Extraction and Characterization of Self-Assembled Collagen Isolated from Grass Carp and Crucian Carp

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of Collagens
2.3. UV Absorption Spectra
2.4. FTIR
2.5. SDS-PAGE Analysis
2.6. DSC
2.7. Self-Assembly of Collagen in Vitro
2.8. Measurement of Collagen Fibril Formation
2.9. SEM
3. Results and Discussion
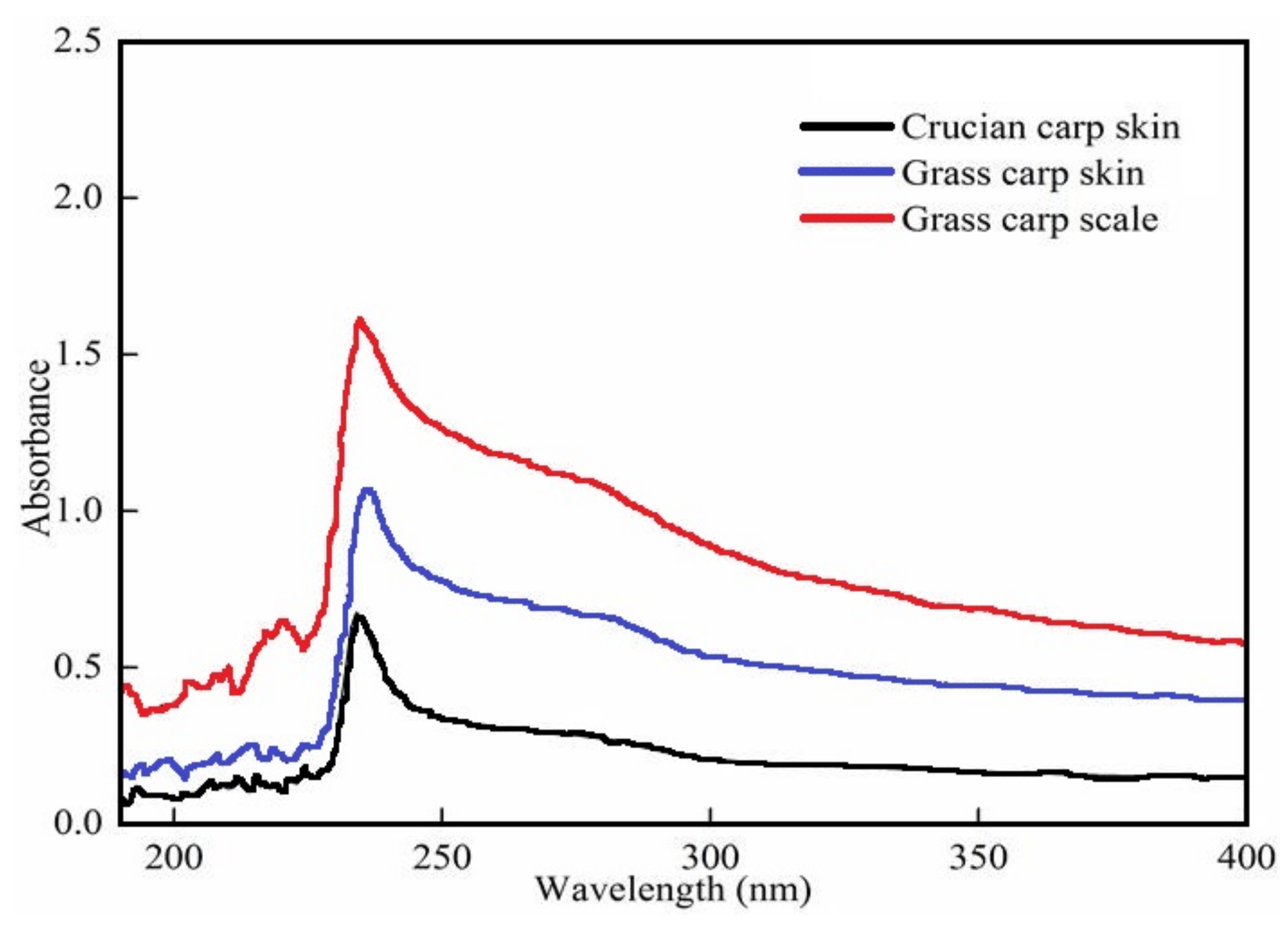
3.1. UV Absorption Spectra of the Extracted Collagen
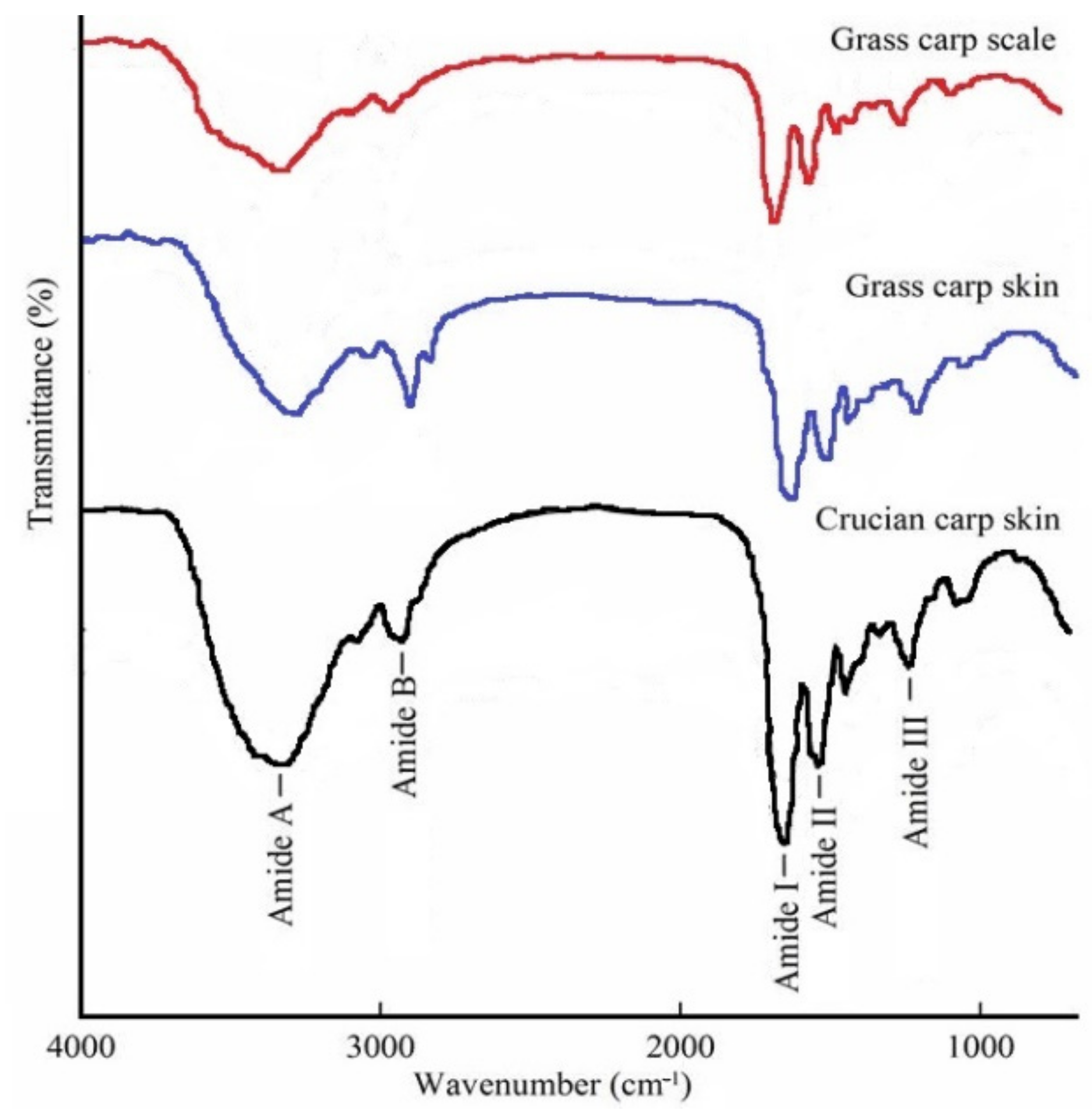
3.2. FTIR Spectroscopy
3.3. SDS-PAGE
3.4. DSC Thermograms
3.5. Self-Assembly of Collagen
3.6. The Morphology of Collagen Films
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Guo, H.; Hong, Z.; Yi, R. Core-shell collagen peptide chelated calcium/calcium alginate nanoparticles from fish scales for calcium supplementation. J. Food Sci. 2015, 80, N1595–N1601. [Google Scholar] [CrossRef]
- Li, C.; Liao, X.; Bi, S. Using collagen fiber as a template to synthesize Tio2 and Fex/Tio2 nanofibers and their catalytic behaviors on the visible light-assisted degradation of orange II. Ind. Eng. Chem. Res. 2010, 49, 3194–3199. [Google Scholar]
- Kadler, K.E.; Baldock, C.; Bella, J.; Boot-Handford, R.P. Collagens at a glance. J. Cell Sci. 2007, 120, 1955. [Google Scholar] [CrossRef] [PubMed]
- Purohit, T.; He, T.; Qin, Z.; Li, T.; Fisher, G.J.; Yan, Y.; Voorhees, J.J.; Quan, T. Smad3-dependent regulation of type i collagen in human dermal fibroblasts: Impact on human skin connective tissue aging. J. Dermatol. Sci. 2016, 83, 80–83. [Google Scholar] [CrossRef]
- Liu, D.; Liang, L.; Regenstein, J.M.; Peng, Z. Extraction and characterisation of pepsin-solubilised collagen from fins, scales, skins, bones and swim bladders of bighead carp (Hypophthalmichthys nobilis). Food Chem. 2012, 133, 1441–1448. [Google Scholar] [CrossRef]
- Noitup, P.; Morrissey, M.T.; Garnjanagoonchorn, W. In vitro self-assembly of silver-line grunt type 1 collagen: Effects of collagen concentrations, ph and temperatures on collagen self-assembly. J. Food Biochem. 2006, 30, 547–555. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Benjakul, S.; Visessanguan, W.; Nagai, T.; Tanaka, M. Characterisation of acid-soluble collagen from skin and bone of bigeye snapper (Priacanthus tayenus). Food Chem. 2005, 89, 363–372. [Google Scholar] [CrossRef]
- Yan, M.; Li, B.; Zhao, X.; Qin, S. Effect of concentration, ph and ionic strength on the kinetic self-assembly of acid-soluble collagen from walleye pollock (Theragra chalcogramma) skin. Food Hydrocoll. 2012, 29, 199–204. [Google Scholar] [CrossRef]
- Zhang, X.; Ookawa, M.; Tan, Y.; Ura, K.; Adachi, S.; Takagi, Y. Biochemical characterisation and assessment of fibril-forming ability of collagens extracted from bester sturgeon huso huso acipenser ruthenus. Food Chem. 2014, 160, 305–312. [Google Scholar] [CrossRef]
- Fessler, J. Self-assembly of collagen. J. Supramol. Struct. 1974, 2, 99–102. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, D.; Wang, Y.; Qin, W. A comparative study of the properties and self-aggregation behavior of collagens from the scales and skin of grass carp (Ctenopharyngodon idella). Int. J. Biol. Macromol. 2018, 106, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Jongjareonrak, A.; Benjakul, S.; Visessanguan, W.; Nagai, T.; Tanaka, M. Isolation and characterisation of acid and pepsin-solubilised collagens from the skin of Brownstripe red snapper (Lutjanus vitta). Food Chem. 2005, 93, 475–484. [Google Scholar] [CrossRef]
- Birk, D.E.; Bruckner, P. Collagen Suprastructures; Springer: Berlin/Heidelberg, Germany, 2005; pp. 185–205. [Google Scholar]
- Friess, W. Collagen—Biomaterial for drug delivery. Eur. J. Pharm. Biopharm. 1998, 45, 113. [Google Scholar] [CrossRef]
- Kumar, N.S.S.; Nazeer, R.A. Erratum to: Wound healing properties of collagen from the bone of two marine fishes. Int. J. Pept. Res. Ther. 2012, 18, 193. [Google Scholar] [CrossRef][Green Version]
- Chen, J.; Li, L.; Yi, R.; Xu, N.; Gao, R.; Hong, B. Extraction and characterization of acid-soluble collagen from scales and skin of tilapia (Oreochromis niloticus). LWT Food Sci. Technol. 2016, 66, 453–459. [Google Scholar] [CrossRef]
- Pal, G.K.; Nidheesh, T.; Suresh, P.V. Comparative study on characteristics and in vitro fibril formation ability of acid and pepsin soluble collagen from the skin of catla (Catla catla) and rohu (Labeo rohita). Food Res. Int. 2015, 76, 804–812. [Google Scholar] [CrossRef]
- Chen, S.; Chen, H.; Xie, Q.; Hong, B.; Chen, J.; Hua, F.; Bai, K.; He, J.; Yi, R.; Wu, H. Rapid isolation of high purity pepsin-soluble type i collagen from scales of red drum fish (Sciaenops ocellatus). Food Hydrocoll. 2016, 52, 468–477. [Google Scholar] [CrossRef]
- Wu, Q.Q.; Li, T.; Wang, B.; Ding, G.F. Preparation and characterization of acid and pepsin-soluble collagens from scales of croceine and redlip croakers. Food Sci. Biothechnol. 2015, 24, 2003–2010. [Google Scholar] [CrossRef]
- Doyle, B.B.; Bendit, E.G.; Blout, E.R. Infrared spectroscopy of collagen and collagen-like polypeptides. Biopolymers 1975, 14, 937–957. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Nalinanon, S.; Benjakul, S.; Kishimura, H. Characteristics of pepsin-solubilised collagen from the skin of splendid squid (Loligo formosana). J. Chem. 2015, 2015, 1–8. [Google Scholar] [CrossRef]
- Abe, Y.; Krimm, S. Normal vibrations of crystalline polyglycine I. Biopolymers 1972, 11, 1817–1839. [Google Scholar] [CrossRef] [PubMed]
- Guzzi Plepis, A.M.D.; Goissis, G.; Das-Gupta, D.K. Dielectric and pyroelectric characterization of anionic and native collagen. Polym. Eng. Sci. 1996, 36, 2932–2938. [Google Scholar] [CrossRef]
- Krimm, S.; Bandekar, J. Vibrational spectroscopy and conformation of peptides, polypeptides, and proteins. In Advances in Protein Chemistry; Academic Press: Cambridge, MA, USA, 1986; Volume 38, p. 181. [Google Scholar]
- Sionkowska, A.; Kozłowska, J.; Skorupska, M.; Michalska, M. Isolation and characterization of collagen from the skin of brama australis. Int. J. Biol. Macromol. 2015, 80, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Benjakul, S.; Nalinanon, S. Compositional and physicochemical characteristics of acid solubilized collagen extracted from the skin of unicorn leatherjacket (Aluterus monoceros). Food Hydrocoll. 2010, 24, 588–594. [Google Scholar] [CrossRef]
- Matmaroh, K.; Benjakul, S.; Prodpran, T.; Encarnacion, A.; Kishimura, H. Characteristics of acid soluble collagen and pepsin soluble collagen from scale of spotted golden goatfish (Parupeneus heptacanthus). Food Chem. 2011, 129, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Kittiphattanabawon, P.; Benjakul, S.; Visessanguan, W.; Shahidi, F. Isolation and properties of acid- and pepsin-soluble collagen from the skin of blacktip shark (Carcharhinus limbatus). Eur. Food Res. Technol. 2010, 230, 475–483. [Google Scholar] [CrossRef]
- Mori, H.; Tone, Y.; Shimizu, K.; Zikihara, K.; Tokutomi, S.; Ida, T.; Ihara, H.; Hara, M. Studies on fish scale collagen of pacific saury (Cololabis saira). Mat. Sci. Eng. C 2013, 33, 174. [Google Scholar] [CrossRef]
- Muyonga, J.; Cole, C.; Duodu, K. Characterisation of acid soluble collagen from skins of young and adult Nile perch (Lates niloticus). Food Chem. 2004, 85, 81–89. [Google Scholar] [CrossRef]
- Usha, R.; Ramasami, T. The effects of urea and n -propanol on collagen denaturation: Using dsc, circular dicroism and viscosity. Thermochim. Acta 2004, 409, 201–206. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, X.; Li, T.; Yang, H.; Zhang, H.; Regenstein, J.M.; Zhou, P. Extraction and characterization of acid- and pepsin-soluble collagens from the scales, skins and swim-bladders of grass carp (Ctenopharyngodon idella). Food Biosci. 2015, 9, 68–74. [Google Scholar] [CrossRef]
- Singh, P.; Benjakul, S.; Maqsood, S.; Kishimura, H. Isolation and characterisation of collagen extracted from the skin of striped catfish (Pangasianodon hypophthalmus). Food Chem. 2011, 124, 97–105. [Google Scholar] [CrossRef]
- Ahmad, M.; Benjakul, S. Extraction and characterisation of pepsin-solubilised collagen from the skin of unicorn leatherjacket (Aluterus monocerous). Food Chem. 2010, 120, 817–824. [Google Scholar] [CrossRef]
- Pati, F.; Adhikari, B.; Dhara, S. Isolation and characterization of fish scale collagen of higher thermal stability. Biores. Technol. 2010, 101, 3737–3742. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Tu, X.; Wang, W.; Nan, J.; Wei, B.; Xu, C.; Lang, H.; Yuling, X.; Sheng, L.; Wang, H. Insight into the role of grafting density in the self-assembly of acrylic acid-grafted-collagen. Int. J. Biol. Macromol. 2019, 128, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, C.; Liu, C. Studies on properities of collagen from fish scale. Mar. Fish. Res. 2000, 21, 57–61. [Google Scholar]
- Tziveleka, L.; Ioannou, E.; Tsiourvas, D.; Berillis, P.; Foufa, E.; Roussis, V. Collagen from the marine sponges Axinella cannabina and Suberites carnosus: Isolation and morphological, biochemical, and biophysical characterization. Mar. Drugs 2017, 15, 152. [Google Scholar] [CrossRef] [PubMed]
- Berillis, P. Marine collagen: Extraction and applications. In Research Trends in Biochemistry, Molecular Biology and Microbiology; SM Group: Dover, DE, USA, 2015; pp. 1–13. [Google Scholar]
- Nagai, T.; Yamashita, E.; Taniguchi, K.; Kanamori, N.; Suzuki, N. Isolation and characterisation of collagen from the outer skin waste material of cuttlefish (Sepia lycidas). Food Chem. 2001, 72, 425–429. [Google Scholar] [CrossRef]
- Aukkanit, N.; Garnjanagoonchorn, W. Temperature effects on type I pepsin-solubilised collagen extraction from silver-line grunt skin and its in vitro fibril self-assembly. J. Sci. Food Agric. 2010, 90, 2627. [Google Scholar] [CrossRef]
- Yan, M.; Qin, S.; Li, J. Study on the self-assembly property of type i collagen prepared from tilapia (Oreochromis niloticus) skin by different extraction methods. Int. J. Food Sci. Technol. 2015, 50, 2088–2096. [Google Scholar] [CrossRef]
- Li, Y.; Asadi, A.; Monroe, M.R.; Douglas, E.P. Ph effects on collagen fibrillogenesis in vitro: Electrostatic interactions and phosphate binding. Mat. Sci. Eng. 2009, 29, 1643–1649. [Google Scholar] [CrossRef]
- Lin, Y.K.; Liu, D.C. Effects of pepsin digestion at different temperatures and times on properties of telopeptide-poor collagen from bird feet. Food Chem. 2006, 94, 621–625. [Google Scholar] [CrossRef]
- Rasheeda, K.; Bharathy, H.; Fathima, N. Vanillic acid and syringic acid: Exceptionally robust aromatic moieties for inhibiting in vitro self-assembly of type I collagen. Int. J. Bol. Macromol. 2018, 113, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhou, P.; Li, T.; Regenstein, J.M. Comparison of acid-soluble collagens from the skins and scales of four carp species. Food Hydrocoll. 2014, 41, 290–297. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.; Lan, W.; Wang, Y.; Ahmed, S.; Liu, Y. Extraction and Characterization of Self-Assembled Collagen Isolated from Grass Carp and Crucian Carp. Foods 2019, 8, 396. https://doi.org/10.3390/foods8090396
He L, Lan W, Wang Y, Ahmed S, Liu Y. Extraction and Characterization of Self-Assembled Collagen Isolated from Grass Carp and Crucian Carp. Foods. 2019; 8(9):396. https://doi.org/10.3390/foods8090396
Chicago/Turabian StyleHe, Li, Wenting Lan, Yue Wang, Saeed Ahmed, and Yaowen Liu. 2019. "Extraction and Characterization of Self-Assembled Collagen Isolated from Grass Carp and Crucian Carp" Foods 8, no. 9: 396. https://doi.org/10.3390/foods8090396
APA StyleHe, L., Lan, W., Wang, Y., Ahmed, S., & Liu, Y. (2019). Extraction and Characterization of Self-Assembled Collagen Isolated from Grass Carp and Crucian Carp. Foods, 8(9), 396. https://doi.org/10.3390/foods8090396