3.2. Milk Fatty Acid Composition
Feeding system was shown to have a significant effect on the majority of fatty acids analysed, as is shown in
Table 7 and
Table 8.
The fatty acid composition of milk is typically derived from two main sources, through
de novo synthesis and via uptake of preformed fatty acids [
19].
De novo milk fatty acids C4–C14 are synthesised by the cow’s mammary gland. The substrates for
de novo synthesis are acetate and ß-hydroxybutyrate, which are products of rumen fiber digestion [
19]. Therefore, the content of
de novo fatty acids in milk has in recent years received a renewed interest as a result of its use as a potential indicator of rumen functioning in the herd [
20]. Diet was shown to have a significant (
P ≤ 0.001) effect on the
de novo fatty acid content of milks. The PALM diet resulted in milk with the highest level of
de novo, FA significantly higher than that of BEET, SOYA, and CONC (
P ≤ 0.001). However, the increased concentration of C12:0 and C14:0 in PALM diet likely resulted in increased levels of absorption of these fatty acids being reflected in the milks.
Diet was shown to have a significant effect on the total proportion of saturated fatty acids (SFA), which was highest in PALM and lowest in the CONC milk. CONC milk had significantly lower SFA than that of BEET (
P = 0.044) and PALM (
P = 0.006), while SOYA also had significantly lower SFA than that of PALM (
P = 0.019). Dairy fat has been the subject of negative consumer attention in the past particularly attributed to its high levels of SFA. However, while particular SFA such as lauric, myristic, and palmitic acid in isolation can increase low-density lipoprotein (LDL)-cholesterol concentrations, milk is a matrix of other components that have an array of potential benefits including increasing the concentration of high-density lipoprotein (HDL)-cholesterol [
21,
22]. In fact, meta-analyses of the topic have concluded that milk and dairy products consumption have at least a neutral effect on health outcomes [
23], and may protect against prevalent chronic diseases [
24]. However more recent data suggests health considerations of products based on SFA alone can often be misinformed. The level of SFA in milk has significant implications for the functional characteristics and processability of high-fat dairy products. Recent studies have demonstrated that increased SFA (particularly C16:0) results in significant alterations to hardness and textural properties of dairy products [
4,
6]. The C16:0 content was highest in BEET-derived milk, which was significantly greater than that of PALM milk (
P = 0.013). Palmitic acid (C16:0) is the most abundant SFA in bovine milk; increased proportions of C16:0 in BEAT-derived milks corresponds with higher contents of C16:0 in the BEET diet (
Table 5).
Among total unsaturated fatty acids (UFA), CONC diet resulted in significantly higher UFA than that of PALM (
P = 0.001). PALM diet resulted in milk with significantly lower poly unsaturated fatty acids (PUFA) than that of BEET, CONC, and SOYA (
P ≤ 0.001). Among dairy PUFA, the isomer conjugated linoleic acid (CLA) c9t11 has been studied for potential health benefits and biological functions, which include impacting immune function and protective effects against cancer, obesity, and atherosclerosis [
25]. The fatty acid CLAc9t11 is produced via the biohydrogenation of linoleic acid in the rumen by rumen microorganisms [
26]. Therefore, CLAc9t11 in milk is highly dependent on the cow feeding system used [
27] and the level of fatty acid substrate available has a direct effect on the its subsequent levels in milk. PALM feeding of cows resulted in milk with significantly reduced CLAc9t11 than that of BEET (
P = 0.002), CONC (
P ≤ 0.001), and SOYA (
P = 0.001) milks. This result can be attributed to the substantially lower levels of linoleic acid in the feed supplement of PALM compared to the other diets (
Table 5).
Diet did not have a significant effect of the total level of mono-unsaturated fatty acids (MUFA) of milk (
P = 0.08). Among the total content of short-chain fatty acids (SCFA), PALM milk had significantly higher SCFA than that of BEET, CONC, and SOYA (
P ≤ 0.001). Among the medium-chain fatty acids (MCFA), BEET milk had significantly higher MCFA than that of PALM milk (
P = 0.014). Among long-chain fatty acids (LCFA), the CONC diet produced milk with highest content of LCFA, significantly higher than that of BEET (
P = 0.021). However, while increased levels of LCFA can have a positive effect on milk fatty acid profile, increased levels of unsaturated LCFA can have a negative effect on the oxidative stability of milk fat [
28].
Given changes in nutritional perceptions of milk and milk fatty acids as a whole, it could be suggested that judging dairy products by their total SFA content alone can be misinformed and erroneous, while other indices such levels of Omega (Ω) 3 and Ω 6 fatty acids should also be considered. As discussed by Patterson et al. [
29], both Ω 3 and Ω 6 are essential fatty acids and precursors to anti-inflammatory and pro-inflammatory eicosanoids respectively. However, dietary changes in line with the western diet has resulted in an increase of Ω 6 fatty acids in the diet coinciding with increases in prevalence of chronic inflammatory diseases. CONC feeding was demonstrated to produce milks with highest level of Ω 3 fatty acids. PALM feeding, however, resulted in milk with significantly lower Ω 3 content than that of CONC (
P ≤ 0.001), BEET (
P = 0.002), and SOYA (
P = 0.001). PALM feeding also produced milk with significantly lower Ω 6 than that of BEET (
P = 0.017) and SOYA (
P = 0.005). The thrombogenic (TI) and atherogenic (AI) indices are both dietary risk indices for cardiovascular disease. CONC milk had the lowest AI of each of the diets and was significantly lower than that of BEET (
P = 0.031) and PALM (
P = 0.004). The SOYA diet also resulted in a significantly lower AI than that of PALM (
P = 0.022).
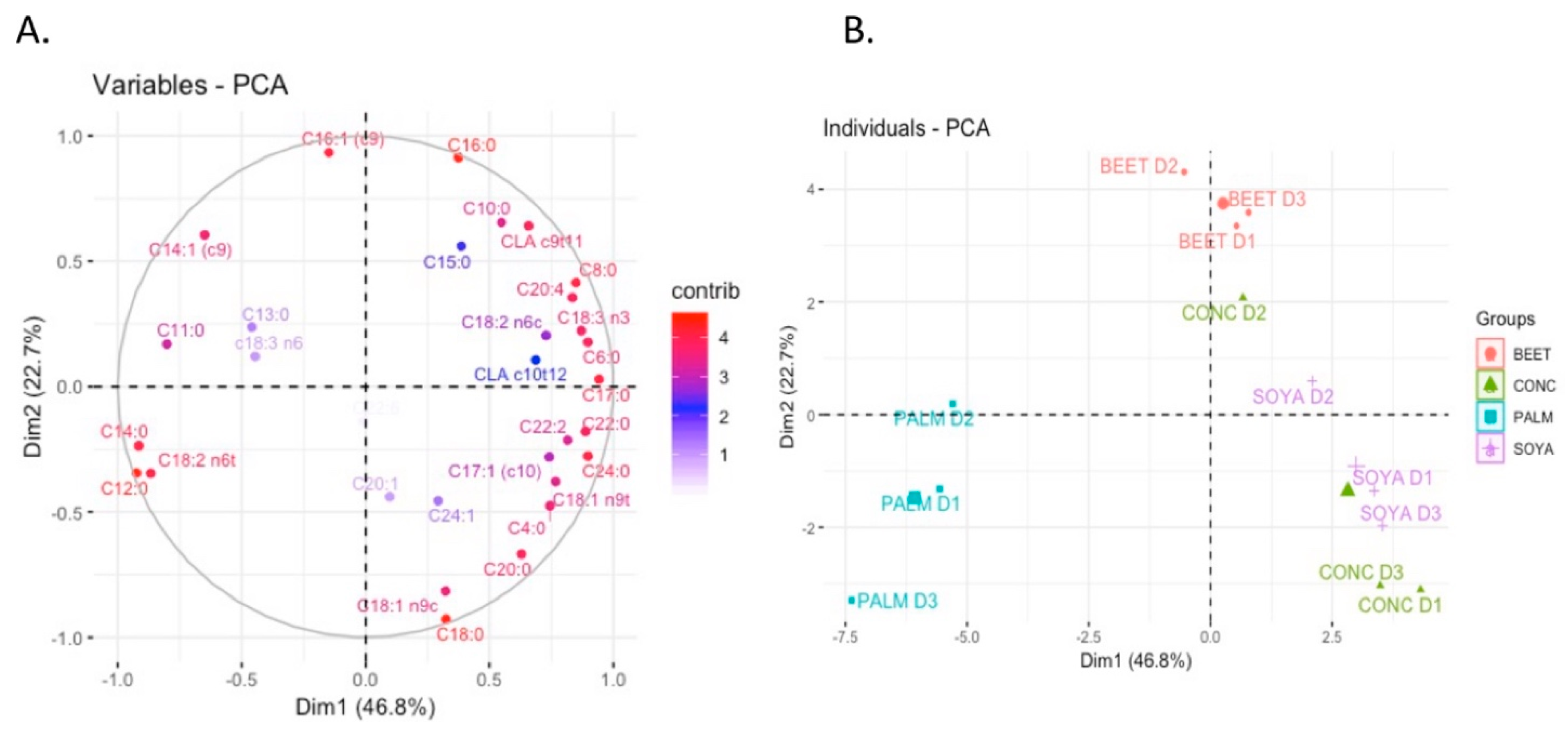
Multivariate analysis of the fatty acids of milks has previously been shown capable of distinguishing milks from pasture versus conventional feeding systems [
3]. Similar results have been shown in this study where there is clear separation between the SOYA-, BEET-, and PALM-derived milks. However, each of the diets appeared to share a similar composition with that of the CONC. This is perhaps to be expected given the similarities between the base ingredients of CONC and other diets composition (
Figure 1). The top 10 fatty acids for the observed separation in
Figure 1 are shown in
Supplementary Figure S1, which include C16:0, C12:0, C18:0, C14:0, C16:1, C8:0, C17:0, C24:0, C18:2n6t, and CLA c9t11. Lauric acid was highest in PALM-derived milks, significantly higher than that of CONC, BEET, and SOYA (
P ≤ 0.001). While C12:0 can be produced by
de novo synthesis, the levels of the fatty acid in the diet can have a direct effect on its concentrations in the final milks as was demonstrated by Hristov et al. [
30], who reported supplementation of C12:0 doubled its concentrations in raw milk. Increased levels of C12:0 in PALM milk corresponds with increased levels in the PALM feed compared to that of the other diets (
Table 5). Stearic acid (C18:0) could be referred to have a neutral effect on human health, as it has not been shown to increase total cholesterol or LDL-cholesterol concentrations [
31]. BEET-derived milks had significantly reduced concentrations than that of CONC (
P = 0.019) and SOYA (
P = 0.047). Myristic acid, along with palmitic acid, have been demonstrated to have cholesterol increasing properties [
32]. Diet had a significant effect on the milk content of myristic acid (C14:0) where it was significantly higher in PALM than that of BEET, SOYA, and CONC (P ≤ 0.001). CONC feeding resulted in the lowest level of C14:0 in milk, which was also significantly lower than that of BEET (
P = 0.044). Such results follow similar trend to the level of C14:0 being provided by the diet. These results are in agreement with those of Hristov et al. [
30], who reported an increase in C14:0 in milk from cows supplemented with myristic acid. Caprylic acid (C8:0) was lowest in PALM milk, significantly lower than CONC (
P = 0.002), SOYA (
P ≤ 0.001), and BEET (
P ≤ 0.001). Levels of odd and branched chain have been receiving increased attention. In particular, C15:0 and C17:0 have been used as biomarkers of intake of ruminant fat by humans [
32]. Odd and branched-chain fatty acids such as C15:0 and C17:0 are derived from
de novo synthesis in the mammary gland from propionate in ruminants, and diet of cows has been demonstrated to have a significant effect on their concentrations in milk [
32]. C17:0 was lowest in the milks in PALM-derived milks, significantly lower than CONC, BEET, or SOYA (
P ≤ 0.001).
O’Callaghan et al. [
3] and Capuano et al. [
33] found that milk fatty acid profiling could be used as a potential tool for verification of cow feeding system, and as such these results progresses this concept in that it is also capable of distinguishing small changes in the pasture-based diet such as the inclusion of supplements.
3.3. Milk Volatile Analysis
Volatile analysis of milks using untargeted GC-MS SPME technique identified 34 compounds present in each of the samples (
Table 9). Measurement of the volatiles in the headspace of products provides a representative view of the volatiles relative to the olfactory response [
34]. There are a variety of factors that can impact the volatile profile of milk. A comprehensive review of the topic has highlighted a direct transfer of many volatiles from grazing or from conserved forage, some of which are related to plant (forage) metabolism or fermentation during conservation processes but many volatiles are also transferred indirectly post rumen metabolism [
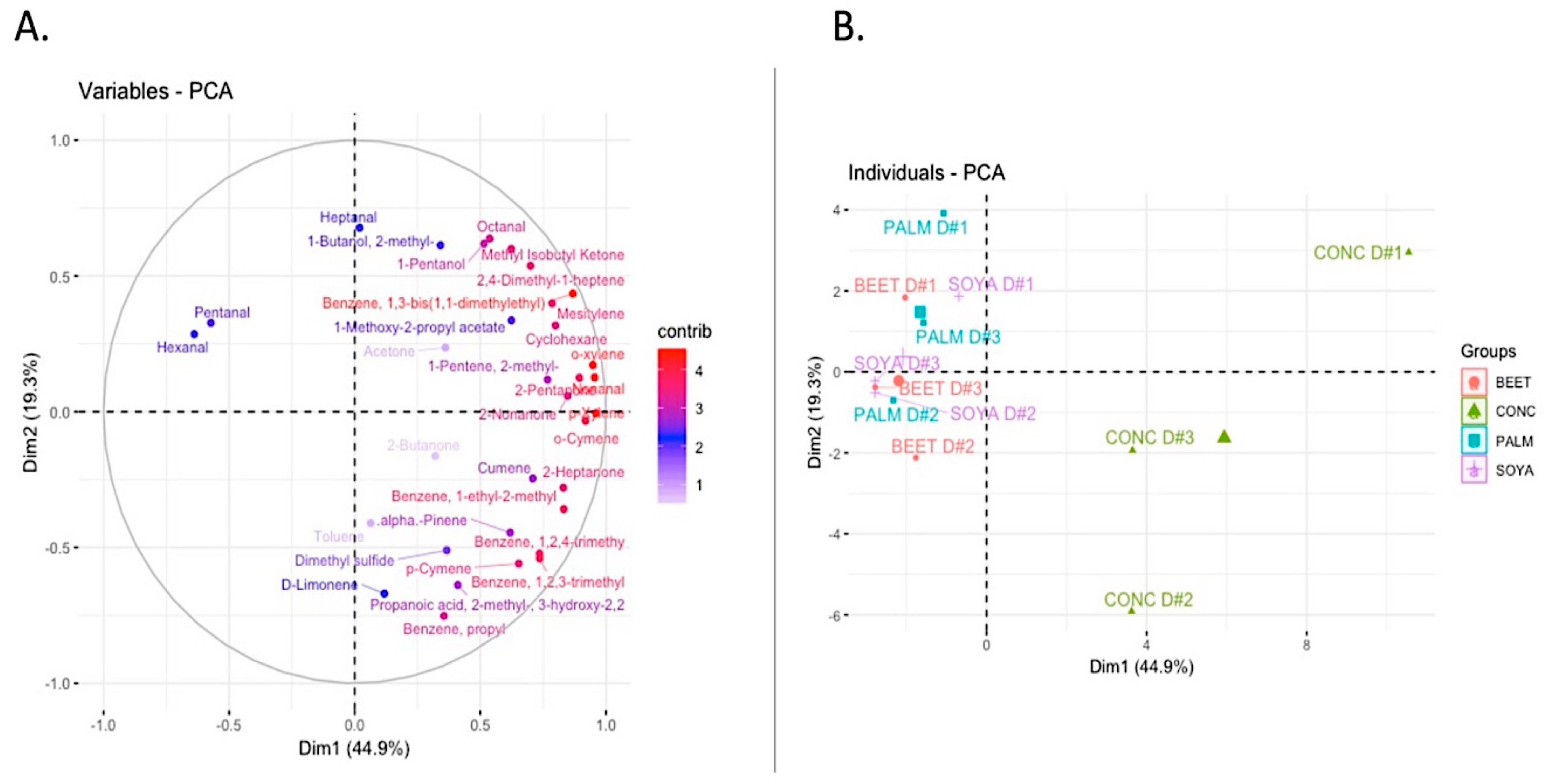
35]. In total, 33 compounds were detected in milks from each of the treatments from a variety of chemical groups including benzenes (10), ketones (6), sulfur compounds (1), alkenes (2), aldehydes (5), terpenes (4), and alcohols (2). Principal component analysis of the data revealed that the volatile profile of the CONC diet was distinctly different to the other supplementary feeding systems (
Figure 2). A number of volatiles were identified to be responsible for this separation (
Figure 2A). Dietary treatment was demonstrated to have a significant effect on the volatile profile of milks. Pentanal and hexanal are both aldehyde compounds typically derived from the degradation and oxidation of fatty acids. Pentanal was present at significantly (
P < 0.05) higher concentrations in PALM than that of BEET and CONC, while levels of pentanal in SOYA were also significantly higher than that of CONC. Hexenal was significantly higher in BEET-, PALM-, and SOYA-derived milks than that of CONC. Hexanal mostly results from the degradation of oleic acid and linoleic acid, while pentenal can be derived from the oxidation of arachidonic and linoleic acid. Indeed, the levels of pentanal in milks were significantly and positively correlated with concentration of milk linoleic acid (
P = 0.019,
r = 0.661). The volatile p-xylene was present at significantly higher levels in CONC than that of BEET, PALM, and SOYA. The presence of p-xylene may be the result of carotenoid degradation, namely ß-carotene degradation [
36], or possibly directly transferred from the feed [
37].
3.4. Native Casein Micelle Size and Milk Gelation Properties
Feeding system was demonstrated to have a significant effect (
P = 0.013) on the size of casein micelles and the gelation properties of the milks (
Table 10). All z-average size distributions showed normal, mono-modal distributions. The CONC diet milks had the largest micelle size (167 nm), which was significantly greater than that of SOYA milk (153 nm) (
P = 0.011), while the micelle size of PALM milks (163 nm) was also greater than that of SOYA (
P = 0.058). Devold et al. [
38] reported a similar trend of effect of feeding regimen on casein micelle size, whereby cows fed an ecologically rolled barley supplement produced significantly larger casein micelles than cows fed commercial supplementary concentrates. While genetic factors are typically reported to determine the size of casein micelles, micelle size is also affected by heat treatment, which causes pH-dependent interactions between β-lactoglobulin and κ-casein (on or off the casein micelle). Such factors impact on the volume fraction and packing arrangements of interacting casein micelles. The presence of minerals, especially calcium, also affect micellar interactions through neutralization of charge repulsion-induced stability. Neither of these influences were determined in the present study but should be considered in the future.
Alterations to casein micelle size and other characteristics of milk can be an important consideration as it has been previously demonstrated to play a significant role in the gelation properties of milks [
39]. Glantz et al. [
39] showed that milk gelation was affected by pH, protein profile, casein micelle size, and lactose. Gelation of milk forms the basis for the production of stabilized, high-value dairy products such as yoghurt, cheese, and their variants. There was a significant effect between feeding systems on the gelation properties of milks (
P ≤ 0.01). The storage modulus of SOYA-derived milks was significantly higher than each of the other diets, storage modulus of SOYA > PALM > BEET > CONC, respectively. The CONC milk had the lowest storage modulus, significantly lower than that of PALM (
P = 0.016) and SOYA (
P ≤ 0.01). A similar trend was observed for loss modulus measurement whereby SOYA had significantly higher scores than BEET (
P = 0.003) and CONC (
P = 0.001), loss modulus of SOYA > PALM > BEET > CONC.
Gelation properties of milks can be influenced by a variety of factors including protein profile, milk composition, and stage of lactation. It seems that differences in the gelation properties observed in this study correlated with diet-induced changes to casein micelle size. Indeed, there was a significant negative correlation between casein micelle size and storage and loss modulus values (
P = 0.009,
r = −0.712 and
P = 0.014,
r = −0.686, respectively). It has been demonstrated, however, that combined changes in the casein micelle size and MFG size can influence curd firmness [
40]. While the links between cow feeding system and casein micelle size requires further investigation, given these findings on impact of cow diet supplementation on milk gelation properties, dairy manufacturers should be aware of the types of dietary supplements used on farms and the potential implications for downstream product properties.

 ,
, 



{kind=link}
{kind=link}