Preventive Effect of Blueberry Extract on Liver Injury Induced by Carbon Tetrachloride in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of BE
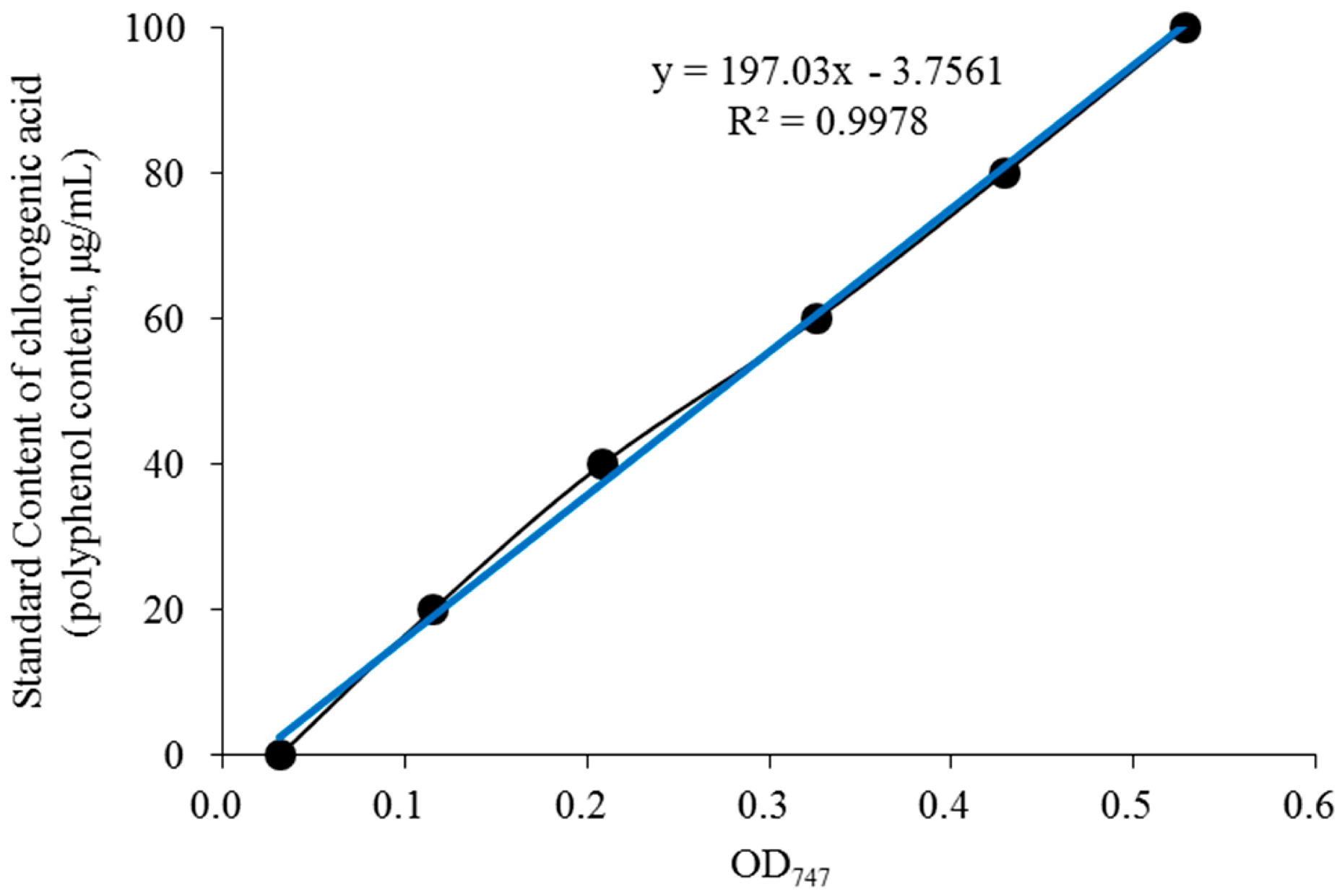
2.2. Determination of the Polyphenol Content in BE
2.3. Mouse Experiment
2.4. Determination of the Serum Aspartate Aminotransferase (AST), Alanine Aminotransferase (ALT), Triglyceride (TG) and Total Cholesterol (TC) Levels
2.5. Determination of The Serum Cytokine Interleukin-6 (IL-6), Tumor Necrosis Factor-α (TNF-α) and Interferon-γ (IFN-γ) Levels
2.6. Determination of the SOD and TBARS Levels in Liver Tissue
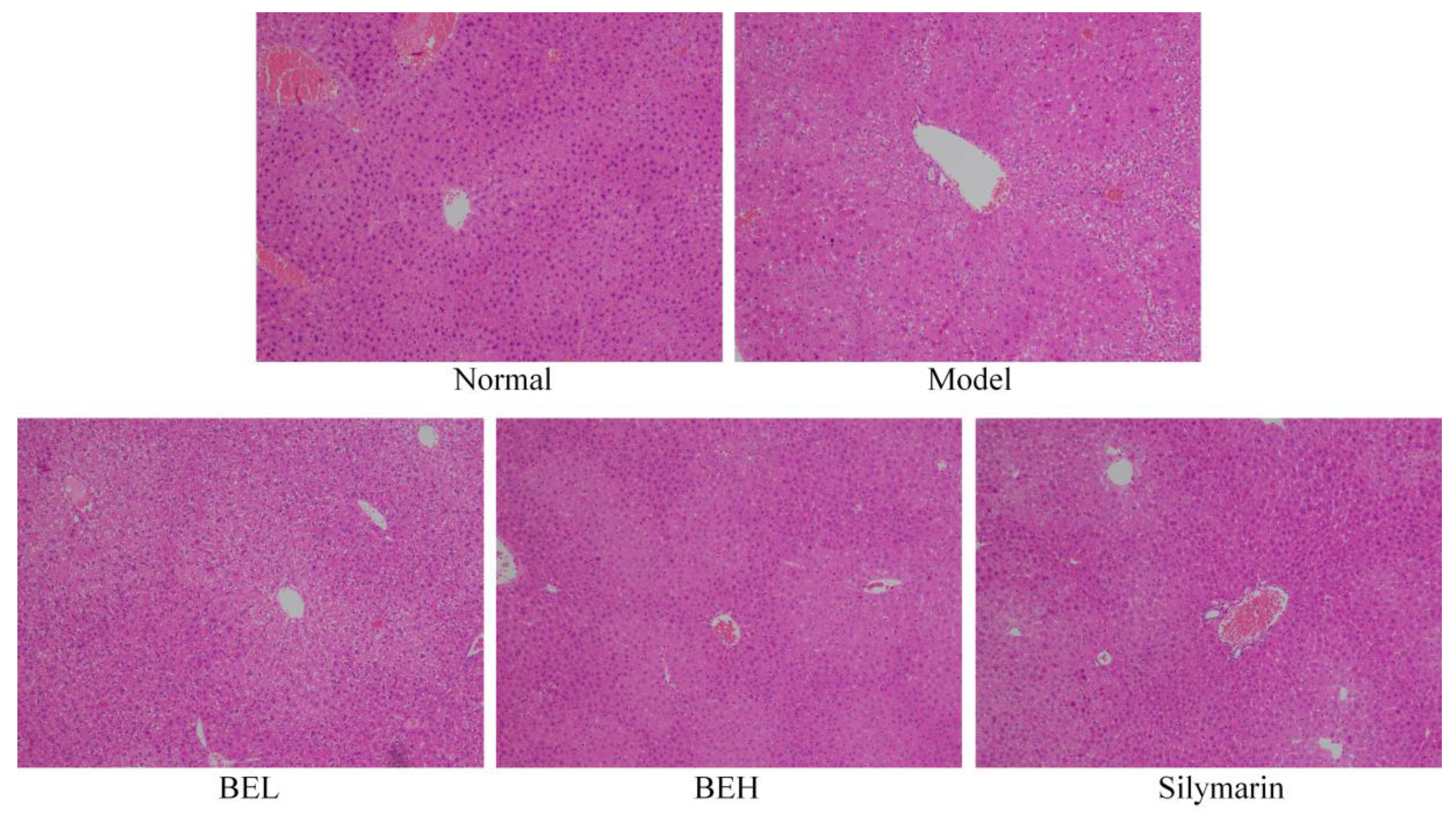
2.7. Pathological Observation of Liver Tissue
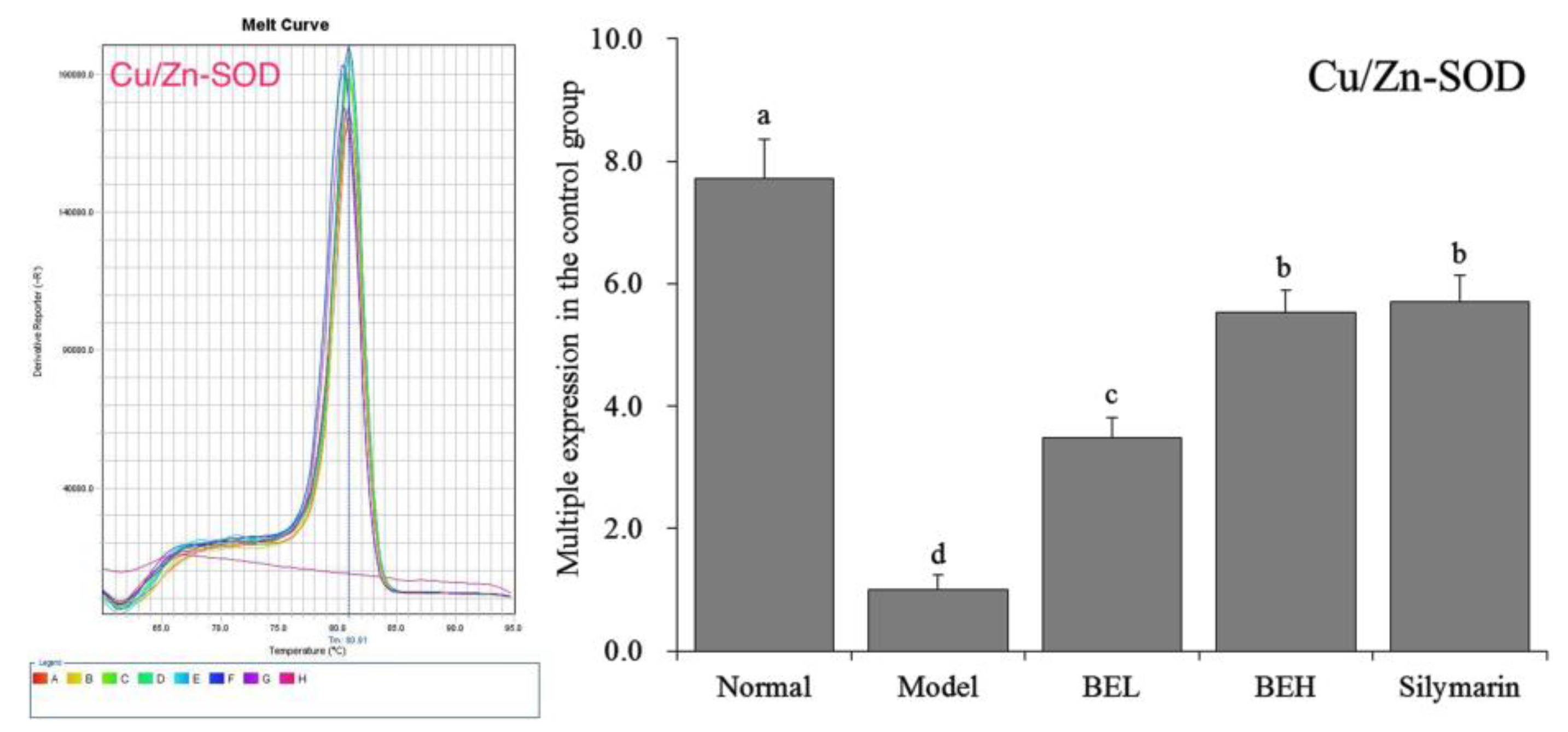
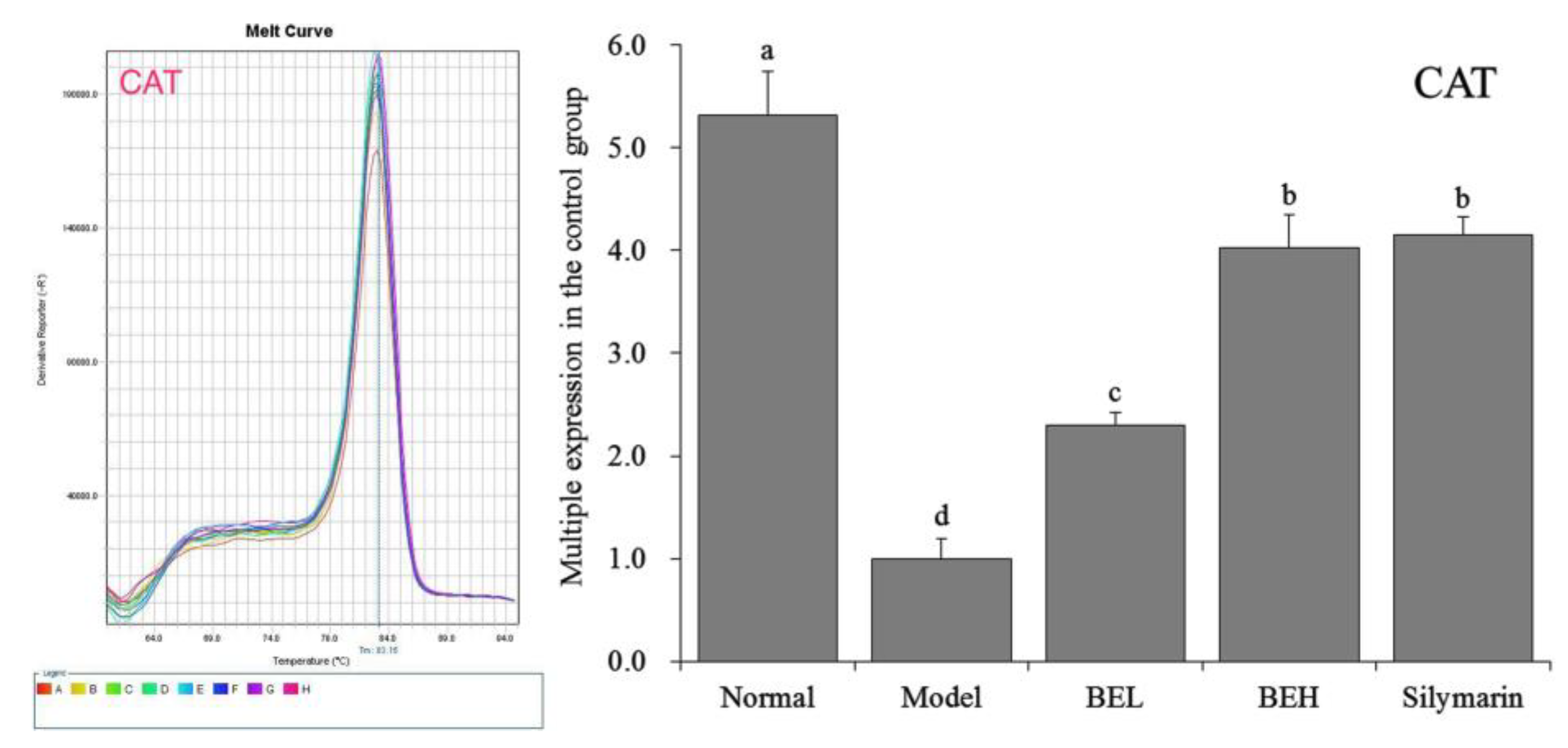
2.8. Quantitative PCR (qPCR) Assay
2.9. Statistical Analysis
3. Results
3.1. Polyphenol Content in the BE Extract
3.2. The Body Weight, Liver Weight and Liver Index
3.3. Mouse Serum AST, ALT, TG and TC Levels
3.4. Serum Cytokine IL-6, TNF-α and IFN-γ Levels
3.5. SOD Activity and TBARS Level in Mouse Liver Tissue
3.6. Pathological Observation of Mouse Liver
3.7. Cu/Zn-SOD mRNA Expression in Mouse Liver Tissue
3.8. Mn-SOD mRNA Expression in Mouse Liver Tissue
3.9. CAT mRNA Expression in Mouse Liver Tissue
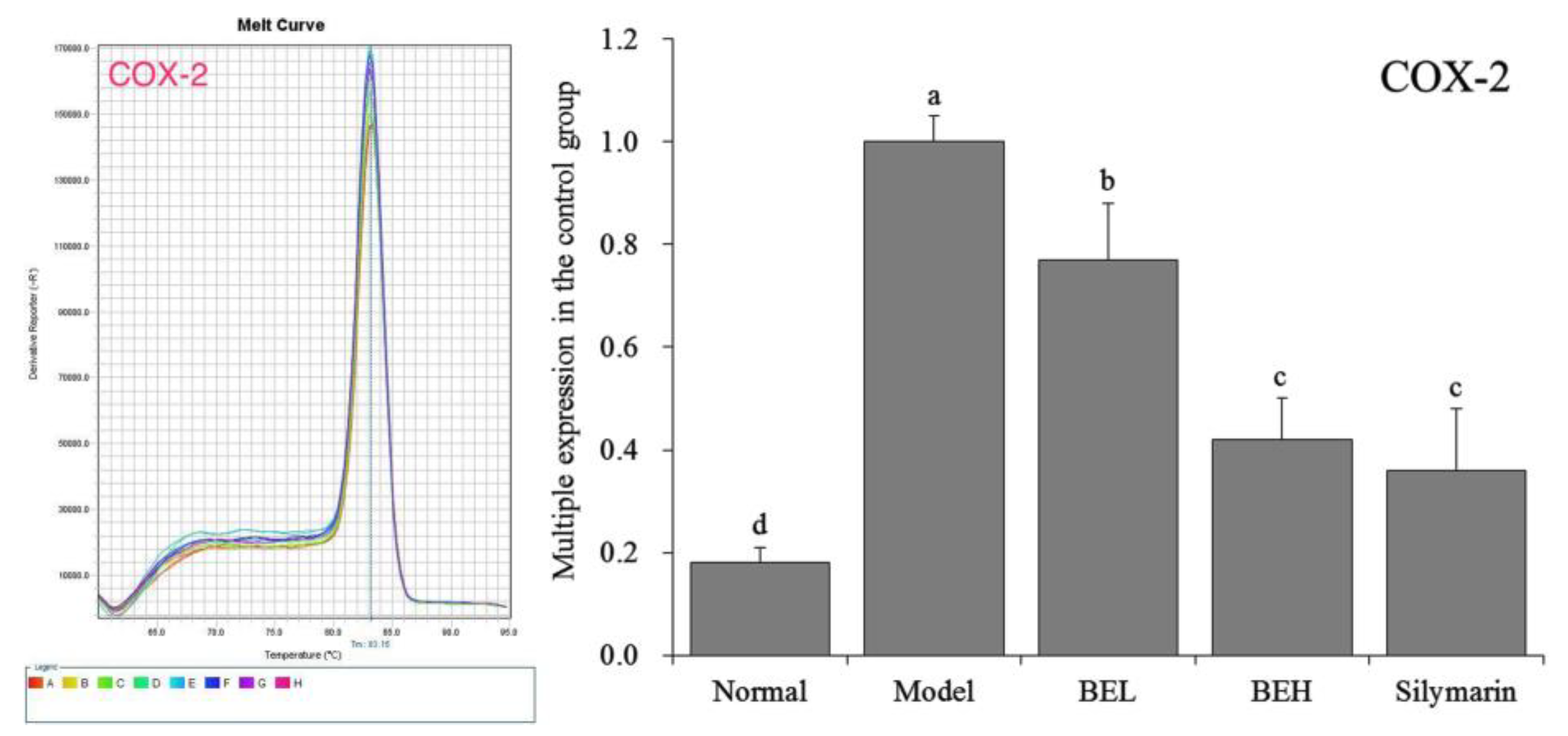
3.10. COX-2 mRNA Expression in Mouse Liver Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wilson, M.A.; Shukitt-Hale, B.; Kalt, W.; Ingram, D.K.; Joseph, J.A.; Wolkow, C.A. Blueberry polyphenols increase lifespan and thermotolerance in Caenorhabditis elegans. Aging Cell 2006, 5, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.H.; Racicot, K.; Pilkenton, S.J.; Kwon, Y.I.; Apostolidis, E. Blueberry extract inhibits carbohydrate-hydrolyzing enzymes and these inhibitory activities are not proanthocyanidin dependent. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 127–136. [Google Scholar] [CrossRef]
- Rimando, A.M.; Khan, S.I.; Mizuno, C.S.; Ren, G.; Mathews, S.T.; Kim, H.; Yokoyama, W. Evaluation of PPARα activation by known blueberry constituents. J. Sci. Food Agric. 2015, 96, 1666–1671. [Google Scholar] [CrossRef] [PubMed]
- Zhan, W.W.; Si, Z.J.; Wang, C.P.; Zeng, Q.P. Study on extracting and separation technology of proanthocyanidins from blueberry leaves. J. Cereal Oil 2010, 2010, 39–42. [Google Scholar]
- Correa-Betanzo, J.; Allen-Vercoe, E.; McDonald, J.; Schroeter, K.; Corredig, M.; Paliyath, G. Stability and biological activity of wild blueberry (Vaccinium angustifolium) polyphenols during simulated in vitro gastrointestinal digestion. Food Chem. 2014, 165, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Giacalone, M.; Di Sacco, F.; Traupe, I.; Topini, R.; Forfori, F.; Giunta, F. Antioxidant and neuroprotective properties of blueberry polyphenols: a critical review. Nutr. Neurosci. 2011, 14, 119–125. [Google Scholar] [CrossRef]
- Su, X.; Zhang, J.; Wang, H.; Xu, J.; He, J.; Liu, L.; Zhang, T.; Chen, R.; Kang, J. Phenolic acid profiling, antioxidant, and anti-inflammatory activities, and miRNA regulation in the polyphenols of 16 blueberry samples from China. Molecules 2017, 22, 312. [Google Scholar] [CrossRef]
- Soccio, M.; Laus, M.N.; Alfarano, M.; Dalfino, G.; Panunzio, M.F.; Pastore, D. Antioxidant/Oxidant Balance as a novel approach to evaluate the effect on serum of long-term intake of plant antioxidant-rich foods. J. Funct. Food 2018, 40, 778–784. [Google Scholar] [CrossRef]
- Harman, D. Free radical theory of aging: The “free radical” diseases. Age 1984, 7, 111–131. [Google Scholar] [CrossRef]
- DiSilvestro, R.A.; Carlson, G.P. Inflammation, an inducer of metallothionein, inhibits carbon-tetrachloride-induced hepatotoxicity in rats. Toxicol. Lett. 1992, 60, 175–181. [Google Scholar] [CrossRef]
- Slater, T.F.; Cheeseman, K.H.; Ingold, K.U. Carbon tetrachloride toxicity as a model for studying free-radical mediated liver injury. Philos. Trans. R Soc. Lond B Biol. Sci. 1985, 311, 633–645. [Google Scholar] [CrossRef]
- Muriel, P. Role of free radicals in liver diseases. Hepatol. Int. 2009, 3, 526–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martynenko, A.; Astatkie, T.; Satanina, V. Novel hydrothermodynamic food processing technology. J. Food Eng. 2015, 152, 8–16. [Google Scholar] [CrossRef]
- Satanina, V.; Kalt, W.; Astatkie, T.; Havard, P.; Martynenko, A. Comparison of anthocyanin concentration in blueberries processed using hydrothermodynamic technology and conventional processing technologies. J. Food Process Eng. 2014, 37, 609–618. [Google Scholar] [CrossRef]
- Song, J.L.; Zhu, K.; Feng, X.; Zhao, X. Protective effect Malus pumila Mill leaf polyphenols in reserpineinduced gastric ulcer in mice. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 249–256. [Google Scholar] [CrossRef]
- Zhao, X.; Qian, Y. Preventive effects of Kuding tea crude polyphenols in DSS-induced C57BL/6J mice ulcerative colitis. Sci. Technol. Food Ind. 2017, 38, 357–362. [Google Scholar]
- Wang, R.; Feng, X.; Zhu, K.; Zhao, X.; Suo, H. Preventive activity of banana peel polyphenols on CCl4-induced experimental hepatic injury in Kunming mice. Exp. Ther. Med. 2016, 11, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, B.; Liu, B.; Zhou, X.; Mu, J.; Wang, Q.; Zhao, X.; Yang, Z. Preventive effect of Lactobacillus fermentum CQPC03 on activated carbon-induced constipation in ICR mice. Medicina 2018, 54, 89. [Google Scholar] [CrossRef]
- Qian, Y.; Zhang, J.; Zhou, X.; Yi, R.; Mu, J.; Long, X.; Pan, Y.; Zhao, X.; Liu, W. Lactobacillus plantarum CQPC11 isolated from sichuan pickled cabbages antagonizes D-galactose-induced oxidation and aging in mice. Molecules 2018, 23, 3026. [Google Scholar] [CrossRef]
- Qian, Y.; Zhang, J.; Fu, X.; Yi, R.; Sun, P.; Zou, M.; Long, X.; Zhao, X. Preventive effect of raw liubao tea polyphenols on mouse gastric injuries induced by HCl/ethanol via anti-oxidative stress. Molecules 2018, 23, 2848. [Google Scholar] [CrossRef]
- Wang, L.; Cheng, D.; Wang, H.; Di, L.; Zhou, X.; Xu, T.; Yang, X.; Liu, Y. The hepatoprotective and antifibrotic effects of Saururus chinensis against carbon tetrachloride induced hepatic fibrosis in rats. J. Ethnopharmacol. 2009, 126, 487–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Long, X.; Yi, R.; Zhao, X. Polyphenols in Liubao tea can prevent CCl4-Induced hepatic damage in mice through its antioxidant capacities. Nutrients 2018, 10, 1280. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.L.; Zhang, B.; Xing, G.; Wang, F.; Hu, Z. Protective effect of naringenin against acetaminophen-induced acute liver injury in metallothionein (MT)-null mice. Food Funct. 2013, 4, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Kongo, M.; Sasaki, E.; Nishida, K.; Ishiguro, I. Therapeutic effect of melatonin on carbon tetrachloride-induced acute liver injury in rats. J. Pineal Res. 2000, 28, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Masaki, T.; Chiba, S.; Tatsukawa, H.; Yasuda, T.; Noguchi, H.; Seike, M.; Yoshimatsu, H. Adiponectin protects LPS-induced liver injury through modulation of TNF-alpha in KK-Ay obese mice. Hepatology 2004, 40, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.W.; Yue, L. Effect and mechanisms of FR167653, a dual inhibitor of TNF-alpha and IL-1, on BCG plus LPS induced-liver injury. Inflamm. Res. 2005, 54, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Ohta, A.; Sekimoto, M.; Sato, M.; Koda, T.; Nishimura, S.; Iwakura, Y.; Sekikawa, K.; Nishimura, T. Indispensable role for TNF-alpha and IFN-gamma at the effector phase of liver injury mediated by Th1 cells specific to hepatitis B virus surface antigen. J. Immunol. 2000, 165, 956–961. [Google Scholar] [CrossRef]
- Campo, G.M.; Squadrito, F.; Ceccarelli, S.; Calò, M.; Avenoso, A.; Campo, S.; Squadrito, G.; Altavilla, D. Reduction of carbon tetrachloride-induced rat liver injury by IRFI 042, a novel dual vitamin E-like antioxidant. Free Radic. Res. 2001, 34, 379–393. [Google Scholar] [CrossRef]
- Sipes, I.G.; el Sisi, A.E.; Sim, W.W.; Mobley, S.A.; Earnest, D.L. Reactive oxygen species in the progression of CCl4-induced liver injury. Adv. Exp. Med. Biol. 1991, 283, 489–497. [Google Scholar]
- Mihas, A.A.; Kanji, V.K.; Mihas, T.A.; Joseph, R.M.; Markov, A.K.; Heuman, D.M. Fructose diphosphate attenuates the acetaminophen-induced liver injury in the rat evidence for involvement of nitric oxide. Res. Commun. Mol. Pathol. Pharmacol. 2003, 113–114, 253–266. [Google Scholar]
- Zhong, Z.; Froh, M.; Wheeler, M.D.; Smutney, O.; Lehmann, T.G.; Thurman, R.G. Viral gene delivery of superoxide dismutase attenuates experimental cholestasis-induced liver fibrosis in the rat. Gene Ther. 2002, 9, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maczewski, M.; Duda, M.; Pawlak, W.; Beresewicz, A. Endothelial protection from reperfusion injury by ischemic preconditioning and diazoxide involves a SOD-like anti-O2- mechanism. J. Physiol. Pharmacol. 2004, 55, 537–550. [Google Scholar] [PubMed]
- Deng, X.; Wu, K.; Wan, J.; Li, L.; Jiang, R.; Jia, M.; Jing, Y.; Zhang, L. Aminotriazole attenuated carbon tetrachloride-induced oxidative liver injury in mice. Food Chem. Toxicol. 2012, 50, 3073–3078. [Google Scholar] [CrossRef] [PubMed]
- Kimmey, M.B.; Lanas, A. Review article: Appropriate use of proton pump inhibitors with traditional nonsteroidal anti-inflammatory drugs and COX-2 selective inhibitors. Aliment. Pharmacol. Ther. 2004, 19, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Ciriminna, R.; Albanese, L.; Di Stefano, V.; Delisi, R.; Avellone, G.; Meneguzzo, F.; Pagliaro, M. Beer produced via hydrodynamic cavitation retains higher amounts of xanthohumol and other hops prenylflavonoids. LWT Food Sci. Technol. 2018, 91, 160–167. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence |
|---|---|
| Cu/Zn-SOD | Forward: 5′-AACCAGTTGTGTTGTCAGGAC-3′ |
| Reverse: 5′-CCACCATGTTTCTTAGAGTGAGG-3′ | |
| Mn-SOD | Forward: 5′-CAGACCTGCCTTACGACTATGG-3′ |
| Reverse: 5′-CTCGGTGGCGTTGAGATTGTT-3′ | |
| CAT | Forward: 5′-GGAGGCGGGAACCCAATAG-3′ |
| Reverse: 5′-GTGTGCCATCTCGTCAGTGAA-3′ | |
| COX-2 | Forward: 5′-GGTGCCTGGTCTGATGATG–3′ |
| Reverse: 5′-TGCTGGTTTGGAATAGTTGCT–3′ | |
| GAPDH | Forward: 5′-AGGTCGGTGTGAACGGATTTG-3′ |
| Reverse: 5′-GGGGTCGTTGATGGCAACA-3′ |
| Group | Body Weight (g) | Liver Weight (g) | Liver Index |
|---|---|---|---|
| Normal | 46.58 ± 1.42 a | 1.94 ± 0.22 b | 4.16 ± 0.3 c |
| Model | 37.11 ± 1.31 b | 2.20 ± 0.22 ab | 5.93 ± 0.4 a |
| BEL | 45.87 ± 1.12 a | 2.44 ± 0.38 a | 5.32 ± 0.4 ab |
| BEH | 46.63 ± 1.08 a | 2.25 ± 0.24 ab | 4.83 ± 0.3 b |
| Silymarin | 47.26 ± 1.22 a | 2.18 ± 0.17 ab | 4.61 ± 0.4 b |
| Group | AST (U/L) | ALT (U/L) | TC (mg/dL) | TG (mg/dL) |
|---|---|---|---|---|
| Normal | 12.37 ± 3.85 d | 5.36 ± 0.87 d | 108.32 ± 5.22 d | 5265.32 ± 87.35 d |
| Model | 149.83 ± 18.32 a | 28.91 ± 3.36 a | 635.28 ± 31.88 a | 16387.20 ± 233.15 a |
| BEL | 109.86 ± 5.25 b | 19.32 ± 3.08 b | 487.36 ± 25.87 b | 11538.69 ± 256.32 b |
| BEH | 58.36 ± 4.82 c | 11.18 ± 2.12 c | 245.23 ± 19.68 c | 8325.05 ± 315.24 c |
| Silymarin | 53.20 ± 6.22 c | 10.65 ± 2.31 c | 239.58 ± 23.89 c | 8158.95 ± 298.23 c |
| Group | IL-6 (pg/mL) | TNF-α (pg/mL) | IFN-γ (pg/mL) |
|---|---|---|---|
| Normal | 23.01 ± 2.13 d | 312.65 ± 11.08 e | 39.12 ± 1.52 d |
| Model | 59.33 ± 3.87 a | 733.48 ± 29.38 a | 81.30 ± 4.72 a |
| BEL | 46.30 ± 2.81 b | 541.69 ± 25.63 b | 65.17 ± 5.25 b |
| BEH | 33.05 ± 2.26 c | 409.71 ± 16.78 c | 51.36 ± 3.97 bc |
| Silymarin | 30.87 ± 3.01 c | 367.97 ± 11.36 d | 47.36 ± 3.32 c |
| Group | SOD (U/mg) | TBARS (nmol/mg) |
|---|---|---|
| Normal | 126.86 ± 8.33 a | 1.51 ± 0.44 d |
| Model | 28.39 ± 6.28 d | 4.83 ± 0.52 a |
| BEL | 69.35 ± 5.59 c | 3.75 ± 0.31 b |
| BEH | 92.79 ± 5.36 b | 2.27 ± 0.25 c |
| Silymarin | 95.02 ± 6.30 b | 2.11 ± 0.29 c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Fang, Y.; Yi, R.; Zhao, X. Preventive Effect of Blueberry Extract on Liver Injury Induced by Carbon Tetrachloride in Mice. Foods 2019, 8, 48. https://doi.org/10.3390/foods8020048
Liu B, Fang Y, Yi R, Zhao X. Preventive Effect of Blueberry Extract on Liver Injury Induced by Carbon Tetrachloride in Mice. Foods. 2019; 8(2):48. https://doi.org/10.3390/foods8020048
Chicago/Turabian StyleLiu, Bihui, Yuan Fang, Ruokun Yi, and Xin Zhao. 2019. "Preventive Effect of Blueberry Extract on Liver Injury Induced by Carbon Tetrachloride in Mice" Foods 8, no. 2: 48. https://doi.org/10.3390/foods8020048
APA StyleLiu, B., Fang, Y., Yi, R., & Zhao, X. (2019). Preventive Effect of Blueberry Extract on Liver Injury Induced by Carbon Tetrachloride in Mice. Foods, 8(2), 48. https://doi.org/10.3390/foods8020048