Medium Chain Triglyceride (MCT) Oil Affects the Immunophenotype via Reprogramming of Mitochondrial Respiration in Murine Macrophages


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Oil Treatments
2.2. Animals and Dietary Interventions
2.3. Bone Marrow-Derived Macrophage (BMDM) Culture and Macrophage Polarization
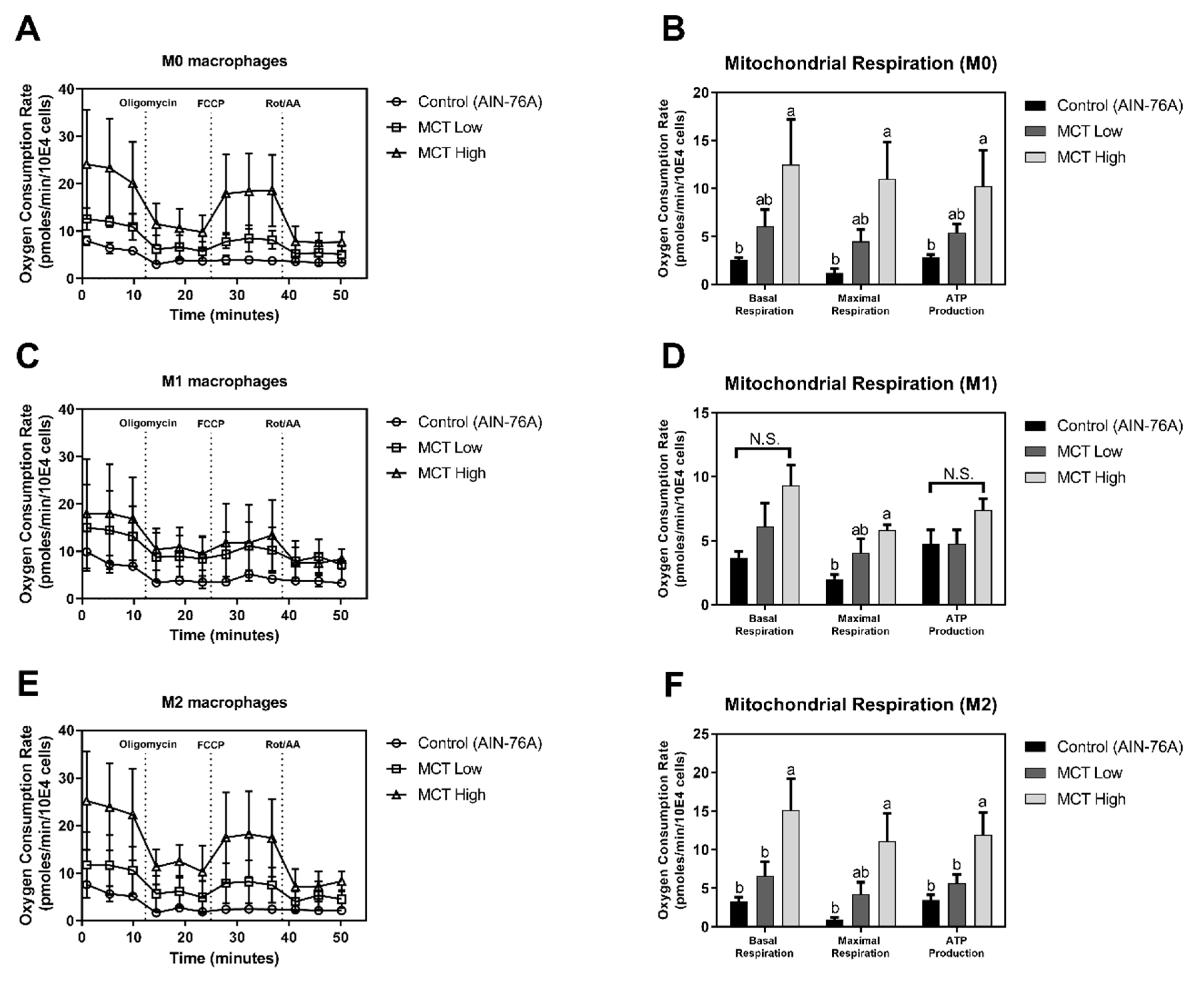
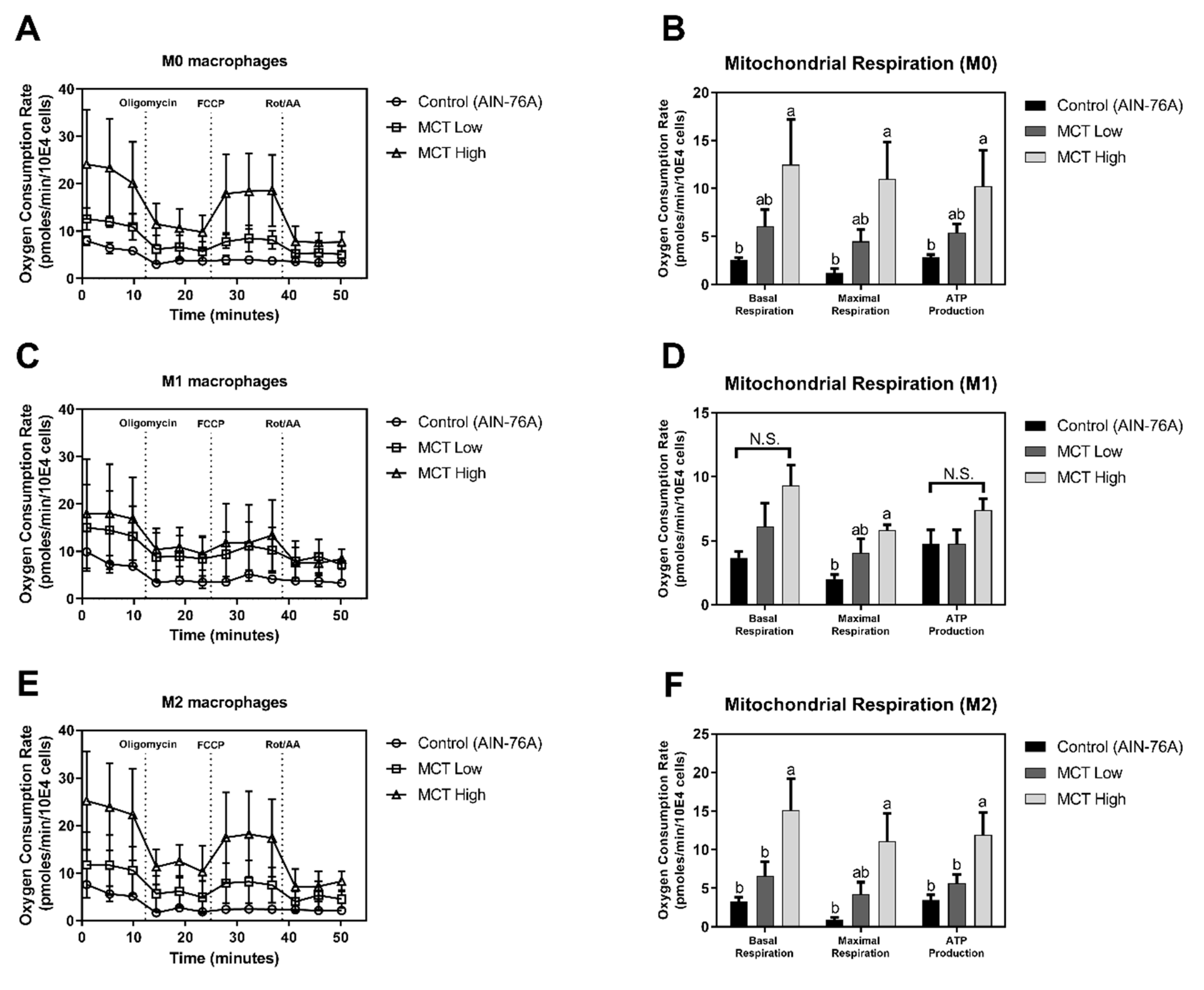
2.4. Measurement of Oxygen Consumption Rate
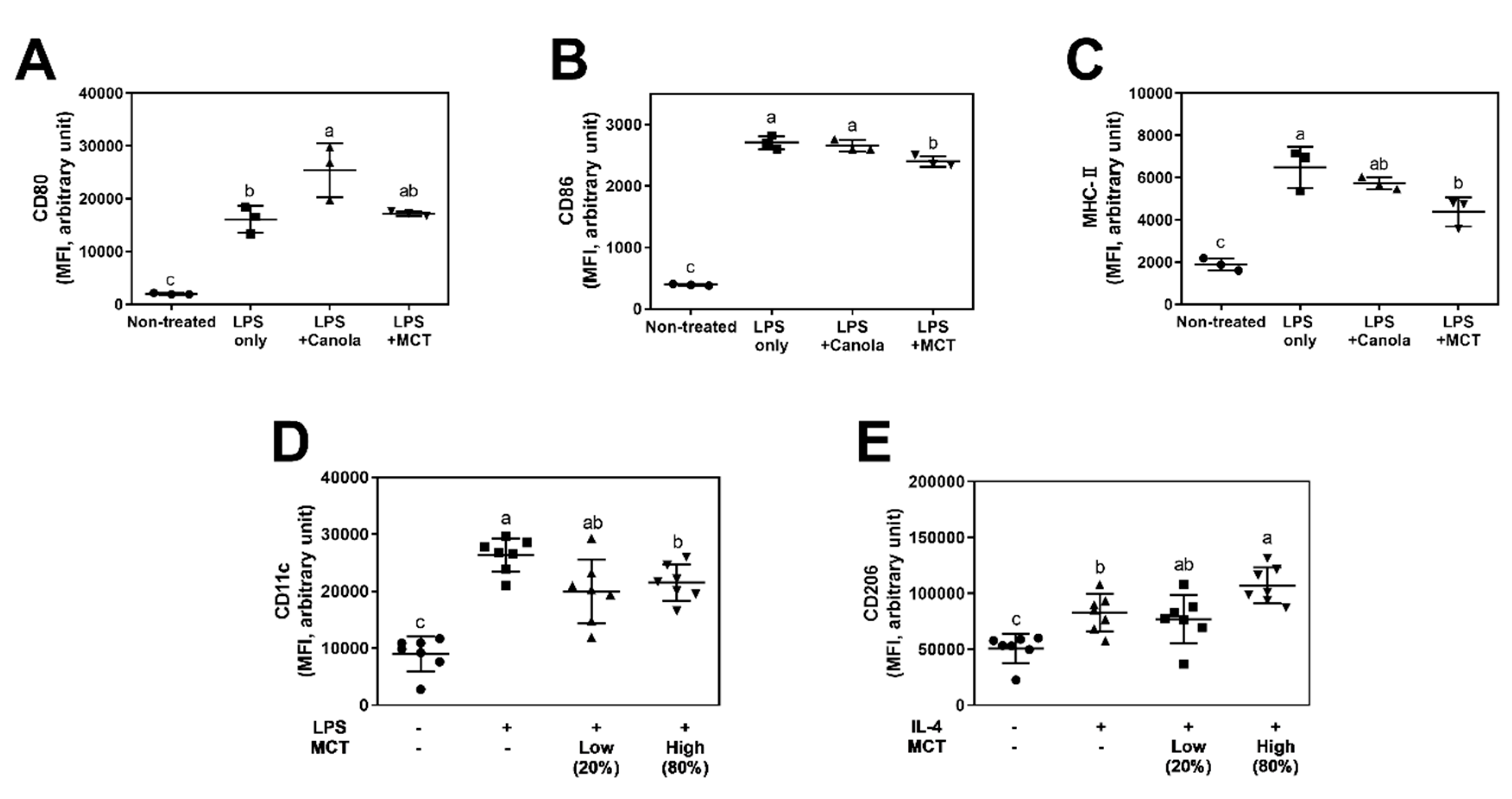
2.5. Assessments of Macrophage Activation Markers
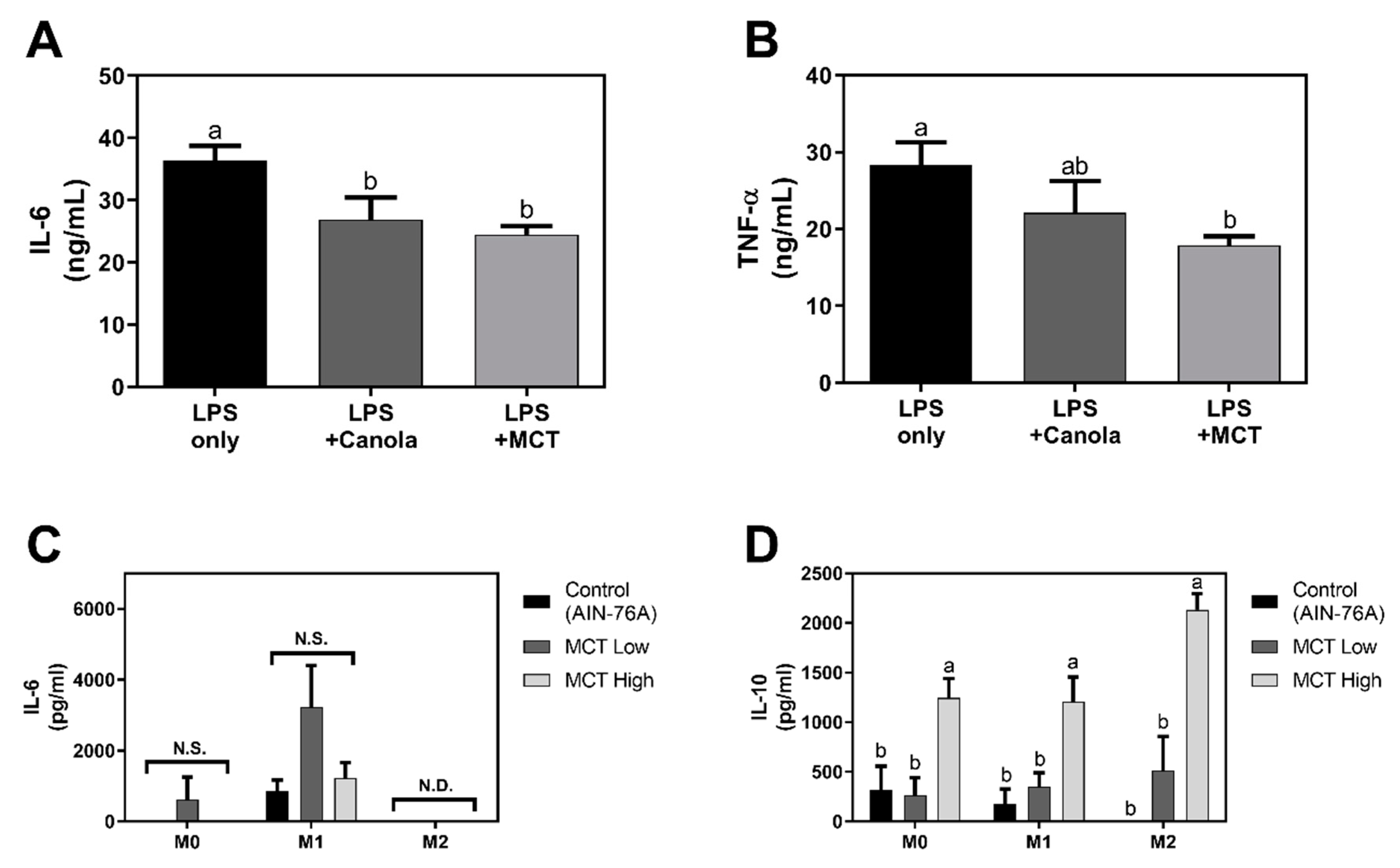
2.6. Cytokine Quantification
2.7. Statistical Analysis
2.8. Ethics Statement
3. Results
3.1. MCT Oil Up-Regulates Mitochondrial Respiration in Macrophages
3.2. MCT Oil Downregulates Activation Marker Expression on Macrophage Surfaces
3.3. MCT Oil Induces an Anti-Inflammatory M2-Like Phenotype in Macrophages
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- West, A.P.; Shadel, G.S.; Ghosh, S. Mitochondria in innate immune responses. Nat. Rev. Immunol. 2011, 11, 389–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, E.L.; O’Neill, L.A. Reprogramming mitochondrial metabolism in macrophages as an anti-inflammatory signal. Eur. J. Immunol. 2016, 46, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Geeraerts, X.; Bolli, E.; Fendt, S.M.S.-M.; Van Ginderachter, J.A.A. Macrophage Metabolism as Therapeutic Target for Cancer, Atherosclerosis, and Obesity. Front. Immunol. 2017, 8, 289. [Google Scholar] [CrossRef]
- Pike, L.S.; Wu, M. Rapid analysis of glycolytic and oxidative substrate flux of cancer cells in a microplate. PLoS ONE 2014, 9, e109916. [Google Scholar]
- Huang, S.-C.; Everts, B.; Ivanova, Y.; O’Sullivan, D.; Nascimento, M.; Smith, A.M.M.; Beatty, W.; Love-Gregory, L.; Lam, W.Y.Y.; O’Neill, C.M.M.; et al. Cell-intrinsic lysosomal lipolysis is essential for alternative activation of macrophages. Nat. Immunol. 2014, 15, 846–855. [Google Scholar] [CrossRef] [Green Version]
- Qu, Q.; Zeng, F.; Liu, X.; Wang, Q.J.; Deng, F. Fatty acid oxidation and carnitine palmitoyltransferase I: Emerging therapeutic targets in cancer. Cell Death Dis. 2016, 7, e2226. [Google Scholar] [CrossRef]
- Nagao, K.; Yanagita, T. Medium-chain fatty acids: Functional lipids for the prevention and treatment of the metabolic syndrome. Pharm. Res. 2010, 61, 208–212. [Google Scholar] [CrossRef]
- Marten, B.; Pfeuffer, M.; Schrezenmeir, J. Medium-chain triglycerides. Int. Dairy J. 2006, 16, 1374–1382. [Google Scholar] [CrossRef]
- Yu, S.; Choi, J.H.; Kim, H.J.; Park, S.H.; Go, G.; Kim, W. In Vitro Evidence of Anti-Inflammatory and Anti-Obesity Effects of Medium-Chain Fatty Acid-Diacylglycerols. J. Microbiol. Biotechnol. 2017, 27, 1617–1627. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.; Yu, S.; Choi, G.S.; Kim, J.H.; Baik, M.; Su, S.T.; Kim, W. Puffing of Rehmannia glutinosa enhances anti-oxidant capacity and down-regulates IL-6 production in RAW 264.7 cells. Food Sci. Biotechnol. 2019, 28, 1235–1240. [Google Scholar] [CrossRef]
- Jablonski, K.A.; Amici, S.A.; Webb, L.M.; Ruiz-Rosado, J.D.D.; Popovich, P.G.; Partida-Sanchez, S.; Guerau-De-arellano, M. Novel markers to delineate murine M1 and M2 macrophages. PLoS ONE 2015, 10, e0145342. [Google Scholar] [CrossRef] [PubMed]
- Loganes, C.; Ballali, S.; Minto, C. Main Properties of Canola Oil Components: A Descriptive Review of Current Knowledge. Open Agric. J. 2016, 10, 69–74. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Cheng, G.; Hardy, M.; Ouari, O.; Lopez, M.; Joseph, J.; Zielonka, J.; Dwinell, M.B. A review of the basics of mitochondrial bioenergetics, metabolism, and related signaling pathways in cancer cells: Therapeutic targeting of tumor mitochondria with lipophilic cationic compounds. Redox Biol. 2018, 14, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Chung, S.H. Anti-inflammatory effect of α-linolenic acid and its mode of action through the inhibition of nitric oxide production and inducible nitric oxide synthase gene expression via NF-κB and mitogen-activated protein kinase pathways. J. Agric. Food Chem. 2007, 55, 5073–5080. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-C.; Zou, X.-B.; Chai, Y.-F.; Yao, Y.-M. Macrophage polarization in inflammatory diseases. Int. J. Biol. Sci. 2014, 10, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.; Kim, J.Y. Chondroprotective effect of curcumin and lecithin complex in human chondrocytes stimulated by IL-1β via an anti-inflammatory mechanism. Food Sci. Biotechnol. 2019, 28, 547–553. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Zhou, X.; Li, W.; Wang, S.; Zhang, P.; Wang, Q.; Xiao, J.; Zhang, C.; Zheng, X.; Xu, X.; Xue, S.; et al. YAP Aggravates Inflammatory Bowel Disease by Regulating M1/M2 Macrophage Polarization and Gut Microbial Homeostasis. Cell Rep. 2019, 27, 1176–1189. [Google Scholar] [CrossRef]
- Kelly, B.; O’Neill, L.A.J. Metabolic reprogramming in macrophages and dendritic cells in innate immunity. Cell Res. 2015, 25, 771–784. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, H.; Mao, C.; Sun, M.; Dominah, G.; Chen, L.; Zhuang, Z. Fatty acid oxidation contributes to IL-1β secretion in M2 macrophages and promotes macrophage-mediated tumor cell migration. Mol. Immunol. 2018, 94, 27–35. [Google Scholar] [CrossRef]
- Van den Bossche, J.; Baardman, J.; Otto, N.A.; van der Velden, S.; Neele, A.E.; van den Berg, S.M.; Luque-Martin, R.; Chen, H.J.; Boshuizen, M.C.S.; Ahmed, M.; et al. Mitochondrial Dysfunction Prevents Repolarization of Inflammatory Macrophages. Cell Rep. 2016, 17, 684–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.S.; Jung, D.Y.; Morel, C.; Lakhani, S.A.; Kim, J.K.; Flavell, R.A.; Davis, R.J. JNK expression by macrophages promotes obesity-induced insulin resistance and inflammation. Science 2013, 339, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Yoon, H.-J.; Kim, S.-Y.; Yoon, Y.-R. A Medium-Chain Fatty Acid, Capric Acid, Inhibits RANKL-Induced Osteoclast Differentiation via the Suppression of NF-κB Signaling and Blocks Cytoskeletal Organization and Survival in Mature Osteoclasts. Mol. Cells 2014, 37, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage activation and polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-Z.; Wang, X.; Wang, Y.; Niu, A.; Wang, S.; Zou, C.; Harris, R.C. IL-4/IL-13–mediated polarization of renal macrophages/dendritic cells to an M2a phenotype is essential for recovery from acute kidney injury. Kidney Int. 2017, 91, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Raschke, W.C.; Baird, S.; Ralph, P.; Nakoinz, I. Functional macrophage cell lines transformed by abelson leukemia virus. Cell 1978, 15, 261–267. [Google Scholar] [CrossRef]
- Wu, W.H.H.; Lin, B.Y.Y.; Kuo, Y.H.H.; Huang, C. Triglycerides constituted of short and medium chain fatty acids and dicarboxylic acids in Momordica charantia, as well as capric acid, inhibit PGE2 production in RAW264.7 macrophages. Food Chem. 2009, 117, 306–311. [Google Scholar] [CrossRef]
- Bertevello, P.L.L.; De Nardi, L.; Torrinhas, R.S.S.; Logullo, A.F.F.; Waitzberg, D.L.L. Partial Replacement of omega-6 Fatty Acids with Medium-Chain Triglycerides, but Not Olive Oil, Improves Colon Cytokine Response and Damage in Experimental Colitis. J. Parenter. Enter. Nutr. 2012, 36, 442–448. [Google Scholar] [CrossRef]
- Kono, H.; Fujii, H.; Ogiku, M.; Tsuchiya, M.; Ishii, K.; Hara, M. Enteral diets enriched with medium-chain triglycerides and N-3 fatty acids prevent chemically induced experimental colitis in rats. Transl. Res. 2010, 156, 282–291. [Google Scholar] [CrossRef]
- Druml, W.; Fishcer, M.; Pidlich, J.; Lenz, K. Fat elimination in chronic hepatic failure: Long-chain vs. medium-chain triglycerides. Am. J. Clin. Nutr. 1995, 61, 812–817. [Google Scholar] [CrossRef]
- Kadochi, Y.; Mori, S.; Fujiwara-Tani, R.; Luo, Y.; Nishiguchi, Y.; Kishi, S.; Fujii, K.; Ohmori, H.; Kuniyasu, H. Remodeling of energy metabolism by a ketone body and medium-chain fatty acid suppressed the proliferation of CT26 mouse colon cancer cells. Oncol. Lett. 2017, 14, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, H.; Ogawa, A.; Kasai, M.; AOYAMA, T. Effect of Randomly Interesterified Triacylglycerols Containing Medium- and Long-Chain Fatty Acids on Energy Expenditure and Hepatic Fatty Acid Metabolism in Rats. Biosci. Biotechnol. Biochem. 2005, 69, 1811–1818. [Google Scholar] [CrossRef] [PubMed]
- Schönfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Dayrit, F.M. Lauric acid is a medium-chain fatty acid, coconut oil is a medium-chain triglyceride. Philipp. J. Sci. 2014, 143, 157–166. [Google Scholar]
- Nomura, M.M.; Liu, J.; Rovira, I.I.I.; Gonzalez-Hurtado, E.; Lee, J.; Wolfgang, M.J.J.; Finkel, T. Fatty acid oxidation in macrophage polarization. Nat. Immunol. 2016, 17, 216–217. [Google Scholar] [CrossRef]
- Johnson, A.R.; Qin, Y.; Cozzo, A.J.; Freemerman, A.J.; Huang, M.J.; Zhao, L.; Sampey, B.P.; Milner, J.J.; Beck, M.A.; Damania, B.; et al. Metabolic reprogramming through fatty acid transport protein 1 (FATP1) regulates macrophage inflammatory potential and adipose inflammation. Mol. Metab. 2016, 5, 506–526. [Google Scholar] [CrossRef]
- Huang, S.C.C.; Smith, A.M.; Everts, B.; Colonna, M.; Pearce, E.L.E.J.; Schilling, J.D.; Pearce, E.L.E.J. Metabolic Reprogramming Mediated by the mTORC2-IRF4 Signaling Axis Is Essential for Macrophage Alternative Activation. Immunity 2016, 45, 817–830. [Google Scholar] [CrossRef] [Green Version]
- Wanten, G.J.A.; Naber, A.H.J.; Kruimel, J.W.; Tool, A.T.J.; Roos, D.; Jansen, J.B.M.J. Influence of structurally different lipid emulsions on human neutrophil oxygen radical production. Eur. J. Clin. Investig. 1999, 29, 357–363. [Google Scholar] [CrossRef]
- Wanten, G.J.A.; Geijtenbeek, T.B.H.; Raymakers, R.A.P.; van Kooyk, Y.; Roos, D.; Jansen, J.B.M.J.; Naber, A.H.J. Medium-chain, triglyceride-containing lipid emulsions increase human neutrophil β2 integrin expression, adhesion, and degranulation. J. Parenter. Enter. Nutr. 2000, 24, 228–233. [Google Scholar] [CrossRef]
- Wanten, G.J.A.; Roos, D.; Naber, A.H.J. Effects of structurally different lipid emulsions on human neutrophil migration. Clin. Nutr. 2000, 19, 327–331. [Google Scholar] [CrossRef]
- Wanten, G.J.; Curfs, J.H.; Meis, J.F.; Naber, A.H. Phagocytosis and killing of Candida albicans by human neutrophils after exposure to structurally different lipid emulsions. J. Parenter. Enter. Nutr. 2001, 25, 9–13. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Normal (AIN-76A) grams/kg | High-MCT grams/kg | Low-MCT grams/kg |
|---|---|---|---|
| Casein | 200 | 200 | 200 |
| dl-methionine | 3 | 3 | 3 |
| Corn Starch | 150 | 150 | 150 |
| Sucrose | 500 | 500 | 500 |
| Cellulose | 50 | 50 | 50 |
| Corn oil | 50 | 10 | 40 |
| Medium-chain fatty acid oil | 0 | 40 | 10 |
| Mineral mix | 35 | 35 | 35 |
| Vitamin mix | 10 | 10 | 10 |
| Choline bitartrate | 2 | 2 | 2 |
| Total | 1000 | 1000 | 1000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Go, G.-w.; Kim, W. Medium Chain Triglyceride (MCT) Oil Affects the Immunophenotype via Reprogramming of Mitochondrial Respiration in Murine Macrophages. Foods 2019, 8, 553. https://doi.org/10.3390/foods8110553
Yu S, Go G-w, Kim W. Medium Chain Triglyceride (MCT) Oil Affects the Immunophenotype via Reprogramming of Mitochondrial Respiration in Murine Macrophages. Foods. 2019; 8(11):553. https://doi.org/10.3390/foods8110553
Chicago/Turabian StyleYu, Seungmin, Gwang-woong Go, and Wooki Kim. 2019. "Medium Chain Triglyceride (MCT) Oil Affects the Immunophenotype via Reprogramming of Mitochondrial Respiration in Murine Macrophages" Foods 8, no. 11: 553. https://doi.org/10.3390/foods8110553
APA StyleYu, S., Go, G.-w., & Kim, W. (2019). Medium Chain Triglyceride (MCT) Oil Affects the Immunophenotype via Reprogramming of Mitochondrial Respiration in Murine Macrophages. Foods, 8(11), 553. https://doi.org/10.3390/foods8110553