Multistep Optimization of ?-Glucosidase Extraction from Germinated Soybeans (Glycine max L. Merril) and Recovery of Isoflavone Aglycones


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Soybean Germination Process
2.3. CCD-Based Optimization of Extraction of Active β-Glucosidase
2.4. CCD-Based Optimization of β-Glucosidase-Assisted Conversion of Conjugated Isoflavones into Their Corresponding Aglycones
2.5. Determination of β-Glucosidase Activity
2.6. Extraction and Determination of Isoflavones by Ultra-High-Performance Liquid Chromatography (UHPLC)
2.7. Statistical Analysis
3. Results and Discussion
3.1. Optimization of β-Glucosidase Extraction from Germinated Soybean Epicotyls
3.2. Optimization of the Recovery of Aglycones Using β-Glucosidase from Germinated Soybean Epicotyls
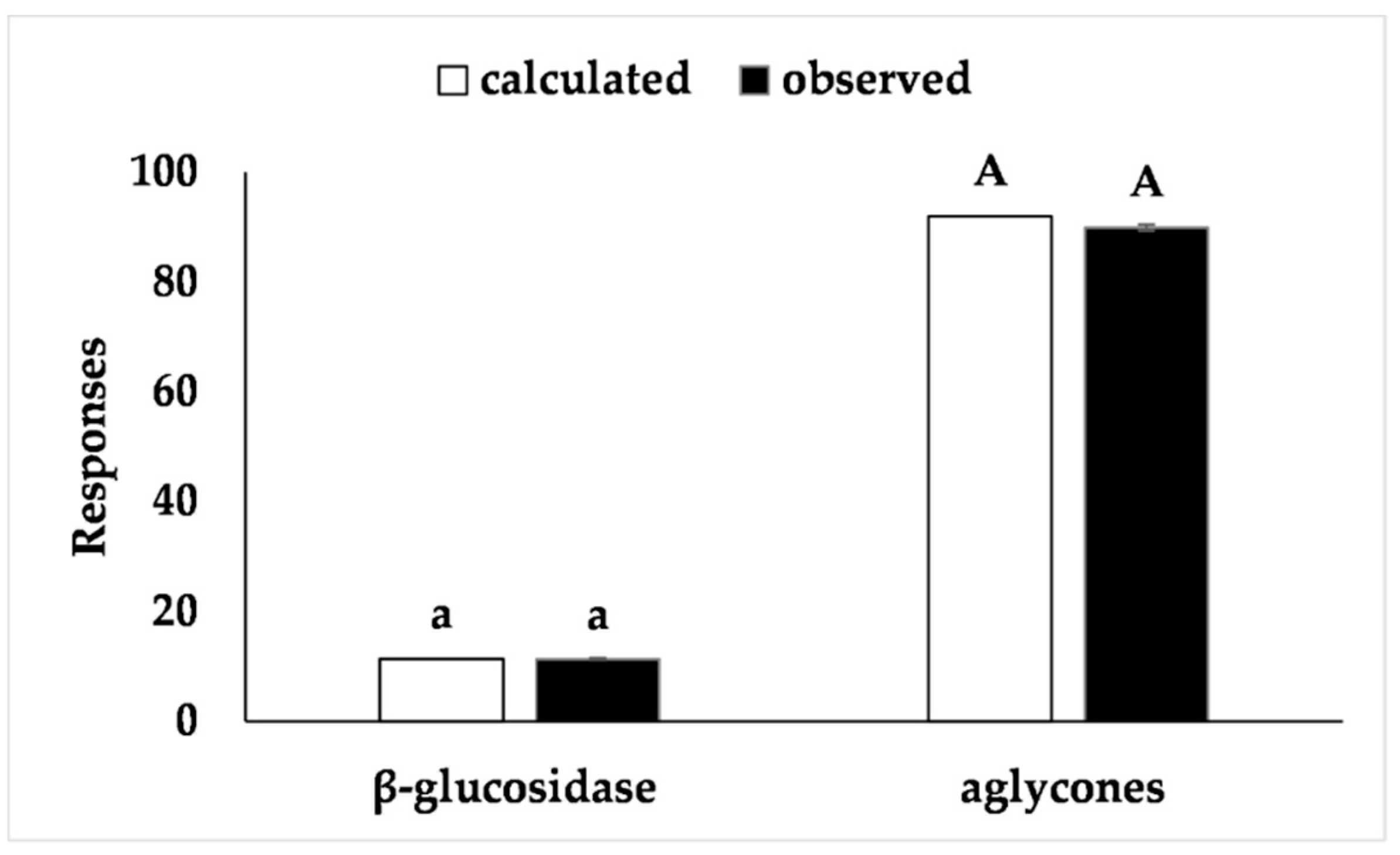
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hsieh, M.-C.; Graham, T.L. Partial purification and characterization of a soybean β-glucosidase with high specific activity towards isoflavone conjugates. Phytochemistry 2001, 58, 995–1005. [Google Scholar] [CrossRef]
- Villares, A.; Rostagno, M.A.; García-Lafuente, A.; Guillamón, E.; Martínez, J.A. Content and profile of isoflavones in soy-based foods as a function of the production process. Food Bioprocess Technol. 2011, 4, 27–38. [Google Scholar] [CrossRef]
- He, F.-J.; Chen, J.-Q. Consumption of soybean, soy foods, soy isoflavones and breast cancer incidence: Differences between Chinese women and women in western countries and possible mechanisms. Food Sci. Hum. Wellness 2013, 2, 146–161. [Google Scholar] [CrossRef]
- Brandi, M.L. Natural and synthetic isoflavones in the prevention and treatment of chronic diseases. Calcif. Tissue Int. 1997, 61, S5–S8. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, H.; Mazur, W. Phyto-oestrogens and western diseases. Ann. Med. 1997, 29, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Falcão, H.G.; Handa, C.L.; Silva, M.B.R.; de Camargo, A.C.; Shahidi, F.; Kurozawa, L.E.; Ida, E.I. Soybean ultrasound pre-treatment prior to soaking affects β-glucosidase activity, isoflavone profile and soaking time. Food Chem. 2018. [Google Scholar] [CrossRef]
- Kudou, S.; Fleury, Y.; Welti, D.; Magnolato, D.; Uchida, T.; Kitamura, K.; Okubo, K. Malonyl isoflavone glycosides in soybean seeds (Glycine max Merrill). Agric. Biol. Chem. 1991, 55, 2227–2233. [Google Scholar] [CrossRef]
- Wang, H.J.; Murphy, P.A. Isoflavone content in commercial soybean foods. J. Agric. Food Chem. 1994, 42, 1666–1673. [Google Scholar] [CrossRef]
- Tsukamoto, C.; Shimada, S.; Igita, K.; Kudou, S.; Kokubun, M.; Okubo, K.; Kitamura, K. Factors affecting isoflavone content in soybean seeds: Changes in isoflavones, saponins, and composition of fatty acids at different temperatures during seed development. J. Agric. Food Chem. 1995, 43, 1184–1192. [Google Scholar] [CrossRef]
- Walsh, K.R.; Zhang, Y.C.; Vodovotz, Y.; Schwartz, S.J.; Failla, M.L. Stability and bioaccessibility of isoflavones from soy bread during in vitro digestion. J. Agric. Food Chem. 2003, 51, 4603–4609. [Google Scholar] [CrossRef] [PubMed]
- Fukutake, M.; Takahashi, M.; Ishida, K.; Kawamura, H.; Sugimura, T.; Wakabayashi, K. Quantification of genistein and genistin in soybeans and soybean products. Food Chem. Toxicol. 1996, 34, 457–461. [Google Scholar] [CrossRef]
- Izumi, T.; Piskula, M.K.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kubota, Y.; Kikuchi, M. Soy isoflavone aglycones are absorbed faster and in higher amounts than their glucosides in humans. J. Nutr. 2000, 130, 1695–1699. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, M.; Obata, A.; Fukushima, D. Objectionable flavor of soy milk developed during the soaking of soybeans and its control. J. Food Sci. 1989, 54, 602–605. [Google Scholar] [CrossRef]
- Esen, A. Purification and partial characterization of maize (Zea mays L.) beta-glucosidase. Plant Physiol. 1992, 98, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.C.; Cheng, W.Y.; Wu, R.Y.; Huang, C.J.; Lee, K.T. Hydrolysis of black soybean isoflavone glycosides by Bacillus subtilis natto. Appl. Microbiol. Biotechnol. 2006, 73, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Chuankhayan, P.; Rimlumduan, T.; Svasti, J.; Cairns, J.R.K. Hydrolysis of soybean isoflavonoid glycosides by Dalbergia β-glucosidases. J. Agric. Food Chem. 2007, 55, 2407–2412. [Google Scholar] [CrossRef] [PubMed]
- Handa, C.L.; Couto, U.R.; Vicensoti, A.H.; Georgetti, S.R.; Ida, E.I. Optimisation of soy flour fermentation parameters to produce β-glucosidase for bioconversion into aglycones. Food Chem. 2014, 152, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Morant, A.V.; Jørgensen, K.; Jørgensen, C.; Paquette, S.M.; Sánchez-Pérez, R.; Møller, B.L.; Bak, S. β-Glucosidases as detonators of plant chemical defense. Phytochemistry 2008, 69, 1795–1813. [Google Scholar] [CrossRef] [PubMed]
- Santosh, T.R.; Balasubramanian, K.K.; Lalitha, K. Enhancement of β-Glucosidase and β-Galactosidase of Trigonella foenum-graecum by exposure to the allelochemical mimosine. J. Agric. Food Chem. 1999, 47, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.L.L.; Mandarino, J.M.G.; Carrão-Panizzi, M.C.; Oliveira, M.C.N.; Campo, C.B.H.; Nepomuceno, A.L.; Ida, E.I. β-Glucosidase activity and isoflavone content in germinated soybean radicles and cotyledons. J. Food Biochem. 2006, 30, 453–465. [Google Scholar] [CrossRef]
- Yoshiara, L.Y.; Madeira, T.B.; Ribeiro, M.L.L.; Mandarino, J.M.G.; Carrão-Panizzi, M.C.; Ida, E.I. β-Glucosidase activity of soybean (Glycine max) embryonic axis germinated in the presence or absence of light. J. Food Biochem. 2011, 36, 699–705. [Google Scholar] [CrossRef]
- Ferreres, F.; Grosso, C.; Gil-Izquierdo, A.; Valentão, P.; Mota, A.T.; Andrade, P.B. Optimization of the recovery of high-value compounds from pitaya fruit by-products using microwave-assisted extraction. Food Chem. 2017, 230, 463–474. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Zhang, L.-L.; Yue, X.-Y.; Liang, J.; Jiang, J.; Gao, X.-L.; Yue, P.-X. Optimization of ultrasound-assisted extraction of phenolic compounds and anthocyanins from blueberry (Vaccinium ashei) wine pomace. Food Chem. 2016, 204, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Liyana-Pathirana, C.; Shahidi, F. Optimization of extraction of phenolic compounds from wheat using response surface methodology. Food Chem. 2005, 93, 47–56. [Google Scholar] [CrossRef]
- Carrǎo-Panizzi, M.C.; Bordingnon, J.R. Activity of beta-glucosidase and levels of isoflavone glucosides in soybean cultivars affected by the environment. Pesq. Agropec. Bras. 2000, 35, 873–878. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, M.; Sasaki, J.; Murao, S. Studies on β-glucosidases from soybeans that hydrolyze daidzin and genistin: Isolation and characterization of an isozyme. Biosci. Biotechnol. Biochem. 1995, 59, 1623–1627. [Google Scholar] [CrossRef]
- Yoshiara, L.Y.; Madeira, T.B.; Delaroza, F.; Da Silva, J.B.; Ida, E.I. Optimization of soy isoflavone extraction with different solvents using the simplex-centroid mixture design. Int. J. Food Sci. Nutr. 2012, 63, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Coward, L.; Smith, M.; Kirk, M.; Barnes, S. Chemical modification of isoflavones in soyfoods during cooking and processing. Am. J. Clin. Nutr. 1998, 68, 1486S–1491S. [Google Scholar] [CrossRef] [PubMed]
- Hösel, W.; Todenhagen, R. Characterization of a β-glucosidase from Glycine Max which hydrolyses coniferin and syringin. Phytochemistry 1980, 19, 1349–1353. [Google Scholar] [CrossRef]
- Matsuura, M.; Obata, A. β-Glucosidases from soybeans hydrolyze daidzin and genistin. J. Food Sci. 1993, 58, 144–147. [Google Scholar] [CrossRef]
- Marazza, J.A.; Garro, M.S.; Savoy de Giori, G. Aglycone production by Lactobacillus rhamnosus CRL981 during soymilk fermentation. Food Microbiol. 2009, 26, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Song, X.; Yu, J. Overexpression of β-glucosidase from Thermotoga maritima for the production of highly purified aglycone isoflavones from soy flour. World J. Microbiol. Biotechnol. 2009, 25, 2165–2172. [Google Scholar] [CrossRef]
- John, J.A.; Shahidi, F. Phenolic compounds and antioxidant activity of Brazil nut (Bertholletia excelsa). J. Funct. Foods 2010, 2, 196–209. [Google Scholar] [CrossRef]
- Peng, H.; Li, W.; Li, H.; Deng, Z.; Zhang, B. Extractable and non-extractable bound phenolic compositions and their antioxidant properties in seed coat and cotyledon of black soybean (Glycinemax (L.) Merr). J. Funct. Foods 2017, 32, 296–312. [Google Scholar] [CrossRef]
- Arruda, H.S.; Pereira, G.A.; de Morais, D.R.; Eberlin, M.N.; Pastore, G.M. Determination of free, esterified, glycosylated and insoluble-bound phenolics composition in the edible part of araticum fruit (Annona crassiflora mart.) and its by-products by HPLC-ESI-MS/MSms. Food Chem. 2018, 245, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Ambigaipalan, P.; de Camargo, A.C.; Shahidi, F. Identification of phenolic antioxidants and bioactives of pomegranate seeds following juice extraction using HPLC-DAD-ESI-MSn. Food Chem. 2017, 221, 1883–1894. [Google Scholar] [CrossRef] [PubMed]
- De Camargo, A.C.; Regitano-d'Arce, M.A.B.; Rasera, G.B.; Canniatti-Brazaca, S.G.; do Prado-Silva, L.; Alvarenga, V.O.; Sant'Ana, A.S.; Shahidi, F. Phenolic acids and flavonoids of peanut by-products: Antioxidant capacity and antimicrobial effects. Food Chem. 2017, 237, 538–544. [Google Scholar] [CrossRef] [PubMed]
- De Camargo, A.C.; Regitano-d'Arce, M.A.B.; Shahidi, F. Phenolic profile of peanut by-products: antioxidant potential and inhibition of alpha-glucosidase and lipase activities. J. Am. Oil Chem. Soc. 2017, 94, 959–971. [Google Scholar] [CrossRef]
- Tremocoldi, M.A.; Rosalen, P.L.; Franchin, M.; Massarioli, A.P.; Denny, C.; Daiuto, É.R.; Paschoal, J.A.R.; Melo, P.S.; De Alencar, S.M. Exploration of avocado by-products as natural sources of bioactive compounds. PLoS ONE 2018, 13, e0192577. [Google Scholar] [CrossRef] [PubMed]
- Perera, N.; Ambigaipalan, P.; Shahidi, F. Epigallocatechin gallate (EGCG) esters with different chain lengths fatty acids and their antioxidant activity in food and biological systems. J. Food Bioact. 2018, 1, 124–133. [Google Scholar]
- Oldoni, T.L.C.; Melo, P.S.; Massarioli, A.P.; Moreno, I.A.M.; Bezerra, R.M.N.; Rosalen, P.L.; da Silva, G.V.J.; Nascimento, A.M.; Alencar, S.M. Bioassay-guided isolation of proanthocyanidins with antioxidant activity from peanut (Arachis hypogaea) skin by combination of chromatography techniques. Food Chem. 2016, 192, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.Y.; Shahidi, F. Antioxidant activity of resveratrol ester derivatives in food and biological model systems. Food Chem. 2018, 261, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Amarowicz, R.; Pegg, R.B. The potential protective effects of phenolic compounds against low-density lipoprotein oxidation. Curr. Pharm. Des. 2017, 23, 2754–2766. [Google Scholar] [CrossRef] [PubMed]
- De Camargo, A.C.; Regitano-d’Arce, M.A.B.; Gallo, C.R.; Shahidi, F. Gamma-irradiation induced changes in microbiological status, phenolic profile and antioxidant activity of peanut skin. J. Funct. Foods 2015, 12, 129–143. [Google Scholar] [CrossRef]
- Rahman, M.J.; de Camargo, A.C.; Shahidi, F. Phenolic and polyphenolic profiles of chia seeds and their in vitro biological activities. J. Funct. Foods 2017, 35, 622–634. [Google Scholar] [CrossRef]
- Sun, S.; Kadouh, H.C.; Zhu, W.; Zhou, K. Bioactivity-guided isolation and purification of α-glucosidase inhibitor, 6-O-D-glycosides, from tinta cão grape pomace. J. Funct. Foods 2016, 23, 573–579. [Google Scholar] [CrossRef]
- Bustanji, Y.; Issa, A.; Mohammad, M.; Hudaib, M.; Tawah, K.; Alkhatib, H.; Almasri, I.; Al-Khalidi, B. Inhibition of hormone sensitive lipase and pancreatic lipase by Rosmarinus officinalis extract and selected phenolic constituents. J. Med. Plant Res. 2010, 4, 2235–2242. [Google Scholar]
- Denny, C.; Melo, P.S.; Franchin, M.; Massarioli, A.P.; Bergamaschi, K.B.; De Alencar, S.M.; Rosalen, P.L. Guava pomace: A new source of anti-inflammatory and analgesic bioactives. BMC Complement. Altern. Med. 2013, 13, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Khang, D.T.; Dung, T.N.; Elzaawely, A.A.; Xuan, T.D. Phenolic profiles and antioxidant activity of germinated legumes. Foods 2016, 5, 27. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Hong, G.-E.; Zhang, H.; Yang, C.-Y.; Han, K.-H.; Mandal, P.K.; Lee, C.-H. Production of the isoflavone aglycone and antioxidant activities in black soymilk using fermentation with Streptococcus thermophilus S10. Food Sci. Biotechnol. 2015, 24, 537–544. [Google Scholar] [CrossRef]
- Gosslau, A.; Li, S.; Ho, C.T.; Chen, K.Y.; Rawson, N.E. The importance of natural product characterization in studies of their anti-inflammatory activity. Mol. Nutr. Food Res. 2011, 55, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Amarowicz, R.; Shahidi, F. Antioxidant activity of faba bean extract and fractions thereof. J. Food Bioact. 2018, 2, 112–118. [Google Scholar]
- Larkin, T.; Price, W.E.; Astheimer, L. The key importance of soy isoflavone bioavailability to understanding health benefits. Crit. Rev. Food Sci. Nutr. 2008, 48, 538–552. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Faughnan, M.; Hoey, L.; Wähälä, K.; Williamson, G.; Cassidy, A. Bioavailability of phyto-oestrogens. Br. J. Nutr. 2003, 89, S45–S58. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | Variation Levels | ||||
|---|---|---|---|---|---|
| −1.41 | −1 | 0 | +1 | +1.41 | |
| X1 = Temperature (°C) | 23 | 25 | 30 | 35 | 37 |
| X2 = pH | 3.6 | 4.0 | 5.0 | 6.0 | 6.4 |
| Independent Variables | Variation Levels | ||||
|---|---|---|---|---|---|
| −1.41 | −1 | 0 | +1 | +1.41 | |
| X3 = Temperature (°C) | 13.9 | 20.0 | 35.0 | 50.0 | 56.2 |
| X4 = pH | 3.39 | 4.00 | 5.50 | 7.00 | 7.61 |
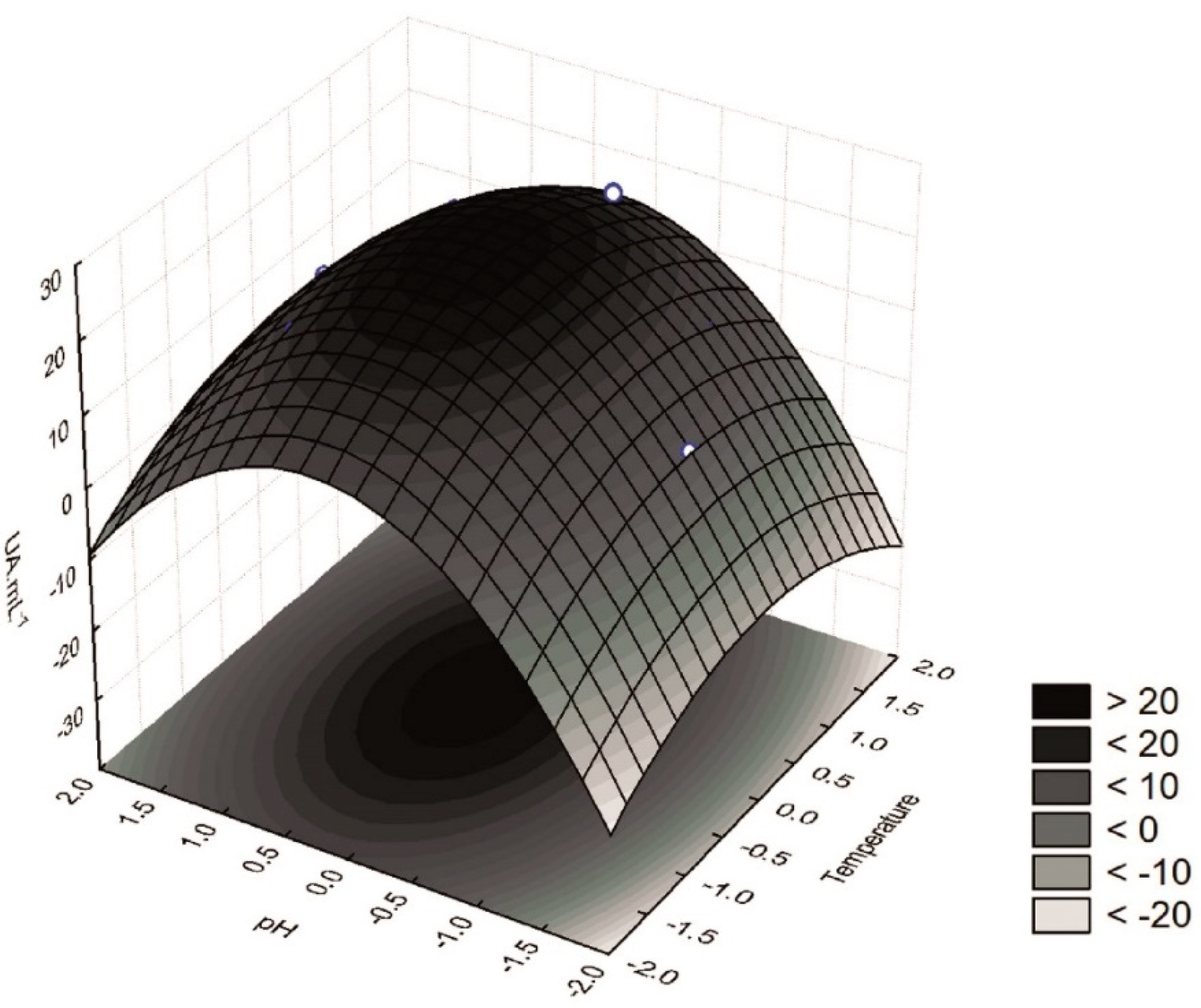
| Variation Source | SS | DF | MS | F Test | p | R2 |
|---|---|---|---|---|---|---|
| X1 (T) (linear) | 0.558 | 1 | 0.558 | 0.659 | 0.428 | 0.94 |
| X1 (T) (quadratic) | 19.038 | 1 | 19.038 | 22.492 | 0.002 | |
| X2 (pH) (linear) | 62.340 | 1 | 62.340 | 73.650 | 0.000 | |
| X2 (pH) (quadratic) | 137.038 | 1 | 137.038 | 161.900 | 0.000 | |
| Interaction X1X2 | 0.120 | 1 | 0.120 | 0.142 | 0.711 | |
| Error | 13.543 | 16 | 0.846 | |||
| Total | 214.527 | 21 |
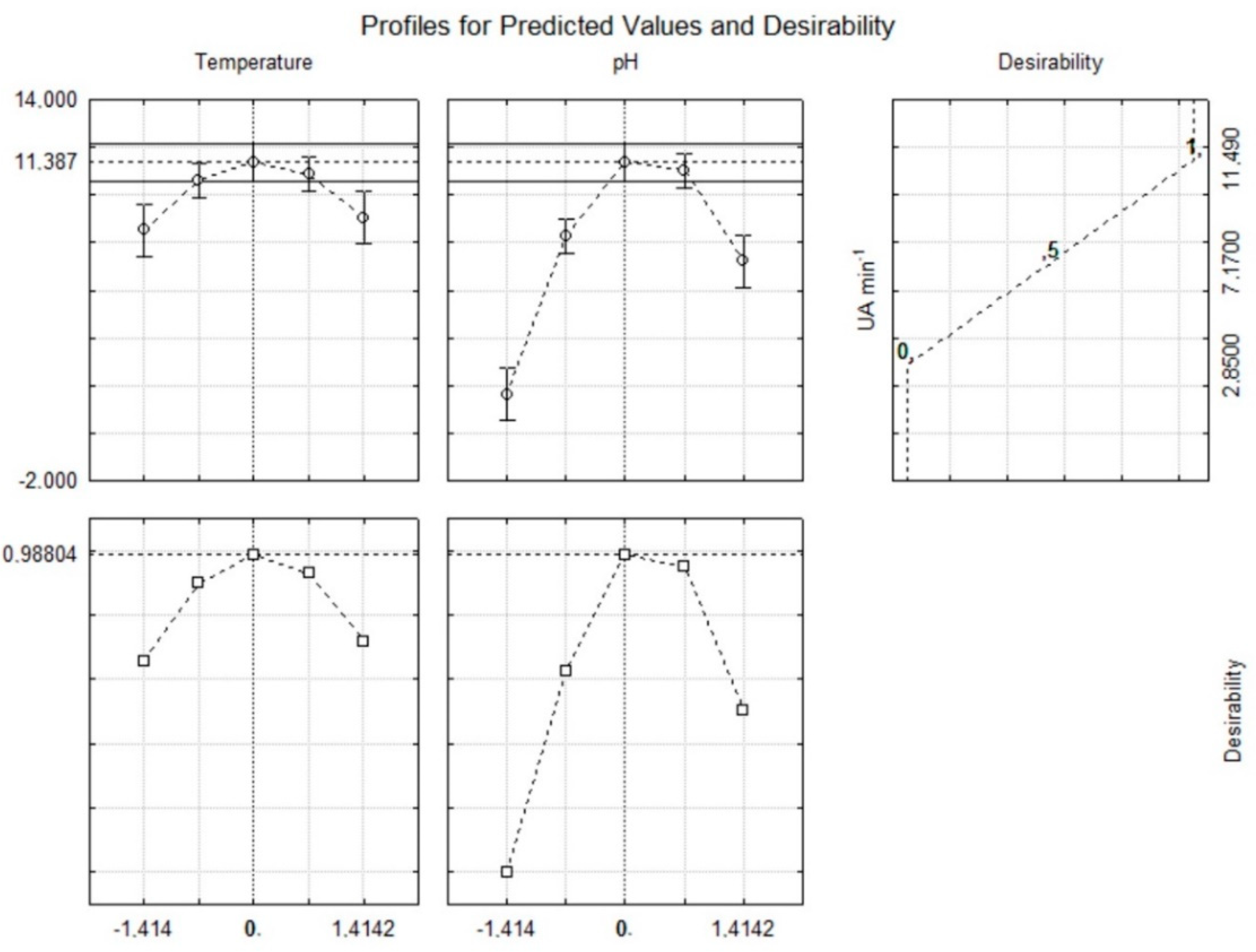
| Assays | Coded Variables | Decoded Variable | Response Function (Y) | ||
|---|---|---|---|---|---|
| x1 | x2 | T (°C) (X1) | pH (X2) | β-Glucosidase Activity (UA mL−1) | |
| 1 | −1 | −1 | 25.0 | 4.0 | 8.16 |
| 2 | −1 | 1 | 25.0 | 6.0 | 17.7 |
| 3 | 1 | −1 | 35.0 | 4.0 | 6.22 |
| 4 | 1 | 1 | 35.0 | 6.0 | 16.7 |
| 5 | 0 | 0 | 30.0 | 5.0 | 22.4 |
| 6 | 0 | 0 | 30.0 | 5.0 | 23.0 |
| 7 | 0 | 0 | 30.0 | 5.0 | 23.0 |
| 8 | −1.41 | 0 | 23.0 | 5.0 | 16.5 |
| 9 | 1.41 | 0 | 37.0 | 5.0 | 20.7 |
| 10 | 0 | −1.41 | 30.0 | 3.6 | 5.76 |
| 11 | 0 | 1.41 | 30.0 | 6.4 | 13.9 |
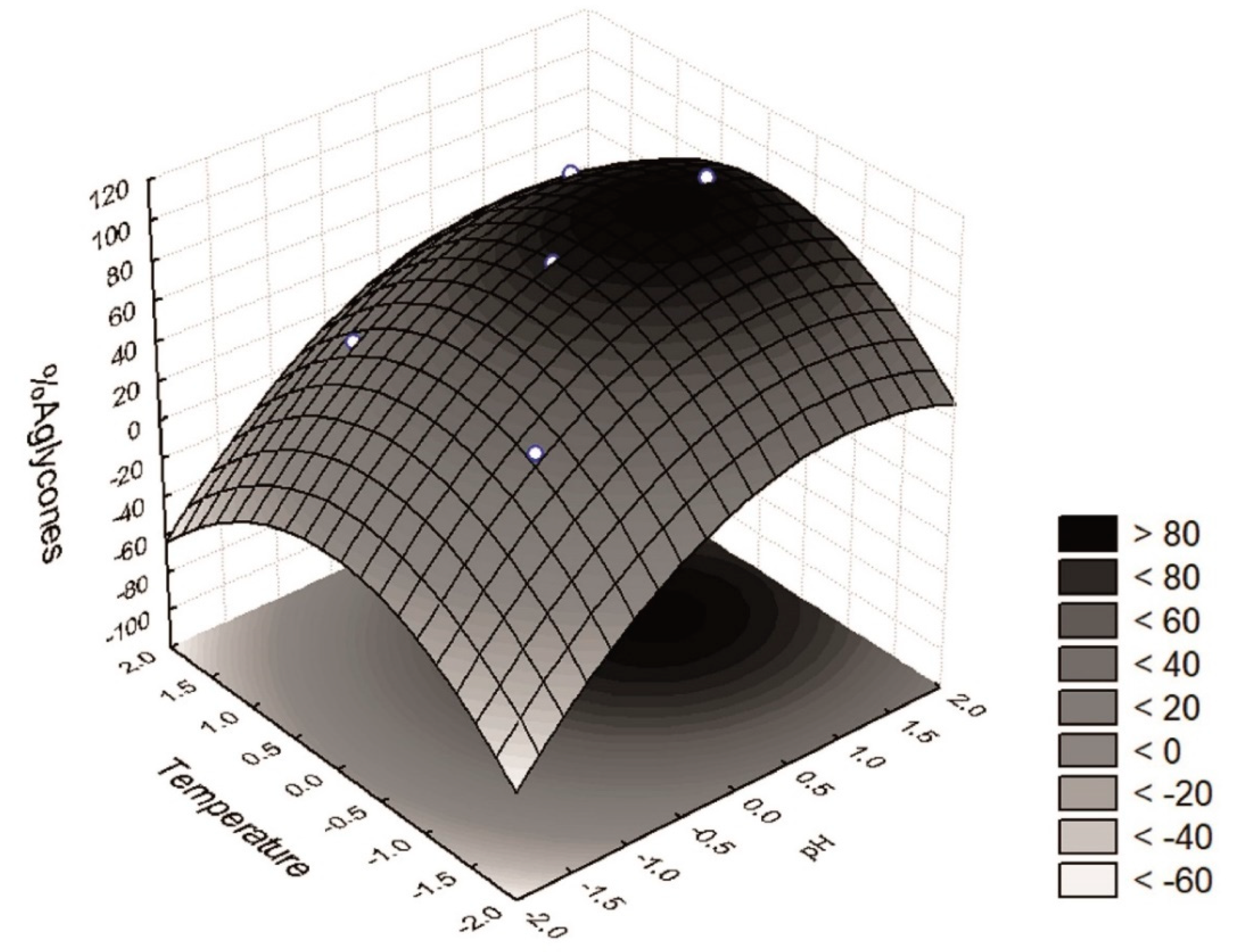
| Variation Source | SS | DF | MS | F test | p | R2 |
|---|---|---|---|---|---|---|
| Block | 22.050 | 1 | 22.050 | 0.17864 | 0.683680 | 0.86 |
| (X4) pH (Linear) | 4217.201 | 1 | 4217.201 | 34.16513 | 0.000385 | |
| (X4) pH (Quadratic) | 872.759 | 1 | 872.759 | 7.07055 | 0.028848 | |
| (X3) T (Quadratic) | 1328.402 | 1 | 1328.402 | 10.76189 | 0.011179 | |
| Error | 987.486 | 8 | 123.436 | |||
| Total | 7231.600 | 12 |
| Assays | Block | Coded Variables | Decoded Variables | Response Function (W) | ||
|---|---|---|---|---|---|---|
| x3 | x4 | T (°C) (X3) | pH (X4) | % Aglycones * | ||
| 1 | 1 | −1 | −1 | 20.0 | 4.00 | 47.5 |
| 2 | 1 | +1 | −1 | 50.0 | 4.00 | 44.6 |
| 3 | 1 | −1 | +1 | 20.0 | 7.00 | 68.6 |
| 4 | 1 | +1 | +1 | 50.0 | 7.00 | 84.0 |
| 5 (c) | 1 | 0 | 0 | 35.0 | 5.50 | 88.6 |
| 6 (c) | 1 | 0 | 0 | 35.0 | 5.50 | 79.0 |
| 7 (c) | 1 | 0 | 0 | 35.0 | 5.50 | 88.6 |
| 8 | 2 | 0 | −1.41 | 35.0 | 3.39 | 11.6 |
| 9 | 2 | 0 | +1.41 | 35.0 | 7.61 | 98.7 |
| 10 | 2 | −1.41 | 0 | 13.9 | 5.50 | 48.6 |
| 11 | 2 | +1.41 | 0 | 56.2 | 5.50 | 47.1 |
| 12 (c) | 2 | 0 | 0 | 35.0 | 5.50 | 85.0 |
| 13 (c) | 2 | 0 | 0 | 35.0 | 5.50 | 88.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshiara, L.Y.; Madeira, T.B.; De Camargo, A.C.; Shahidi, F.; Ida, E.I. Multistep Optimization of ?-Glucosidase Extraction from Germinated Soybeans (Glycine max L. Merril) and Recovery of Isoflavone Aglycones. Foods 2018, 7, 110. https://doi.org/10.3390/foods7070110
Yoshiara LY, Madeira TB, De Camargo AC, Shahidi F, Ida EI. Multistep Optimization of ?-Glucosidase Extraction from Germinated Soybeans (Glycine max L. Merril) and Recovery of Isoflavone Aglycones. Foods. 2018; 7(7):110. https://doi.org/10.3390/foods7070110
Chicago/Turabian StyleYoshiara, Luciane Yuri, Tiago Bervelieri Madeira, Adriano Costa De Camargo, Fereidoon Shahidi, and Elza Iouko Ida. 2018. "Multistep Optimization of ?-Glucosidase Extraction from Germinated Soybeans (Glycine max L. Merril) and Recovery of Isoflavone Aglycones" Foods 7, no. 7: 110. https://doi.org/10.3390/foods7070110
APA StyleYoshiara, L. Y., Madeira, T. B., De Camargo, A. C., Shahidi, F., & Ida, E. I. (2018). Multistep Optimization of ?-Glucosidase Extraction from Germinated Soybeans (Glycine max L. Merril) and Recovery of Isoflavone Aglycones. Foods, 7(7), 110. https://doi.org/10.3390/foods7070110