGTCT | [23] |
| wl-39874 | ACCTACTGGCATAGAGGATAA | ||
| ** Wzy, Serotype O6 | wl-40041 | ATGGTGAAGGGAACGACT | [23] |
| wl-40042 | ATCCCCGTGCTATGAGAC | ||
| ** Wzy, Serotype O7 | wl-40039 | CATTTCCAGATTATTACCTTTC | [23] |
| wl-40040 | ACACTGGCGATTCTACCC |
| Sample Number | Description of Food Products | Country of Origin | 16S rRNA Reference * (Sequence Variation, % Similarity) | 16S rRNA Sequence Type | rpoB Reference ** (Sequence Variation, % Similarity) | rpoB Sequence Type |
|---|---|---|---|---|---|---|
| SRL-91 | Pet food | Canada | GU122174, This report (Identical, 100%) | C. malonaticus | CP013940, This report (8-point-mutation, 99%) | C. malonaticus |
| NR_102802, This report (Identical, 100%) | C. turicensis | |||||
| SRL-186 | Pet food | USA | KF360293, This report (Identical, 100%) | C. malonaticus | CP013940, This report (Identical, 100%) | C. malonaticus |
| KU364482, This report (Identical, 100%) | C. sakazakii | |||||
| SRL-12 | Produce | Mexico | CP004091, This report (Identical, 100%) | C. sakazakii | CP013940, This report (3-point-mutation, 99%) | C. malonaticus |
| JF330141, This report (3-point-mutation, 99%) | C. sakazakii | |||||
| SRL-13 | Produce | USA | CP012266, This report (Identical, 100%) | C. dublinensis | AB980795, This report (9-point-mutation, 98%) | C. dublinensis |
| KU364468, This report (Identical, 100%) | C. sakazakii | |||||
| SRL-42 | Produce | Colombia | KC818225, This report (2-point-mutation, 99%) | C. malonaticus | CP013940, This report (Identical, 100%) | C. malonaticus |
| KU543632, This report (2-point-mutation, 99%) | C. sakazakii | |||||
| SRL-94 | Produce | USA | KC109002, This report (2-point-mutation, 99%) | C. malonaticus | CP013940, This report (Identical, 100%) | C. malonaticus |
| CP004091, This report (2-point-mutation, 99%) | C. sakazakii | |||||
| HQ880409, This report (2-point-mutation, 99%) | C. turicensis | |||||
| SRL-102 | Spice | Ghana | KC109002, This report (2-point-mutation, 99%) | C. malonaticus | JX425275, This report (3-point-mutation, 99%) | C. sakazakii |
| CP004091, This report (2-point-mutation, 99%) | C. sakazakii | |||||
| HQ880409, This report (2-point-mutation, 99%) | C. turicensis | |||||
| SRL-126 | Spice | Chile | CP012266, This report (Identical, 100%) | C. dublinensis | JX425283, This report (6-point-mutation, 99%) | C. dublinensis |
| KU364468, This report (Identical, 100%) | C. sakazakii | |||||
| SRL-179 | Spice | India | KU364464, This report (Identical, 100%) | C. sakazakii | JF330150, This report (Identical, 100%) | C. sakazakii |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda, N.; Banerjee, P.; Simpson, S.; Kerdahi, K.; Sulaiman, I.M. Molecular Surveillance of Cronobacter spp. Isolated from a Wide Variety of Foods from 44 Different Countries by Sequence Typing of 16S rRNA, rpoB and O-Antigen Genes. Foods 2017, 6, 36. https://doi.org/10.3390/foods6050036
Miranda N, Banerjee P, Simpson S, Kerdahi K, Sulaiman IM. Molecular Surveillance of Cronobacter spp. Isolated from a Wide Variety of Foods from 44 Different Countries by Sequence Typing of 16S rRNA, rpoB and O-Antigen Genes. Foods. 2017; 6(5):36. https://doi.org/10.3390/foods6050036
Chicago/Turabian StyleMiranda, Nancy, Pratik Banerjee, Steven Simpson, Khalil Kerdahi, and Irshad M. Sulaiman. 2017. "Molecular Surveillance of Cronobacter spp. Isolated from a Wide Variety of Foods from 44 Different Countries by Sequence Typing of 16S rRNA, rpoB and O-Antigen Genes" Foods 6, no. 5: 36. https://doi.org/10.3390/foods6050036